
TLR4蛋白在颞叶癫痫大鼠及患者脑组织中表达的改变及意义
2013-11-23 11:50刘希金殷亚萍杨志勇孙圣刚邓学军
中国神经免疫学和神经病学杂志 2013年3期
刘希金 殷亚萍 杨志勇 孙圣刚 邓学军
颞叶癫痫海马区存在明显的神经元丢失、胶质细胞增生和突触重建,被认为是导致颞叶癫痫的病理生理基础。在癫痫动物模型及颞叶癫痫患者脑组织中可见小胶质细胞和星形胶质细胞过度活化、神经元丢失,同时癫痫发作时,脑组织中肿瘤坏死因子(tumor necrosis factor,TNF)、环氧化酶-1(cydooxygenase-1,COX-1)、环氧化酶-2 (cydooxygenase-2,COX-2)、白细胞介素-1(interleukin-1,IL-1)等炎性反应因子表达亦相应增高,这些证据表明炎性反应介质可能在癫痫的发生中重要作用[1-3]。
Toll样受体4(TLR4)信号通路是一个信号转导家族,是介导免疫反应的主要信号通路,其主要分布于小胶质细胞和星形胶质细胞等免疫反应细胞的细胞膜上[4]。TLR4受体蛋白与相应的配体结合后,诱发下游信号核因子κB(nuclear factor κB,NFκB)的活化,进而诱导炎性反应因子的产生[5-6]。但是有关TLR4信号通路在癫痫发病机制作用中的研究相对较少[7-8]。
本研究通过检测大鼠颞叶癫痫模型海马及颞叶癫痫患者颞叶内侧脑组织中TLR4蛋白的表达变化,旨在探讨TLR4信号通路在颞叶癫痫发病机制中的可能作用。
1 材料和方法
1.1 动物及人脑组织 健康成年雄性SD大鼠100只,由华中科技大学同济医学院实验动物中心提供,体质量200~230g,标准环境(24~25℃,50%~60%湿度,12h昼夜交替)下饲养。该实验获得华中科技大学同济医学院动物伦理学批准。人脑组织样本由协和医院脑外科提供,患者知情并签署伦理学文件。共取得26份人脑组织,其中20份来自20例癫痫患者,且均有多种抗癫痫药物的暴露史,但癫痫控制欠佳,另6份来自6例脑外伤且无癫痫及癫痫药物暴露史的患者作为对照组。
1.2 方法
1.2.1 癫痫模型建立与分组:随机选取10只大鼠作为盐水对照组,其余大鼠建立癫痫模型。给予大鼠腹腔注射氯化锂(按体质量127mg/kg),17.5h后按体质量1mg/kg给予腹腔注射阿托品(购自中国上海禾丰制药有限公司),0.5h后按体质量15mg/kg给予腹腔注射毛果芸香碱(购自美国Sigma-Aldrich公司)。癫痫发作等级参照Racine标准[9],观察0.5h后若未达到4级及以上发作,则按体质量15mg/kg重复注射毛果芸香碱1次,最大剂量不超过60mg/kg,若至最大剂量仍无癫痫发作(无急性发作组)则停止注射。成功诱发急性发作的8只大鼠于急性期(3d内)处死(急性发作组),剩余大鼠按体质量10mg/kg给予腹腔注射地西泮(购自中国天津药业焦作有限公司)以终止急性发作,然后单笼饲养并观察,经(33±6.5)d潜伏期后,部分进入慢性自发性发作期(慢性自发性发作组),其余则为无慢性自发性发作组。
1.2.2 脑电图(EEG)监测:各组大鼠分别行EEG监测,每只大鼠分别监测2h。将大鼠麻醉固定,用细针分别插入双侧耳(参考电极A)、前额中央(frontal pole1,FP1;frontal pole2,FP2)、耳 下(temporal 1,T1;temporal 2,T2)、枕部(occipital 1,O1;occipital 2,O2),另一端接入脑电信号处理系统[10]。分析各组大鼠在相同时间(2h)内棘波(spike-wave discharges,SWDs)的数量[11],以棘波发放数目评估大鼠脑电情况。
1.2.3 免疫组化:大鼠经水合氯醛麻醉,打开胸腔,经心尖部插入灌流针,以生理盐水冲尽血液,待灌流液清亮,用多聚甲醛灌流液固定,断头取脑并置多聚甲醛液中浸泡,石蜡包埋,切片。将石蜡切片置于65℃烘箱中烘烤2h后用磷酸盐缓冲液(PBS)冲洗3次,每次5min。切片置于乙二胺四乙酸(EDTA)缓冲液中微波修复,中火至沸后断电,间隔10min低火至沸,自然冷却后以PBS洗3次,每次5min。切片放入3%(体积分数)过氧化氢溶液,室温下孵育10min,阻断内源性过氧化物酶,用PBS洗3次,每次5min,甩干后5%(质量浓度)牛血清白蛋白(BSA)封闭20min以封闭电荷,去除BSA液,每张切片加入50μL稀释的TLR4抗体(编号:ab22408,批号:76B357.1,购自美国Abcam公司)覆盖组织,4℃过夜,用PBS洗3次,每次5min,去除PBS液,每张切片加100μL二抗(武汉谷歌公司),4℃孵育50min,PBS洗3次,每次5min,去除PBS液,每张切片加50~100μL新鲜配制二氨基联苯胺(DAB)溶液,显微镜控制显色,显色完全后,蒸馏水冲洗,苏木素复染,1%(体积分数)盐酸酒精分化(1s),自来水冲洗,氨水返蓝,流水冲洗,切片经过梯度酒精脱水干燥,二甲苯透明,中性树胶封固。每张切片于相同光度及相同放大倍数(×400)下拍照,采用图像分析软件6.0(Image-Pro Plus 6.0)分析每张图片的平均吸光度,以平均吸光度表示TLR4蛋白的表达水平。
人脑组织的处理:经手术切除病灶取样后,于多聚甲醛液中浸泡24h,待标本充分固定后,石蜡包埋,切片。余操作同上。
1.3 统计学处理 数据输入SPSS V12.0数据处理软件(Chicago,IL,USA),计量资料以均数±标准差表示,多组间均数比较采用单因素方差(AVONA)分析,两两比较采用SNK检验;人脑数据采用t检验。以P<0.05表示具有统计学差异。
2 结果
2.1 大鼠行为学观察 大鼠腹腔注射毛果芸香碱10~30min后,63只出现癫痫急性发作,表现为流涎、结膜充血等反应,伴有点头、眨眼、咀嚼及湿狗样震颤,最后出现站立、跌倒或旋转,持续数分钟至半小时不等。发作后1h给予腹腔注射地西泮终止发作,其中8只未能有效终止发作而死亡。随机选取8只成功诱发癫痫急性发作的大鼠为急性发作组。17只诱发癫痫急性发作的大鼠经潜伏期后进入慢性自发性发作期,另30只大鼠在视频监测观察中未出现慢性自发性发作。潜伏期内大鼠食欲下降、消瘦,慢性自发性发作期可见大鼠间断出现全身抽搐、旋转,每次持续数秒钟至数分钟不等。
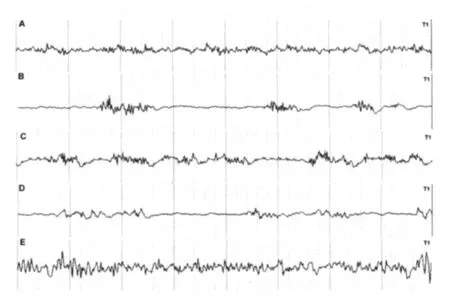
图1 各组大鼠EEG情况(T1导联)
2.2 EEG检测 在急性发作组及慢性自发性发作组大鼠均可见明显SWDs发放(图1),明显高于其他3组(P<0.05),且慢性自发性发作组SWDs数目高于急性发作组(P<0.05);盐水对照组未见SWDs,无急性发作组及无慢性自发性发作组偶见SWDs发放,且无急性发作组和无慢性自发性发作组比较无统计学差异(P>0.05),盐水对照组和其他4组相比有统计学差异(P<0.05)(表1)。
表1 各组大鼠EEG SWDs数目和CA3区TLR4蛋白水平 (,n=8)

表1 各组大鼠EEG SWDs数目和CA3区TLR4蛋白水平 (,n=8)
注:A:盐水对照组;B:无急性发作组;C:急性发作组;D:无慢性自发性发作组;E:慢性自发性发作组;与A组比较,*P<0.05;分别与B、D组比较,#P<0.05;与C组比较,△P<0.05;
组别 SWDs数量(个数) TLR4蛋白水平(A值)A 0.00±0.00 0.07858±0.01098 B 255.25±36.78* 0.08438±0.01422 C 723.00±94.11*# 0.13810±0.01264*#D 322.75±47.55* 0.08672±0.00701 E 1046.25±110.41*#△ 0.19650±0.01386*#△F值138.8384 105.862 P值 <0.001 <0.001
2.3 大鼠免疫组化结果 盐水对照组、无急性发作组、急性发作组、无慢性自发性发作组和慢性自发性发作组大鼠海马组织CA3区均有TLR4蛋白的表达(图2)。盐水对照组、无急性发作组、无慢性自发性发作组大鼠海马组织CA3区有少量TLR4蛋白表达,3组间比较无统计学差异(P>0.05);TLR4蛋白在急性发作组及慢性自发性发作组的表达均高于其他3组(P<0.05);慢性自发性发作组高于急性发作组(P<0.05)(表1)。
2.4 人脑免疫组化结果 TLR4蛋白在颞叶癫痫患者脑组织切片表达水平(0.79100±0.01750)高于对照组(0.15700±0.02588)(P<0.05)(图3)。
3 讨论
癫痫是一种中枢神经系统疾病,大多数患者经正规治疗后可有效控制癫痫发作,但有一部分患者未能有效控制仍可见频繁发作,临床上称为难治性癫痫,该部分绝大多数患者为颞叶癫痫[12]。癫痫的发病机制有多种学说,但是没有一种确切的机制能全面阐述癫痫[13]。

图2 各组大鼠CA3区TLR4蛋白表达(箭头所示,免疫组织化学×400)A:盐水对照组;B:无急性发作组;C:急性发作组;D:无慢性自发性发作组;E:慢性自发性发作组图3 TLR4蛋白在人脑组织切片中的表达(箭头所示,免疫组织化学×400)A:对照组;B癫痫组
近年来,国内外研究报道,炎性反应因子参与了癫痫的发病过程[1,14-15]。TLR4受体在免疫反应细胞的细胞膜上广泛表达。TLR4受体活化后,诱导下游的NF-κB进入细胞核,参与一系列的反应,促使炎性反应因子的释放和免疫反应细胞的增生与活化,而活化的免疫细胞释放更多的炎性反应因子,同时增生活化的星形胶质细胞功能缺陷,调节兴奋性氨基酸的功能降低,使神经元兴奋性增加,可能是导致癫痫的反复发作的病理机制。但是,有关于TLR4信号通路在癫痫发病中的研究报道较少[7-8]。
海马是大脑边缘系统的一部分,是易导致癫痫发生的敏感部位,这是颞叶癫痫发病的解剖学基础。以往的动物实验中,动物的分组往往忽略了无癫痫急性发作和无慢性自发性发作大鼠的实验数据并鲜有EEG数据,而这些实验数据可能对阐述疾病有着一定的意义。EEG可以为癫痫严重程度的评估提供比较客观的证据。增加无癫痫急性发作组,可表明研究指标与癫痫发作的因果关系,验证该指标是癫痫急性发作的因还是癫痫急性发作的果;癫痫作为一种较为特殊的疾病,有的患者经历一次痫性发作以后可能再无发作,有的患者在经历一次发作后会有更多的痫性发作,临床研究中很难获得只发作一次而再无发作患者的脑组织样本,而有癫痫急性发作但无慢性自发性发作的大鼠与仅有一次痫性发作的患者有类似的特点。因此,在实验研究中增加这两个动物分组也许能够更加详尽体现研究指标的变化趋势,揭示研究指标在不同时点发挥的作用,阐述其在颞叶癫痫的不同时期表达改变的意义。本实验通过建立动物颞叶癫痫实验模型并辅以EEG和视频观察,细化了动物模型分组,增加了无癫痫急性发作和有癫痫急性发作但无慢性自发性发作这两个动物分组,以求更加有效地阐述TLR4信号通路在颞叶癫痫发病机制中的作用。该研究结果显示,颞叶癫痫大鼠模型TLR4蛋白主要表达于CA1、CA3、齿状回和丘脑,在皮质也有一定的表达;TLR4蛋白在急性发作组和慢性自发性发作组表达增加,并且慢性自发性发作组高于急性发作组,而在其他各组则无统计学差异;并且TLR4蛋白的表达水平与EEG具有类似的变化趋势,提示TLR4信号通路在动物颞叶癫痫点燃后即有一定程度的表达上调,而急性发作期后的过度表达可能是癫痫反复发作的原因。
虽然动物模型可以在一定程度上模拟人类颞叶癫痫,但动物实验仍有一定的局限性,不能完全模拟人类颞叶癫痫疾病。因此,该研究同时检测了TLR4蛋白在人脑组织中的表达,研究发现在癫痫患者脑组织切片中TLR4蛋白的表达也高于对照组。由于采取健康人脑组织样本不符合伦理学,故该研究以脑外伤后切除病灶周围的正常脑组织作为对照。
综上所述,该研究结果表明,TLR4蛋白的过度表达可能是癫痫发病的结果,而并非癫痫发生的始动点。对于癫痫急性发作后TLR4信号通路表达上调是否是导致癫痫反复发作的原因,有待于更深入的研究。
[1]Kim JE,Ryu HJ,Choi SY,et al.Tumor necrosis factor-alpha-mediated threonine 435phosphorylation of p65nuclear factor-kappaB subunit in endothelial cells induces vasogenic edema and neutrophil infiltration in the rat piriform cortex following status epilepticus[J].J Neuroinflammation,2012,9:6.
[2]Omran A,Peng J,Zhang C,et al.Interleukin-1beta and microRNA-146ain an immature rat model and children with mesial temporal lobe epilepsy[J].Epilepsia,2012,53(7):1215-1224.
[3]Levin J R,Serrano G,Dingledine R.Reduction in delayed mortality and subtle improvement in retrograde memory performance in pilocarpine-treated mice with conditional neuronal deletion of cyclooxygenase-2gene[J].Epilepsia,2012,53(8):1411-1420.
[4]茅东升,来婷婷,陈琰,等.Toll信号通路在中枢神经系统损伤中的作用[J].中国生物化学与分子生物学报,2010(2):121-127.
[5]Anderson KV.Toll signaling pathways in the innate immune response[J].Curr Opin Immunol,2000,12(1):13-19.
[6]Akira S,Takeda K,Kaisho T.Toll-like receptors:critical proteins linking innate and acquired immunity[J].Nat Immunol,2001,2(8):675-680.
[7]向秋莲,张慈柳,彭镜,等.TLR4、MRP8在内侧颞叶癫痫幼大鼠海马中表达的动态变化[J].中风与神经疾病杂志,2012,29(7):580-584.
[8]Sharma A.Immune response also connects autism and epilepsy[J].Nat Med,2011,17(8):922.
[9]Racine RJ.Modification of seizure activity by electrical stimulation.I.After-discharge threshold[J].Electroencephalogr Clin Neurophysiol,1972,32(3):269-279.
[10]杜春玲,邓学军,王芳,等.Semaphorin-3A与苔藓纤维出芽在毛果芸香碱致痫大鼠中的改变及意义[J].中国神经免疫学和神经病学杂志,2008,15(6):434-437.
[11]Yang Z,Liu X,Yin Y,et al.Involvement of 5-HT(7)receptors in the pathogenesis of temporal lobe epilepsy[J].Eur J Pharmacol,2012,685(1-3):52-58.
[12]Mathern GW,Pretorius JK,Babb TL.Quantified patterns of mossy fiber sprouting and neuron densities in hippocampal and lesional seizures[J].J Neurosurg,1995,82(2):211-219.
[13]肖波,江利敏.癫痫的发病机制[J].临床内科杂志,2004,21(9):577-580.
[14]Omran A,Peng J,Zhang C,et al.Interleukin-1beta and microRNA-146ain an immature rat model and children with mesial temporal lobe epilepsy[J].Epilepsia,2012,53(7):1215-1224.
[15]Levin J R,Serrano G,Dingledine R.Reduction in delayed mortality and subtle improvement in retrograde memory performance in pilocarpine-treated mice with conditional neuronal deletion of cyclooxygenase-2gene[J].Epilepsia,2012,53(8):1411-1420.
猜你喜欢
流行色(2022年7期)2022-08-31
临床骨科杂志(2021年1期)2021-12-03
保健与生活(2021年21期)2021-11-11
云南医药(2021年3期)2021-07-21
科学与财富(2020年15期)2020-07-04
移动通信(2020年4期)2020-05-07
家庭科学·新健康(2019年9期)2019-10-21
移动通信(2019年4期)2019-06-25
现代信息科技(2018年4期)2018-07-12
意林·少年版(2018年24期)2018-01-05
