
放牧对高寒草地4种草光响应和荧光特性的影响
2012-08-20 08:01陈建纲张德罡徐长林柳小妮
草业科学 2012年4期
孙 英,陈建纲,张德罡,徐长林,柳小妮
(甘肃农业大学草业学院 草业生态系统教育部重点实验室 中美草地畜牧业可持续发展研究中心,甘肃 兰州730070)
祁连山地处青藏、蒙新、黄土三大高原交汇地带,作为青藏高原和北部内陆荒漠地区重要的分水岭,形成了独具特色的生物多样性和成土条件[1],在维系西北高原气候、保持水土方面具有重要的战略意义。祁连山东段天祝县高寒草地分布在高海拔、低热量的生态环境中,植被外貌低矮,组成植物多,系低温生态类型;利用方式和改良途径有较高的一致性,且具有较高的生产力和经济价值[2];同时,是当地最重要的草地资源,畜牧业发展的支柱。高寒草地的主要利用方式是放牧,而草地生态系统的首要功能是通过绿色植物的光合作用固定太阳能,进行植物生产,供家畜采食。同时,光合作用可以直观地反映草地植物的生长状况和在放牧后补偿生长的情况和能力。因此,在高寒草地开展光合作用的研究,对于适时、合理地利用、保护、恢复草地具有非常重要的现实意义和理论意义。
在高寒地区,7-9月是牧草生长最旺盛的季节,而天祝县年降水也主要集中于这3个月,自然光照不足会导致光合作用的日变化无法真实反映植物的光合水平和潜力。而用人工光源模拟光梯度则可以从一定程度上解决这一问题。植物光合-光响应曲线是在一定的模拟光强梯度下绘制的光合速率随光强变化的曲线,反映了净光合速率随光强变化的特性[3],通过拟合光响应曲线可以了解植物光反应过程的光合效率[4]。叶绿素荧光也可以作为光合作用研究的有效探针。目前,关于高寒草地光合生理特征,国内外学者做了一定的研究[5-11],大都集中于某些草种的光合作用日变化。在放牧啃食和践踏显著影响植物对光和水分的利用方面,国内外学者也做过一些研究[12-14]。常乐等[15]研究过4种园林地被植物在受到践踏胁迫后叶绿素荧光特性的变化,证明叶绿素荧光参数能很好地反应植物受到的胁迫情况。本研究总结前人的经验,从光响应曲线和荧光参数的差异等方面,探讨了草地4种优势植物在放牧草地(Grazing grassland,G)和禁 牧草地(Non-grazing grassland,NG)中光合特性的变化情况,以期探讨放牧对高寒草地植物光合作用的影响,为适时、合理地利用和保护高寒草地提供科学的理论依据。
1 材料与方法
1.1 研究区域自然概况 研究区域位于甘肃省天祝县金强河地区,地处祁连山东段,海拔2 920~3 800m,气候寒冷潮湿,空气稀薄,太阳辐射强,水热同期。年均温-0.1℃,1月均温-18.3℃,7月均温12.4℃;全年无绝对无霜期,仅分冷、热两季,冷季长达7个月。植物生长季为120~140d。年均降水量416mm,多为地形雨,集中于7-9月;年均蒸发量1 592mm[16]。
1.2 样地概况 样地设置在南泥沟河谷地,甘肃农业大学天祝高山草原试验站,海拔2 950m。站周围建有围栏,围栏内为禁牧草地,作为对照,面积约0.15hm2,于2004年夏禁牧至今;20cm土壤容积含水量为5.1%。围栏外侧为放牧草地,面积约0.67hm2,于每年5-11月进行放牧,其中有成年羊19只,羊羔14只,放牧强度为39羊单位·hm-2;20 cm土壤容积含水量为6.4%。该草地属于披碱草-嵩草草地型,牧草种类丰富,优势种有矮嵩草(Kobresia humilis)、垂穗披碱草(Elymus nutans)、赖草(Leymus secalinus)、珠芽蓼(Polygonum viviparum)、扁蓿豆(Melissitus rutenica)、黄花棘豆(Oxytropis ochrocephala)、二裂委陵菜(Potentilla bifurca)等,植被盖度95%。
试验材料为该草地型中频度最高的矮嵩草、垂穗披碱草、扁蓿豆、黄花棘豆4种植物。
1.3 方法 试验选择晴朗天气(7月1-10日),采用GFS-3000便携式光合仪,测定时段为9:30-11:30,每次随机选取每种优势植物的1个充分展开的叶片。测定过程中,将叶温设置为25℃,相对湿度设置为60%,CO2浓度设为390μmol·mol-1,用GFS-3000自带光源模拟光强梯度为2 000、1 700、1 400、1 100、800、500、200、100、50、25、10、0 μmol·m-2·s-1,改变光照强度后,最少稳定时间设置为60s,当测量结果变异率小于0.05时记录数据,3次重复。
植物的各种光响应参数需通过一些经验模型或机理模型的拟合来得到。目前,直角双曲线模型和非直角双曲线模型应用最广泛[17]。徐伟洲等[18]通过拟合白羊草(Bothriochloa ischaemum)的光响应曲线,比较了这两种模型,得出非直角双曲线模型的拟合结果更为准确。因此,本研究采用双曲线模型来进行拟合。
在0~200μmol·m-2·s-1光合有效辐射强度之间,净光合速率-光合有效辐射(Pn-PAR)回归直线方程与X轴的交点(即Pn为零时),得出植物的光补偿点(LCP);直线方程与Y轴的交点(即PAR为零时),得出植物的暗呼吸速率(Rd);而表观量子效率即此直线方程的斜率(a)。
荧光参数的测定从早上9:00开始。利用Imaging-PAM便携式叶绿素荧光仪测定叶绿素荧光参数。用黑布遮光,叶片暗适应20min以上后进行测定,每个处理测定2~4个叶片。经过充分暗适应后,打开弱测量光(不大于0.1μmol·m-2·s-1)测定初始荧光(F0),此时再打开一次饱和脉冲光(3 000μmol·m-2·s-1),测定叶绿素的荧光Fm。然后在自然光下,光下稳态荧光(Ft)稳定后,给一次饱和脉冲光测定Fm′。
由以上几个参数便可以计算出:PSⅡ的最大量子产量(Fv/Fm,反映植物的潜在最大光合能力)、PSⅡ的实际量子产量(Yield,反映植物的实际光合效率)、电子传递的相对速率(ETR)、光化学淬灭(qP,由光合作用引起的荧光淬灭)、非光化学淬灭(NPQ,由热耗散引起的荧光淬灭)等荧光参数值。1.4数据处理 数据结果采用Excel 2003绘图,用SPSS 17.0对数据进行统计与分析。
2 结果与分析
2.1 放牧对光响应曲线的影响 在一定CO2浓度下,随着光合有效辐射(PAR)的增强,4种草本植物净光合速率(Pn)逐渐增大,PAR在0~900 μmol·m-2·s-1之间时Pn增加速度最快,之后这种增大的趋势逐渐减弱(图1)。当PAR超过一定范围后,Pn开始下降。然而,禁牧草地和放牧草地中植物的光合光响应曲线有所不同。
在禁牧草地(NG)中,随着PAR的增强,4种植物Pn整体水平从大到小依次为:扁蓿豆、矮嵩草、黄花棘豆、垂穗披碱草。矮嵩草的光饱和点(LSP)最高,为1 700μmol·m-2·s-1;垂穗披碱草的LSP最低,为1 300μmol·m-2·s-1(表1)。黄花棘豆的光补偿总(LCP)最高(44.36μmol·m-2·s-1),扁蓿豆次之,矮嵩草最低(19.44μmol·m-2·s-1)。最大净光合速率 (Pmax)的 排 列 顺 序 为 :扁 蓿 豆 (1 0.6 7 μmol·m-2·s-1)、矮嵩草(10.23μmol·m-2·s-1)、黄花棘豆 (9.67μmol·m-2·s-1)、垂穗披碱草(8.80μmol·m-2·s-1)。
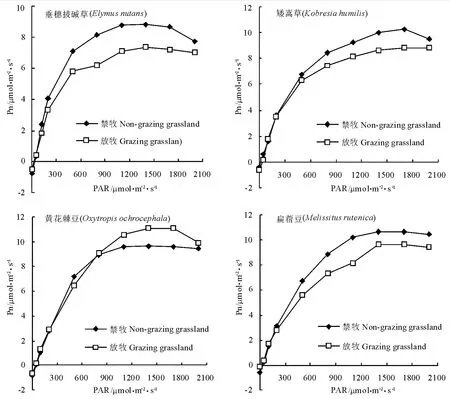
图1 放牧和禁牧草地中4种优势植物光响应曲线的对比Fig.1 Photosynthesis-light response curves of four dominant plants in the grazing and Non-grazing conditions
在放牧草地(G)中,随着PAR的增强,黄花棘豆的Pn增加速度最快;垂穗披碱草的Pmax最低。矮嵩草的LSP仍然最高,扁蓿豆和垂穗披碱草的LSP最低,这与禁牧地的规律相似。LCP的排列顺序略不同于禁牧地,但最高的仍然是黄花棘豆,矮嵩草次之,扁蓿豆最低。Pmax变化趋势不同于禁牧地,黄花棘豆位居第一,其他3种草相比仍然是扁蓿豆>矮嵩草>垂穗披碱草。这说明在放牧之后,扁蓿豆仍然有较大的光合潜力,垂穗披碱草的光合能力仍较弱。
4种植物在放牧和禁牧两种条件下光响应参数的变化不同(图1、表1)。在NG中,当光强超过LSP后,垂穗披碱草和矮嵩草的Pn有下降趋势,而在G中这2种植物这种趋势不明显,黄花棘豆却明显。扁蓿豆的Pn在2种草地中均无此趋势。垂穗披碱草、矮嵩草、扁蓿豆在G中的Pmax显著低于NG(P<0.05),而黄花棘豆在G中的Pmax却显著高于NG(P<0.05)。垂穗披碱草在NG中的LCP高于G(P<0.05),LSP低于 G,同黄花棘豆;矮嵩草在NG中的LCP和LSP都低于G(P<0.05);扁蓿豆在NG中的LCP高于G(P<0.05),而LSP变化不大。垂穗披碱草、黄花棘豆、扁蓿豆在NG中的暗呼吸速率(Rd)高于 G(P<0.05),而矮嵩草高于 G(P<0.05)。4种植物LCP的变化规律与Rd一致,矮嵩草在NG中的低于G,其他3种植物在NG中的高于G中。垂穗披碱草和扁蓿豆在NG中的表观量子效率显著高于G中(P<0.05),而矮嵩草在G中的表观量子效率显著高于NG中(P<0.05),黄花棘豆的表观量子效率没有显著差异。
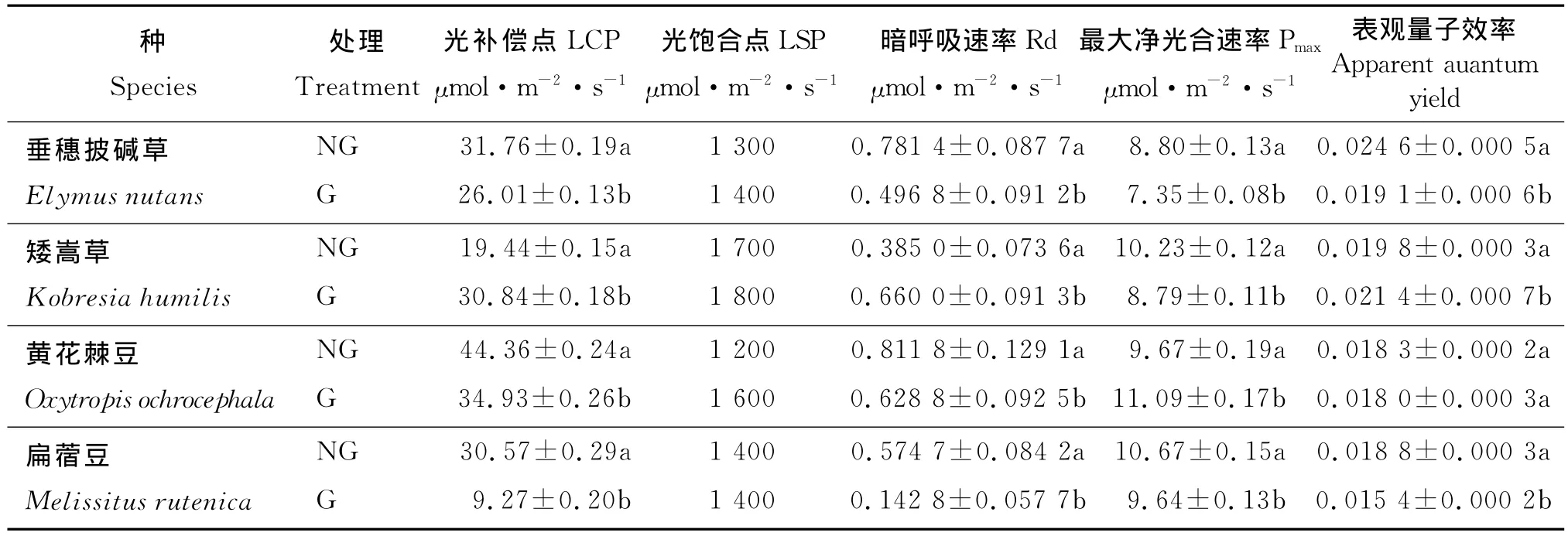
表1 放牧和禁牧草地中4种优势植物净光合速率对光强响应参数表Table 1 Pn-PAR curve parameters of four dominant plants in the grazing and Non-grazing grasslands
2.2 放牧对叶绿素荧光参数的影响
2.2.1 对叶绿素初始荧光(F0)的影响 F0是所有PSⅡ的反应中心全部处于开放态时的荧光产量,在一定程度上与色素含量呈线性关系,即与叶绿素的含量有关。叶绿素接受光后可发射暗红色的荧光信号,而逆境可使这种信号的颜色变暗[19],也就是说颜色加深说明叶片即受到了胁迫。垂穗披碱草和矮嵩草在NG中叶片发出橙红色荧光,颜色均匀一致;而在G中,叶片荧光颜色加深至暗红色,有些部位荧光几乎不可见,说明叶绿素含量已明显降低(图2)。扁蓿豆在G中颜色有局部加深的迹象,但与NG相比差异不明显,说明叶绿素含量有所降低。黄花棘豆的颜色无明显变化。

图2 放牧和禁牧草地中4种优势植物叶绿素初始荧光(F0)成像Fig.2 Image of minimal fluorescence(F0)of four dominant plants in the grazing and Non-grazing grasslands
2.2.2 对 PSⅡ 最大量子产量(Fv/Fm)的影响Fv/Fm在正常条件下变化极小,不受物种和生长条件的影响,但在逆境下该参数明显下降[19]。矮嵩草、黄花棘豆、扁蓿豆3种植物在NG中的Fv/Fm值高于G中,差异显著(P<0.05)。而垂穗披碱草在NG中的Fv/Fm值高于G中(图3)。
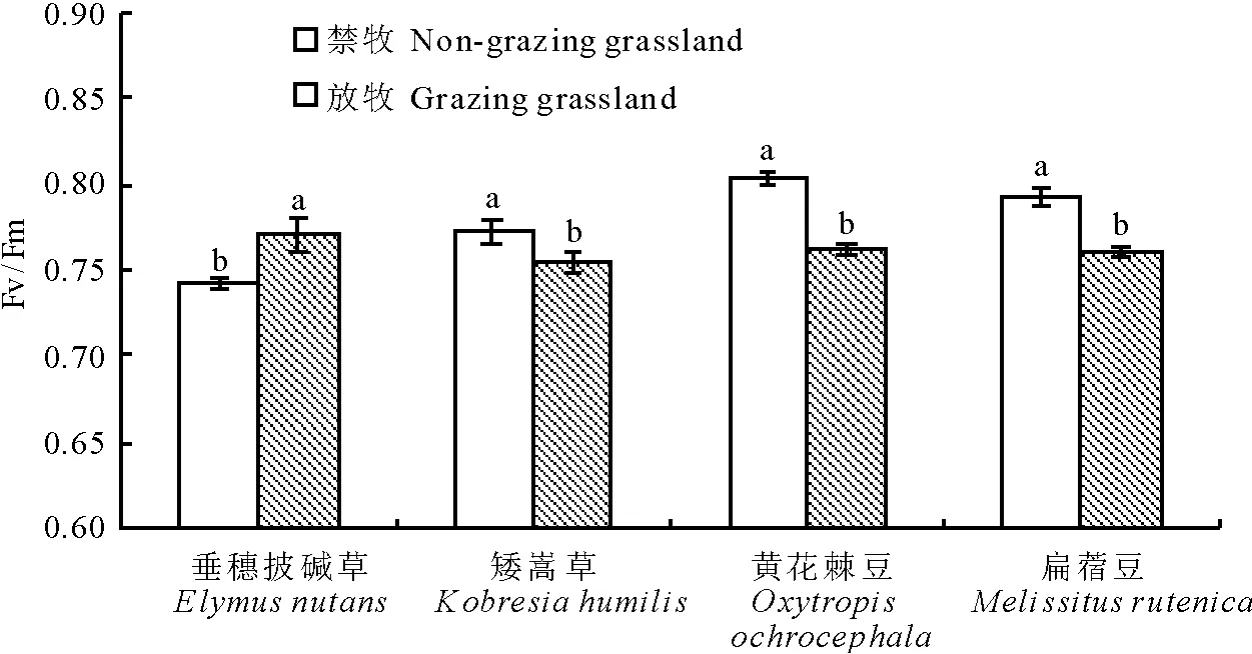
图3 放牧和禁牧草地中4种优势植物叶片PSⅡ最大量子产量(Fv/Fm)的对比Fig.3 Maximal quantum yield of PSⅡ(Fv/Fm)of four dominant plants in the grazing and Non-grazing grasslands

图4 放牧和禁牧草地中4种优势植物叶片光化学猝灭系数(qP)的对比Fig.4 Photochemical quenching coefficient(qP)of four dominant plants in the grazing and Non-grazing grasslands
2.2.3 对光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)的影响 叶绿素荧光猝灭是叶绿体耗散能量的一种途径[19]。分为光化学猝灭和非光化学猝灭。光化学淬灭是由光合作用引起的荧光淬灭,反映了植物光合活性的高低。总体来看(图4),垂穗披碱草和矮嵩草在NG中的光化学猝灭系数(qP)明显高于在G中。黄花棘豆的qP也是NG中的高于G中。扁蓿豆的qP在0~100s时NG中的高于G中,而100s以后则无显著差异。
非光化学猝灭是指由热耗散引起的荧光淬灭,是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分,是一种自我保护机制,对光合机构具有一定的保护作用[20]。垂穗披碱草、矮嵩草、扁蓿豆在G中的非光化学猝灭系数(NPQ)高于在NG;而黄花棘豆在60~140s时间段内NG高于G,这与G中的Pmax高于NG相一致;但140s以后G又高于NG(图5)。总体来看,G中要高于NG中的,这与qP的变化规律相反。
2.2.4 对PSⅡ的电子传递速率(ETR)的差异 ETR反映了实际光强条件下的表观电子传递效率[21]。在胁迫下,植物叶片的ETR会下降[19]。总体来看(图6),4种植物在NG中的ETR均高于G中。矮嵩草和黄花棘豆40~80s时间段,G中的ETR高于NG,这可能与这2种植物的内在调节机制有关,其机理尚不明确。
3 讨论

图5 放牧和禁牧草地中4种优势植物叶片非光化学猝灭系数(NPQ)的对比Fig.5 Contrasts of the non-photochemical quenching coefficient(NPQ)of four kinds of dominant plants in the grazing and Non-grazing grasslands

图6 放牧和禁牧草地中4种优势植物叶片表观电子传递速率(ETR)的对比Fig.6 Contrasts of the electron transport rate(ETR)of four kinds of dominant plants in the grazing and Non-grazing grasslands
植物的光响应曲线是光合作用随着光照强度改变的系列反应曲线,通过曲线可以计算并判断植物的最大(净)光合速率、表观量子效率、暗呼吸速率、光饱和点及光补偿点等,是研究植物光合生理生态过程对环境响应的基础。综合来看,在禁牧和放牧两种草地中,矮嵩草的光饱和点最高,这是因为矮嵩草表皮细胞具有较厚的角质层和蜡质层,并具有发达的栅栏组织,有利于提高光合效率,对强辐射的生态环境有很好的适应性[22]。黄花棘豆的光补偿点最高,这说明黄花棘豆对弱光的利用能力较其他3种植物都弱,而对强光的适应能力较强,能较好的适应强辐射环境。垂穗披碱草的光补偿点、光饱和点和最大净光合速率都很低,扁蓿豆的都较高,说明在这4种草中,垂穗披碱草对强光的适应能力最弱,但能有效利用弱光;而扁蓿豆在较强的光辐射下才能发挥出光合潜力,且具有较强的光合能力。然而,这2种植物中的光合光响应曲线有所不同。在禁牧中,垂穗披碱草和矮嵩草有光抑制的趋势,而放牧中没有光抑制现象。垂穗披碱草、矮嵩草、扁蓿豆在放牧样地中的最大净光合速率显著低于禁牧样地(P<0.05),而黄花棘豆在放牧中的最大净光合速率却显著高于禁牧(P<0.05)。这可能是因为在重牧下,垂穗披碱草、矮嵩草、扁蓿豆作为家畜喜食的牧草,地上部分大部分被采食,整个植株的生长受到影响,导致光合能力下降;放牧导致禾草和嵩草等的比例减少,黄花棘豆作为一种典型的毒草,由此获得了一定的滋生空间[23],叶片的接受光面积增大,光合能力增强。放牧使得4种植物的光饱和点升高(扁蓿豆不显著),矮嵩草的光补偿点、暗呼吸速率也升高,使得垂穗披碱草、黄花棘豆、扁蓿豆的光补偿点、暗呼吸速率都降低。说明放牧提高了这4种植物对强光的适应能力,同时提高了垂穗披碱草、黄花棘豆、扁蓿豆对弱光的利用能力。
叶绿素荧光动力学技术在测定叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用,被称为光合作用研究的有效探针。常乐等[15]研究了4种园林地被植物在受到践踏胁迫后叶绿素荧光特性的变化,证明叶绿素荧光参数能很好地反映植物受到的胁迫情况。在放牧地中,植株受到的损伤,除了肉眼可以看出,叶绿素荧光图像也可以明确指出植株受到的伤害程度。图1显示出垂穗披碱草、矮嵩草和扁蓿豆已经受到了伤害。
对4种植物的最大量子产量、光化学淬灭、非光化学淬灭、电子传递的相对速率等参数的分析结果显示,除垂穗披碱草的最大量子产量值在放牧后升高外(机理尚待进一步试验研究),矮嵩草、黄花棘豆、扁蓿豆3种植物在禁牧中的最大量子产量值高于放牧中,说明这3种植物的光能转换效率下降,已明显受到胁迫。4种植物的光化学淬灭是禁牧高于放牧(扁蓿豆的光化学淬灭差异不显著),非光化学淬灭的变化正好相反,说明在放牧时光合器官明显受损,光合活性降低,有较多的激发能以热的形式散发,虽然光能利用率较禁牧地低,但是在长时间的逆境中,形成了更强的光破坏保护能力。黄花棘豆在60~140s时间段内禁牧高于放牧,这与放牧中的最大净光合速率高于禁牧相一致,进一步说明在黄花棘豆放牧中有着比在禁牧中更强的光合能力。电子传递的相对速率也是禁牧高于放牧,这说明放牧条件下植物叶片的电子传递受到一定的抑制,进而影响到光合作用的正常进行。
[1] 张如力,张如清,肖云峰.绢蝶在祁连山(北坡)寺大隆林牧区的垂直分布及物种多样性[J].草业科学,2005,22(9):9-13.
[2] 刘金荣,谢晓蓉.祁连山高寒草地特点及可持续发展利用对策[J].草原与草坪,2002(2):15-16.
[3] 蒋高明,何维明.一种在野外自然光照条件下快速测定光合作用光响应曲线的新方法[J].植物学报,1999,16(6):712-718.
[4] Sharp R,Matthews M,Boyer J K.Effect and the quantum yield of photosynthesis[J].Plant Physiology,1984(75):95-101.
[5] 田玉强,高琼,张智才,等.青藏高原高寒草地植物光合与土壤呼吸研究进展[J].生态环境学报,2009,18(2):711-721.
[6] 张德罡,曹文侠,蒲小鹏,等.东祁连山杜鹃属植物光合作用特点的研究[J].甘肃农业大学学报,2003,38(2):140-143.
[7] 高丽楠.青藏高原东缘高寒草甸优势植物光合作用日变化的比较研究[D].成都:四川师范大学,2008:7-32.
[8] Pandey S,Kumar N,Kushwaha R.Morpho-anatomical and physiological leaf traits of two alpine herbs,Podophyllum hexandrum and Rheum emodi in the Western Himalaya under different irradiances[J].Photosynthetica,2006,44(1):11-16.
[9] Christina K,Sonja G,Katrin T,et al.Photosynthetic efficiency as a function of thermal stratification and phytoplankton size structure in an oligotrophic alpine lake[J].Hydrobiologia,2007,578(1):29-36.
[10] 贾宏涛,赵成义,盛钰,等.干旱地区紫花苜蓿光合日变化规律研究[J].草业科学,2009,26(7):56-60.
[11] 莫凌,黄玉清,桂堂辉,等.5个热带禾本科牧草品种的光
合-光响应特性研究[J].草业科学,2010,27(9):64-68.
[12] Chen S P,Bai Y F,Zhang L X,et al.Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin o f China[J].Environmental and Experimental Botany,2005(53):65-75.
[13] 赵鸿,王润元,郭铌,等.禁牧对安西荒漠化草原芨芨草光合生理生态特征的影响[J].干旱气象,2007,25(1):63-66.
[14] 闫瑞瑞,卫智军,运向军,等.放牧制度对短花针茅荒漠草原主要植物种光合特性日变化影响的研究[J].草业学报,2009,18(5):160-167.
[15] 常乐,夏宜平,楼建华,等.践踏胁迫对4种园林地被植物叶绿素荧光特性的影响[J].园艺学报,2010,37(10):1673-1678.
[16] 王爱东,尚占环,鱼小军,等.东祁连山北坡高寒灌丛草地围栏与放牧干扰下CO2释放速率的比较研究[J].甘肃农业大学学报,2010,45(1):120-124.
[17] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].农业基础科学,2005,2(8):76-79.
[18] 徐伟洲,徐炳成,段东平,等.不同水肥条件下白羊草光合生理生态特征研究Ⅱ.光响应曲线[J].草地学报,2010,18(6):773-779.
[19] 陈建明,俞晓平,程家安.叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J].浙江农业学报,2006,18(1):51-55.
[20] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1996,16(4):444-448.
[21] 施征,史胜青,肖文发,等.脱水胁迫对梭梭和胡杨苗叶绿素荧光特性的影响[J].林业科学研究,2008,21(4):566-570.
[22] 崔艳.青藏高原东部高草属植物的生态解剖学研究[D].兰州:西北师范大学,2006:1-45.
[23] 王小利,张力,张德罡,等.青海湖地区线叶嵩草型中度与重度退化草地群落的比较研究[J].草业科学,2006,23(10):8-13.
猜你喜欢
草业学报(2022年11期)2022-11-23
美文(2022年18期)2022-10-20
现代畜牧科技(2021年7期)2021-07-28
西藏农业科技(2021年4期)2021-04-18
种子(2018年11期)2018-12-05
中成药(2018年3期)2018-05-07
天然产物研究与开发(2014年3期)2014-04-27
影像科学与光化学(2014年5期)2014-03-11
影像科学与光化学(2014年5期)2014-03-11
无机化学学报(2014年3期)2014-02-28
