
饲粮中添加竹青素对肉鸡脂质代谢的影响
2013-09-20 00:33黄骆镰龚凌霄吴晓琴
动物营养学报 2013年1期
黄骆镰 龚凌霄 刘 聪 吴晓琴 张 英
(浙江大学生物系统工程与食品科学学院,杭州 310058)
竹子是禾本科竹亚科多年生常绿植物,种植面积广,是中国传统的药食同源的植物之一。竹青素(green bamboo extract,GBE)是由竹叶、竹茹、竹笋提取物复配而成的一种新型饲料添加剂,是一种卓越的生物抗氧化剂、脂质代谢调节剂和免疫功能增强剂。GBE内含丰富的竹子次生代谢产物,主要包括黄酮类化合物、酚酸类化合物、生物活性多糖、氨基酸肽类、萜类内酯、蒽醌类以及矿物质元素等,目前已在加州鲈鱼、大黄鱼的养殖上取得了良好的效果[1]。
鸡肉是人们餐桌中受欢迎的菜肴之一,消费量巨大。体内脂肪过多是肉鸡养殖中普遍存在的问题。鸡的腹脂和皮下脂肪沉积过多,影响了鸡的胴体品质,不受消费者欢迎;沉积单位脂肪比沉积单位瘦肉多消耗3倍的能量,体脂沉积过多会降低饲料利用率[2];体脂过多还严重影响鸡肉的加工性能。目前,通过营养调控途径减少家禽脂肪沉积、改善家禽胴体品质已成为动物营养学研究的热点。大量研究已证明,竹子提取物具有良好的调节脂质代谢的功能,Panee等[3]通过体外试验研究证实竹子提取物具有抗脂毒功能;Sun等[4]研究表明竹叶中的黄酮类化合物可以阻碍脂肪的形成;Higa等[5]研究认为炎症与肥胖密切相关,竹叶提取物可以通过降低单核细胞趋化蛋白-1(MCP-1)的含量达到减少小鼠腹部脂肪的作用。结合竹子资源产量高、低成本等特点,GBE具有作为脂质代谢调节剂应用于禽类生产的潜力。而目前,竹子提取物制品在禽类生产中的应用尚未见报道。石岐胡须鸡(商品名:泰安鸡)是优质肉鸡品种,肉质鲜嫩,深受消费者欢迎,而脂肪沉积严重成为影响其生产、加工的核心问题。本试验通过在饲粮中添加一定比例的GBE饲喂石岐胡须鸡,研究GBE对肉鸡脂质代谢的影响,旨在为竹子提取物在肉鸡生产中的应用以及绿色无公害饲料添加剂的开发提供理论与实践依据。
1 材料与方法
1.1 试验材料
GBE,由杭州尤美特科技有限公司提供,含总酚14.25%、多糖11.06%、总三萜4.20%。
1.2 试验设计
选取360只26日龄健康石岐胡须鸡仔鸡(由香港现代农业有限公司提供),初始体重为(375.12±8.30)g,随机分成4组,每组3个重复,每个重复30只鸡。对照组饲喂不添加GBE的基础饲粮,基础饲粮为粉状配合饲料,参考美国NRC(1994)肉鸡营养需要配制,其组成及营养水平见表1。试验组饲喂在基础饲粮中分别添加1.5、3.0和6.0 g/kg GBE的试验饲粮。试验期49 d。
1.3 饲养管理
试验在某养殖场进行,试验前对鸡舍、鸡笼、料槽、喂水器等进行清洗消毒。实行笼养,每笼2只,笼大小为35 cm×35 cm ×35 cm。鸡舍温度为22~27℃,自然采光,保持鸡舍通风、干燥、卫生。肉仔鸡自由采食,充足饮水,按正常免疫程序进行免疫接种。
1.4 样品采集
试验结束后禁食12 h(自由饮水),每个重复随机选取3只鸡称重,活体翅静脉采血,离心收集血清,分装至-20℃冰箱备用;宰杀后分离腹脂、肝脏,分别称重,存于-20℃冰箱备用。
1.5 测定指标及方法
1.5.1 脂肪沉积指标测定
腹脂率:剥离腹部脂肪和肌胃周围脂肪,称重,计算公式如下:
腹脂率(%)=(腹脂重/体重)×100。
皮下脂肪厚度:测定部位在尾椎前端、背正中线切开的皮肤转角处,用游标卡尺测量。
肝脏脂肪含量:按 GB/T 9695.7—2008[6]测定。
肝脏冷冻切片结合油红“O”染色观察:取肝脏冰冻切片(厚度8 μm)固定后水化,置60%异丙醇中漂洗30 s,油红“O”储备液与蒸馏水按6∶4比例混合,静止10 min,切片染色10 min,用60%异丙醇稍微洗去多余染液,再用蒸馏水漂洗,Mayer苏木素复染1 min,0.5%盐酸酒精分化5 s,流水冲洗1 min,0.5%稀氨水蓝化20 s,流水冲洗1 min,甘油明胶封固。每个肝脏样本切2套冰冻切片,以减少病理染色因素对试验的影响。显微镜400倍下观察、拍照。
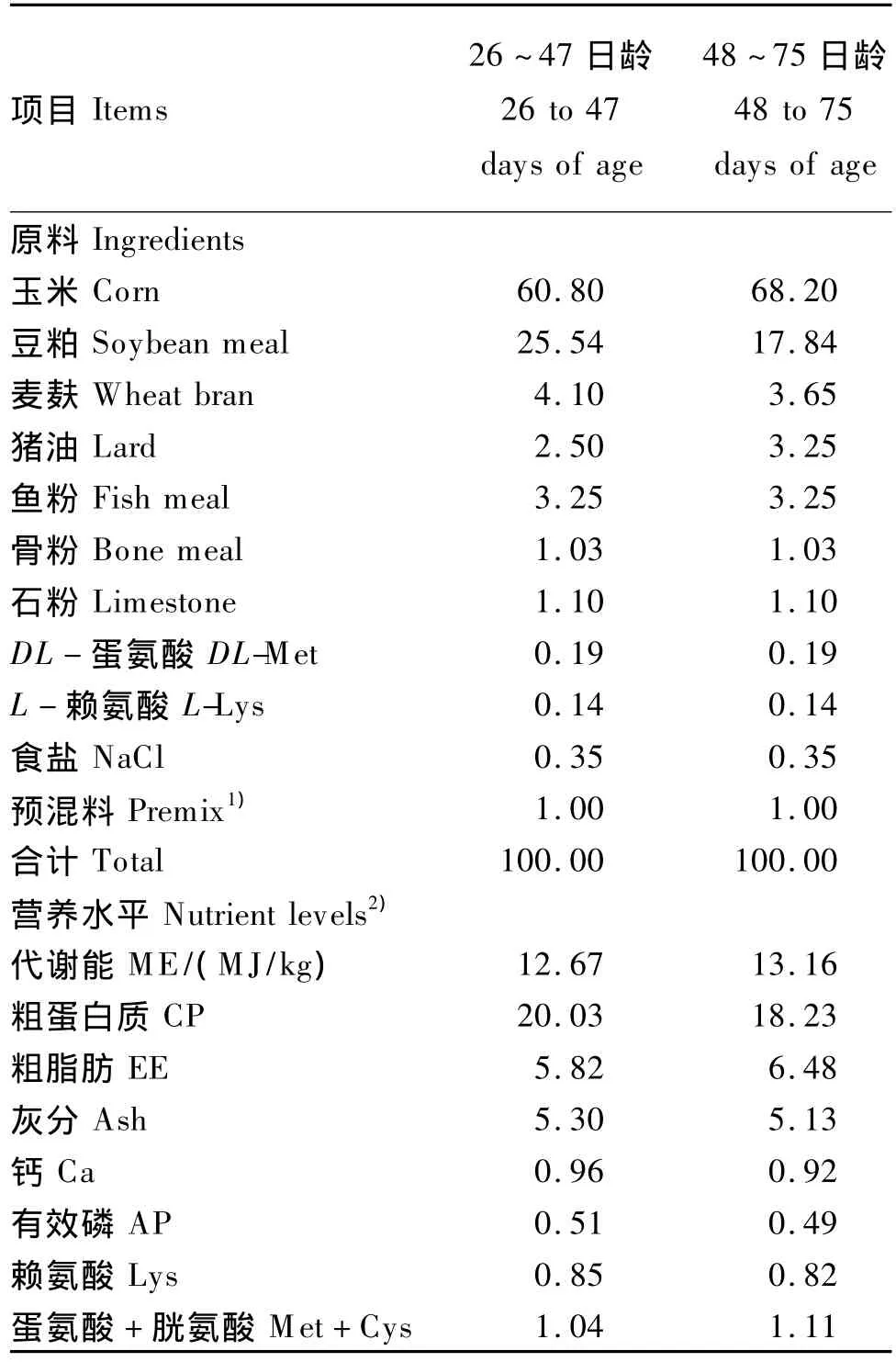
表1 基础饲粮组成及营养水平(风干基础)Table1 Composition and nutrient levels of basal diets(air-dry basis) %
1.5.2 血清生化指标测定
测定时取血清,流水解冻。总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)、瘦素、脂联素、丙二醛(MDA)含量,超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)活性均由南京建成生物工程研究所提供的试剂盒测定。
1.6 数据处理
试验数据采用SPSS 17.0软件进行统计学分析。结果用平均值±标准差表示,显著性检验采用单因素方差分析(one-way ANOVA),采用LSD方法进行多重比较,以P<0.05时为差异显著,以P<0.01时为差异极显著。
2 结果
2.1 饲粮中添加GBE对肉鸡脂肪沉积的影响
由表2可知,与对照组相比,饲粮中添加1.5、3.0和6.0 g/kg GBE肉鸡体重均略有下降,但影响不显著(P>0.05);饲粮中添加1.5和3.0 g/kg GBE显著降低了肉鸡腹脂率(P<0.05),而添加6.0 g/kg GBE则对腹脂率无显著影响(P>0.05);饲粮中添加GBE可降低肉鸡皮下脂肪厚度,其中1.5和6.0 g/kg组达极显著水平(P<0.01),3.0 g/kg组达显著水平(P<0.05);饲粮中添加1.5 g/kg GBE对肉鸡肝脏脂肪含量无显著影响(P>0.05),添加3.0和6.0 g/kg GBE显著降低了肝脏脂肪含量(P<0.05)。

表2 饲粮中添加GBE对肉鸡脂肪沉积的影响Table2 Effects of dietary GBE on fat deposition of broilers
由图1可知,饲粮中添加1.5、3.0和6.0 g/kg GBE,肉鸡肝脏脂肪滴(红色部分)的分布面积随着GBE添加水平的增加而逐渐降低。

图1 肉鸡肝脏冷冻切片结合油红“O”染色Fig.1 Frozen sections of liver in broilers by oil red-O staining(400×)
2.2 饲粮中添加GBE对肉鸡血脂的影响
由表3可知,与对照组相比,饲粮中添加1.5、3.0和6.0 g/kg GBE对肉鸡血清TC含量均无显著影响(P>0.05);饲粮中添加GBE可降低肉鸡血清TG含量,其中1.5和6.0 g/kg组达显著水平(P<0.05),3.0 g/kg组达极显著水平(P<0.01);饲粮中添加1.5和6.0 g/kg GBE对肉鸡血清HDL-C含量无显著影响(P>0.05),添加3.0 g/kg GBE显著提高了血清HDL-C含量(P<0.05);饲粮中添加1.5、3.0和6.0 g/kg GBE对肉鸡血清LDL-C含量均无显著影响(P>0.05)。

表3 饲粮中添加GBE对肉鸡血脂的影响Table3 Effects of dietary GBE on blood lipids of broilers mg/dL
2.3 饲粮中添加GBE对肉鸡血清瘦素和脂联素含量的影响及其与脂肪沉积指标之间的皮尔逊相关系数
由表4可知,与对照组相比,饲粮中添加1.5和3.0 g/kg GBE均显著提高了肉鸡血清中瘦素含量(P<0.05),添加6.0 g/kg GBE肉鸡血清中瘦素含量也有所提高,但差异不显著(P>0.05);饲粮中添加1.5和6.0 g/kg GBE对肉鸡血清中脂联素含量无显著影响(P>0.05),添加3.0 g/kg GBE显著提高了血清中脂联素含量(P<0.05)。

表4 饲粮中添加GBE对肉鸡血清瘦素和脂联素含量的影响Table4 Effects of dietary GBE on leptin and adiponectin contents in serum of broilers
由表5可知,肉鸡血清中瘦素含量与腹脂率、皮下脂肪厚度呈极强负相关;脂联素与肝脏脂肪含量呈极强负相关;血清瘦素含量与脂联素含量两者呈强正相关。

表5 瘦素和脂联素含量与脂肪沉积指标之间的皮尔逊相关系数Table5 Pearson correlation coefficients between leptin or adiponectin contents and fat deposition indices
2.4 饲粮中添加GBE对肉鸡内源性抗氧化酶系的促进及其对脂质过氧化的抑制作用
由表6可知,与对照组相比,饲粮中添加1.5和3.0 g/kg GBE极显著提高了肉鸡血清SOD活性(P<0.01),添加6.0 g/kg GBE显著降低了肉鸡血清SOD活性(P<0.05);饲粮中添加1.5和6.0 g/kg GBE对肉鸡血清中GSH-Px活性无显著影响(P>0.05),添加3.0 g/kg GBE极显著提高了肉鸡血清中GSH-Px活性(P<0.01);饲粮中添加1.5、3.0和6.0 g/kg GBE对肉鸡血清CAT活性均无显著影响(P>0.05);饲粮中添加1.5和3.0 g/kg GBE极显著降低了肉鸡血清MDA含量(P<0.01),添加6.0 g/kg GBE对血清MDA含量无显著影响(P>0.05)。

表6 饲粮中添加GBE对肉鸡内源性抗氧化酶系及脂质过氧化的影响Table6 Effects of dietary GBE on endogenous antioxidant enzyme system and lipid peroxidation of broilers
3 讨论
3.1 饲粮中添加GBE对肉鸡脂肪沉积的影响
腹部是鸡体内蓄积脂肪的主要部位,腹脂和胴体脂肪呈中等程度以上的表型相关和遗传相关,因此,腹脂率和皮下脂肪厚度在一定程度上反映了动物体内脂肪代谢状况[7]。在本试验中,随着饲粮中GBE添加水平的提高,肉鸡的腹脂率呈现先降低后提高的趋势,其中以3.0 g/kg组的腹脂率最低,达显著水平,说明GBE对肉鸡腹脂率的影响存在双向调节的作用。饲粮中添加GBE均能显著降低肉鸡的皮下脂肪厚度,其中1.5和6.0 g/kg组达极显著水平,该结果与GBE对腹脂率的影响趋势不完全一致,可能是肉鸡的腹腔脂肪沉积与皮下脂肪沉积过程是相对独立的过程。GBE 3个添加水平对肉鸡的体重均无显著影响。由此可见,饲粮中添加一定比例的GBE,可降低石岐胡须鸡的体脂沉积,但并不影响鸡的生长性能,本试验很好地解决了石岐胡须鸡脂肪沉积严重的问题。GBE降低肉鸡脂肪含量的作用可能与其含丰富的黄酮类化合物有关,李莉等[8]在饲粮中添加山楂叶总黄酮喂养黄羽肉鸡,发现腹脂率和皮下脂肪率均显著降低,与本试验结果一致。
鸡的脂肪代谢及其调控特点与哺乳类动物不同,鸡脂肪酸合成的主要场所是肝脏[9],鸡体脂沉积所需的脂肪酸大多来自于肝脏合成的脂肪酸[10],因此,鸡肝脏脂肪含量也是反映鸡脂肪代谢状况的重要指标。饲粮中添加1.5 g/kg GBE对肉鸡肝脏脂肪含量无显著影响,而添加水平达3.0和6.0 g/kg时肝脏脂肪含量显著降低。通过鸡肝脏冷冻切片结合油红“O”染色400倍拍摄图片可知,肝脏脂肪滴(红色部分)的分布面积随着GBE添加水平的增加而逐渐降低,与其肝脏脂肪含量检测结果相对应。由此可见,饲粮中添加GBE可以明显降低石岐胡须鸡的肝脏脂肪含量,并呈一定的剂量效应,说明竹青素要起到降低肝脏脂肪沉积的作用必须达到一定的水平。
3.2 饲粮中添加GBE对肉鸡血脂的影响
脂肪沉积及代谢是一个复杂的生理生化过程,受饮食、内分泌、神经及脂肪组织自身等多因素的调控。动物体内脂肪代谢始终处于动态平衡之中,在动物体内脂类代谢中,肝脏、脂肪组织和血浆中的脂质水平是相互联系、相互影响的[8]。研究表明,植物提取物对动物的血脂具有调节作用,谢棒祥等[11]研究发现山楂叶总黄酮和大豆黄酮可显著降低爱拔益加(AA)肉鸡血清中TC、TG含量;Kirk等[12]报道,大豆黄酮可能通过增加低密度脂蛋白受体活性来降低血浆中的TC含量。在本试验中,饲粮中添加1.5、3.0和6.0 g/kg GBE能显著降低肉鸡血清中TG含量,其中3.0g/kg组达极显著水平。家禽脂肪组织发育、脂肪沉积取决于血浆中TG的含量[13],因此,TG含量降低可一定程度上减少肉鸡的体脂沉积。HDL-C主要功能是转移细胞膜中的胆固醇,有利于保护心脑血管,添加3.0 g/kg GBE可显著提高血清HDL-C含量。
3.3 瘦素和脂联素对肉鸡脂质代谢的调控
瘦素是由脂肪细胞合成并分泌的具有多种功能的肽类激素,可通过特异性受体及多重神经肽通路作用于中枢神经系统,抑制食欲,增加能量消耗,降低脂肪沉积,对肥胖过程起拮抗和抑制的作用。瘦素作为负反馈调节信号,参与体脂的负调节,它在稳定能量平衡方面发挥基础性作用[14]。天然植物提取物可通过提高禽类动物的瘦素含量来参与脂质代谢调节,李莉等[8]研究认为,山楂叶总黄酮可显著提高黄羽肉鸡血清中的瘦素含量,从而降低皮下脂肪率和腹脂率。脂联素是一种由脂肪细胞特异性分泌的蛋白质,在血浆中表达浓度相对较高。脂联素在脂肪细胞分化过程中形成,并经胰岛素刺激而分泌,与特定受体中的类胶原功能区结合而影响脂质合成、储运及机体代谢[15]。研究表明,脂联素在调节糖代谢、脂代谢、抗炎等方面起着重要的生理作用[16]。本试验中,饲粮中添加GBE可提高肉鸡血清中的瘦素含量,并且随着添加水平的提高,瘦素含量先提高后降低,其中以3.0 g/kg组的瘦素含量最高,达显著水平,说明GBE对肉鸡血清中瘦素含量的影响存在双向调节的作用。饲粮中添加GBE对肉鸡血清中脂联素的影响趋势与瘦素一致,添加水平为3.0 g/kg时达显著水平。
由瘦素、脂联素含量与脂肪沉积指标的相关性分析可知,血清中瘦素含量与腹脂率、皮下脂肪厚度呈极强负相关;血清脂联素含量与肝脏脂肪含量呈极强负相关;血清瘦素含量与脂联素含量两者呈强正相关。由此可见,瘦素、脂联素含量与脂肪沉积指标呈较高的相关度。
3.4 饲粮中添加GBE对肉鸡内源性抗氧化酶系的促进及其对脂质过氧化的抑制作用
在生物体中自由基为细胞代谢过程中连续不断产生的具有高度活性的物质,一旦机体内清除氧自由基的能力下降,氧自由基浓度升高,便可以对生物膜的不饱和脂质酸诱发产生氧化反应,导致细胞膜、核酸、蛋白质和酶类等发生不可逆的损伤[17]。氧自由基攻击生物膜中的多不饱和脂肪酸引发脂质过氧化反应,形成脂质过氧化产物MDA,因此,MDA的含量常常可反映机体内脂质过氧化的程度,间接地反映出细胞损伤的程度。研究表明,脂质代谢紊乱与抗氧化应激存在密切的关系[18]。在肥胖动物模型中,已发现抗氧化物质防御机制的改变,Beltowski等[19]证实肥胖大鼠伴随着SOD活性的降低;Capel等[20]在肥胖小鼠中发现GSH-Px活性下降。临床观察中也有类似的发现,Ozata等[21]研究表明,肥胖者的铜锌超氧化物歧化酶(CuZn-SOD)和GSH-Px活性极显著低于正常者。目前认为SOD与GSH-Px共同组成消除自由基的酶系防御体系。本试验中,饲粮中添加1.5和3.0 g/kg GBE极显著提高了肉鸡血清SOD活性,而添加水平达6.0 g/kg时,SOD活性反而显著降低,说明GBE对SOD活性影响是双向调节的过程,当添加水平较高时,反而对SOD活性产生抑制作用。3个添加水平均能不同程度地提高血清GSH-Px活性并降低MDA的含量。由此可见,GBE可显著提高肉鸡血清中主要抗氧化酶的活性,减轻体内脂质过氧化反应,从而改善脂质代谢紊乱。
4 结论
①饲粮中添加不同水平的GBE,可显著降低肉鸡的体脂沉积,但不影响鸡的体重,其中以3.0 g/kg添加水平效果最佳。
②饲粮中添加GBE,肉鸡血清中瘦素、脂联素含量和抗氧化酶活性均显著提高,并与脂肪沉积指标呈较高相关度,结合其生物学作用提示瘦素、脂联素和抗氧化酶可能参与肉鸡脂质代谢的调控。
致谢:
感谢香港现代农业有限公司对本研究提供条件支持以及技术指导。
[1]干钢,金成,张高立,等.强化尤美肝泰的人工配合饲料对加州鲈生长及品质的影响[C]//中国环境科学学会2010年学术年会论文集.上海:中国环境科学学会,2010:4312-4317.
[2]CHAMBERS J R,GAVORA J S,FORTIN A.Genetic changes in meat-type chickens in the last twenty years[J].Canadian Journal of Animal Science,1981,61(3):555-563.
[3]PANEE J,LIU W Y,LIN Y L,et al.A novel function of bamboo extract in relieving lipotoxicity[J].Phytotherapy Research,2008,22(5):675-680.
[4]SUN J,YUE Y,TANG F,et al.Simultaneous HPTLC analysis of flavonoids in the leaves of three different species of bamboo[J].Journal of Planar Chromatography,2010,23(1):40-45.
[5]HIGA J K,LIU W Y,BERRY M J,et al.Supplement of bamboo extract lowers serum monocyte chemoattractant protein-1 concentration in mice fed a diet containing a high level of saturated fat[J].British Journal of Nutrition,2011,106(12):1810-1813.
[6]中华人民共和国国家质量监督检验检疫总局.GB/T 9695.7—2008肉与肉制品总脂肪含量测定[S].北京:中国标准出版社,2008.
[7]CAHANER A,NITSAN Z,NIR I.Weight and fat content of adipose and nonadipose tissues in broilers selected for or against abdominal adipose tissue[J].Poultry Science,1986,65(2):215-222.
[8]李莉,朱晓彤,束刚,等.日粮中添加山楂叶总黄酮对黄羽肉鸡脂肪代谢的影响[J].江西农业大学学报,2009,31(4):610-615.
[9]GOODRIDGE A G.Citrate-cleavage enzyme,malic enzyme and certain dehydrogenases in embryonic and growing chicks[J].Biochemical Journal,1968,108(4):663-666.
[10]SMITH S,WITKOWSKI A,JOSHI A K.Structural and functional organization of the animal fatty acid synthase[J].Progress in Lipid Research,2003,42(4):289-317.
[11]谢棒祥,张敏红,杜荣,等.类黄酮对肉仔鸡生产性能及脂质代谢的影响[J].动物营养学报,2002,14(4):49-53.
[12]KIRK E A,SUTHERLAND P,WANG S A,et al.Dietary isoflavones reduce plasma cholesterol and atheroclerosis in C57BL/6 mice but not LDL receptor-deficient mice[J].The Journal of Nutrition,1998,128(6):954-959.
[13]尹靖东,齐广海,霍启光.家禽脂类代谢调控机理的研究进展[J].动物营养学报,2000,12(2):1-7.
[14]BI S,LADENHEIM E E,SCHWARTZ G J,et al.A role for NPY overexpression in the dorsomedial hypothalamus in hyperphagia and obesity of OLETF rats[J].American Journal of Physiology:Regulatory,Integrative and Comparative Physiology,2001,281(1):254-260.
[15]HUG C,WANG J,AHMAD N S,et al.T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(28):10308-10313.
[16]KHARROUBI I,RASSCHAERT J,EIZIRIK D L,et al.Expression of adiponectin receptors in pancreatic β cells[J].Biochemical and Biophysical Research Communications,2003,312(4):1118-1122.
[17]刘耕陶.氧自由基损伤与抗氧化剂[J].生理科学,1988(2):83-84.
[18]OHARA Y,PETERSON T E,HARRISON D G.Hypercholesterolemia increasesendothelialsuperoxide anion production[J].The Journal of Clinical Investigation,1993,96(6):2546-2551.
[19]BELTOWSKI J,WOJCICKA G,GORNY D,et al.The effect of dietary-induced obesity on lipid peroxidation,antioxidant enzymes and total plasma antioxidant capacity[J].Journal of Physiology and Pharmacology,2000,51:883-896.
[20]CAPEL I D,DORRELL H M.Abnormal antioxidant defence in some tissues of congenitally obese mice[J].Biochemical Journal,1984,219(1):41-49.
[21]OZATA M,MERGEN M,OKTENLI C,et al.Increased oxidative stress and hypozincemia in male obesity[J].Clinical Biochemistry,2002,35(8):627-631.
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
实用老年医学(2021年10期)2021-12-05
中国饲料(2021年17期)2021-11-02
昆明医科大学学报(2021年3期)2021-07-22
广东饲料(2016年6期)2016-12-01
现代检验医学杂志(2016年4期)2016-11-15
中国民族医药杂志(2016年6期)2016-05-09
胃肠病学(2016年7期)2016-03-13
安徽医科大学学报(2015年9期)2015-12-16
