
下肢打水在水中游进中的推力和控制机制
2014-05-14 02:39李树屏
天津体育学院学报 2014年2期
李树屏
下肢打水在水中游进中的推力和控制机制
李树屏
下肢击水的核心目标是增大游进的推力。从能量的角度分析,动力(推力)和阻力都来自于涡旋。高效率的推进依赖游进者的身体与流体流动环境相互作用的优化,即涡旋强度的平衡。高水平的打水模式是:下肢各关节依次运动向足部方向产生正弦波动,振动速度不发生衰减。人体系统无论对熟悉(如:脚蹼)还是不熟悉的任务型约束(如:腿蹼),在相应的运动模式未建立起来之前,都可以迅速地在训练前期加以调节。当任务型约束为非熟悉形式时,整合过程会延长。说明运动练习手段和辅助训练器材的时效性是一个值得关注的既有理论价值又有实践意义的课题。
下肢打水;推进力;动态系统理论;约束;动作协调
竞技游泳中对下肢的作用关注较少,人们的注意力常常放在身体姿势和上肢功能上[1],这是因为需要依靠上肢划水增大推进力和身体运动减小游进的阻力[2-4]。实际上,有研究早已证实,下肢的打水对游泳推进力的贡献不容忽视[2,5]。下肢的打水为全身摆动提供了一个稳定的平台,使游泳者得以减少阻力、优化推进力以及旋转的效率[6]。
下肢打水动作不仅在竞赛中有重要作用,而且无论对高水平游泳运动员还是初学者都是常用的训练和练习手段。对无上肢动作的下肢打水的研究主要集中在打水效率的运动学,如打水幅度的高低[7],打水时足的迎水面积大小[8]等。有研究利用电子量角器比较了自由泳、仰泳和蝶泳打水的下肢角度[9],发现,海豚式打水的膝关节最大弯曲角度和屈伸幅度都明显高于交叉式打水,而踝关节角度则无显著性差异。在25 m泳池中,0.5 m的水下比较从1.6~3.1 m/s间5种游速下的3种打水方式(自由式打水、海豚式打水和侧位海豚式打水,躯干和上肢成流线型姿势)的阻力,结果表明,在2.2 m/s游速,3种打水方式的阻力并无差别[10]。回归分析表明,自然幅度的海豚式打水的游速明显高于交叉式打水以及小幅和大幅度的海豚式打水,而踝关节和足的大幅度纵向运动加上较宽的足的面积有利于海豚式打水获得高的游速[11]。
研究下肢打水技术的目的主要在于如何提高打水的效果,其关键在于下肢动作与环境(水)之间相互作用的最佳化,这涉及到下肢打水推进的生物力学特征以及动作协调与优化的机制。
1 下肢打水推进的生物力学
1.1 推进模式
NEUHAUS等[12]提出,人体游泳时腿部的运动模式可分为2种:(1)海豚式,即2腿并拢击水;(2)交叉式,即2腿反相运动,如爬泳,类似于鱼类的尾部摆振。WEBB[13]根据鱼类的身体形状和运动方式将鱼类的推进模式分为3种类型,即全身加尾鳍周期性摆动型(BCF-p型)、全身加尾鳍快变(出发和转向)型(BCF-t型)和半身加腹鳍摆动型(MPF型)。相对于人体的运动,BCF和MPF型分别对应游泳时下肢和上肢的动作(见图1)。BCF模式是从身体的前部开始摆动一直到尾部,其中,从头顶部就开始摆动直至全身,称为全身波动,如鳗鲡科的游动,这种游进方式的加速性和机动性好,但能耗很高。半身波动从身体的下半部开始,速度、效率和机动性较高,如anguilliform(见图2)。尾部波动或围绕某支点的摆振,这是速度最快且效率最高的推进模式,为金枪鱼、海豚和鲨鱼等采用。
MPF模式是从身体摆动方式(如rajiform)到完全依靠鳍的摆动(如labriform)。labriform模式中还可分为2类,阻力型(摇橹型)推进和升力型(拍振型)推进,采用哪种形式,取决于游进的速度。阻力型(摇橹型)推进在低速游进时效率较高,而升力型(拍振型)推进在高速游进时效率较高,人体游泳时上肢的运动方式类似于labriform模式[14-15]。
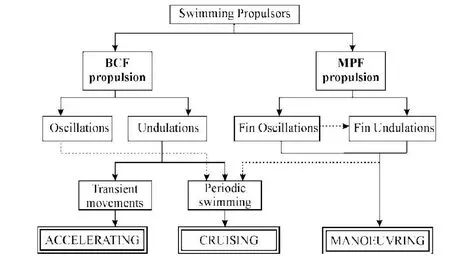
图1 鱼类推进器(鳍)类型与推进模式关系[13]
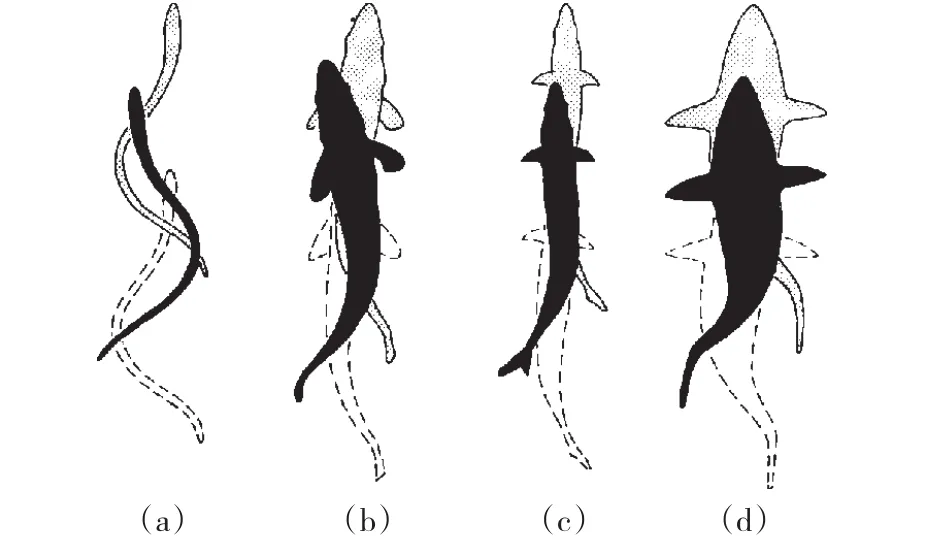
图2 BCF模式渐进图[16]注:(a)为 anguilliform,(b)为 subcarangiform,(c)为 carangiform,(d)为thunniform mode。
1.2 推力产生机制
对水生生物运动的数学和流体动力学研究始于LIGHTHILL[17]和WU[18],研究焦点主要围绕在水生生物的身体或鳍运动时是否产生逆卡门涡列作为推进力的涡旋的特性。尽管对水生生物2种运动模式(subcarangiform和carangiform型(下半身波动),thunniform和ostraciiform型(主要由尾部摆振))的数学处理上有所不同,但从涡旋的特性入手的思路主导了对水生生物推进力的研究。
LIGHTHILL的细长体理论是广泛应用于波动式游进分析的方法。根据细长体理论,身体的每一个波动运动产生向后的动量,依次在两侧产生有规律的涡旋传递到尾波,形成涡列。LIGHTHILL首先提出逆卡门涡列为推进力源的观点,并被应用到单个或成群的鱼类和水生哺乳动物的游进分析[19-21],以及人体的蹼泳运动分析中[22-25]。波动的推力可根据细长体理论由身体环节波动产生的侧向力计算(见图3)。
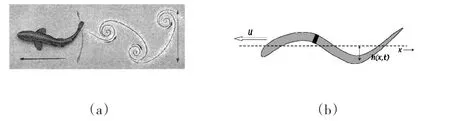
图3 反向卡门涡列图
对摆振式游进(如海豚、鲨鱼等)的研究还包括尾翼的翼型(高展弦比,如新月状尾翼)[27-28],从硬板式尾翼到三维可变形尾翼,以及不同形状下大幅度运动或小幅度运动的分析[21]。
近10多年来,基于对非定常流控制受到关注[29-31],研究证明,许多鱼类可以借助非定常流的能量加强推力和机动性。通过自身的动力学调节,鱼类可以做到高效地游进,可以产生大的推力和调节力,以及使用最小的能耗顺滑地游过流体[31]。
自从LIGHTHILL早期提出概念化的“细长体理论”以来,数学力学模型得到了明显的改进,以适应大幅度的波动分析[32-33],中心线以及尾翼与腹鳍产生的涡列的相互作用[34-36]。WU[18]提出了二维波面理论,将鱼类作为弹性板处理。身体的厚度对推力和阻力的影响也有研究[37-38]。KATZ和WEITHS[39]研究了身体弹性的影响。波动板理论的线性和非线性扩展分析也有研究[40-43]。
1.3 推力和效率的平衡
水生生物运动的研究结果可以为人体游泳所借鉴,特别是逆卡门涡列以及能量的再利用。一些研究者研究了卡门涡列在水下打水和脚蹼游进[22-24,44-47]。当尾鳍在最高点开始向下打水时,产生逆时针方向的涡旋,形成对后方的水的冲量,从而水对脚蹼(人体)产生向前的反作用力冲量;当脚蹼在最低点开始向上打水时,产生顺时针方向的涡旋,同样形成对后方的水的冲量,水仍然对脚蹼(人体)产生向前的反作用力冲量(见图3(a))。
尾涡的产生必须受到控制,一方面,需要产生足够多的涡旋以获得推进力;另一方面,人们需要避免过多的涡旋消耗过多的不能再利用的能量,在推力和能耗中取得平衡。这种平衡部分取决于涡旋与人体相应部位的相互作用。由此可见,打水在运动模式优化中的重要作用。
由此得出的关键概念是,游进者必须发现、产生适量的分布涡旋(卡门涡列),无论是波动还摆振型游进。平衡点能产生足够“有用的”涡旋去克服前进的阻力,同时避免产生多余的涡旋以致耗费不能再利用的能量。高效率的推进依赖游进者的身体和脚蹼与流体流动环境相互作用的优化。
近年来,有关尾翼谐波摆振产生推力原理的研究取得了显著的进展,连同下肢打水运动学的研究,以及数字模拟单蹼游进的研究结果表明,优化的脚蹼击水模式是身体各部依次波浪形运动传递到末端的脚蹼从而形成卡门涡列并加以有效的利用,产生推进力。
2 下肢打水动作协调与优化
2.1 打水训练与动态系统理论
下肢打水动作不仅在竞赛中有重要作用,而且无论对高水平游泳运动员还是初学者都是常用的训练和练习手段。动作练习、训练的目的是优化动作模式,动作练习应该在动作学习的初期尽早进行,因为不恰当的习惯性的动作模式一旦形成,就很难改变。
根据动态系统理论的观点,作为一个复杂系统,人体运动系统的行为由大量相互联系和相互作用的子系统通过系统的“自组织”(self-organization)功能来实现,这种由各类不同人体运动系统部分组合而成的功能单位被称为“协调结构”(coordinative structures)或“协同”(synergies)。这种临时的功能组合是有选择性的,有特定动作目标的,不受人体生物结构限制的系统组织状态,这种协调结构的可变性是运动动作可塑性的重要理论基础[48]。
动力系统理论被较多的应用于考察人体运动系统的动作模式,其主要特点有以下2点。(1)多环节系统协调分析。WICKELGREN[49]曾在《Science》杂志报道,小脑受损的病人对单关节运动的控制尚属正常,但对多关节运动的神经肌肉控制则存在动作协调的功能障碍。这从生物力学的角度说明了神经肌肉系统对单环节(关节)运动的控制与协调是有别于多环节(关节)的。鉴于人体运动是多环节参与的特性,多环节系统的分析有助于更深入地了解人体系统的动作控制和演变的规律。动力系统理论在系统运动学参数的基础上采用相位分析等非线性方法,最常用的是从计算环节(或关节)的相位角入手,分析环节相位角位移—角速度关系(相平面),和对任意2个环节连续相对的相位关系(即2个环节相位角位移—角速度参数的差)。连续相位关系的优势则在于同时体现2个环节的4个位移和速度关系来研究多环节系统的协调和控制功能。(2)长程观测和重复测量。动力系统理论的一个关键概念是“约束”,即人体内外环境对运动的限制。一个熟练的动作形式作为神经—肌肉协调和控制的优化模式,是由身体结构型约束(如身高、体重)、环境型约束(如温度、重力)和任务型约束(如运动的目的、运动时所用运动装备)的相互作用决定的[50]。如穿戴不同的蹼(穿在脚上的脚蹼和穿在腿上的腿蹼),人体环节系统的协调模式是不同的,即约束条件决定了动作的形式[51]。但多数研究往往只关注了人体运动系统对约束条件的最初的反应,这并不能保证所得到的系统对约束的反应和适应的真实性[52]。
2.2 打水动作模式的优化
有研究报道,通过比较不同水平游泳练习者打水动作模式的横向研究,分析游泳初学者在“学习游泳”(打水)过程中与高水平游泳运动员动作模式的不同之处[1]。研究发现,随着训练任务的完成,各水平游泳初学者在打水训练中下肢各关节的运动趋向于采取适当的运动幅度,从而产生简单的正弦运动。对髋、膝、踝关节竖直运动的傅里叶分析计算出身体波动的传递速度以及生物学噪声和变异度。髋-膝和膝-踝波动速度比指数表明,多数低水平的游泳练习者下肢关节的协调程度没有达到高打水效率的要求。而有明显的证据显示,高水平的打水动作特征是:各关节依次运动向足部方向产生正弦波动,振动速度不减小甚至增大,生物学噪声低和变异度小。
通过熟悉任务型约束的不同组合形式,运动员学习快速适应竞赛中约束条件的各种变化,这种适应约束条件变化的能力(速度)取决于2个因素。(1)运动员是否“熟悉”该任务约束。熟悉的任务约束定义为在训练中经常出现的约束,非熟悉的任务约束则是训练中从未出现过的约束形式。(2)任务约束变化的引入是否会使得动作模式需要优化。因此,当任务型约束发生变化时,会有4种情况出现:(1)形成新的动作模式以适应非熟悉的任务型约束;(2)形成新的动作模式以适应熟悉的任务型约束;(3)以不变的运动模式,通过肌肉的协同作用应对不同的任务约束条件以适应非熟悉的任务型约束[53];(4)以不变的运动模式,通过肌肉的协同作用应对不同的任务约束条件以适应熟悉的任务型约束。
2.3 打水动作模式形成和演变的机制
有关训练的次数(时间与频率)与适应任务型约束变化的研究少见报道。目前的研究有2个值得注意的问题:(1)多数研究没有涉及在不同约束条件之间,优化的动作模式在什么程度上是不同的;(2)多数研究往往只关注了人体运动系统对约束条件的最初反应,这并不能保证所得到的系统对约束的反应和适应的真实性[53]。
根据动态系统理论,肌肉系统会自组织协同以适应给定的约束条件下的运动,适应约束条件形成动作模式的速度取决于系统寻找“显现因子”所需的时间。因此,当需要形成新的动作模式但缺乏对特定约束条件的体验(经验)时,系统的适应(调节)过程就会较长。上文表明,高效的下肢打水动作是肢体各环节依次由近端向远端作正弦波动,由于系统“知道”最佳动作模式的特征,因而有助于加快对变化的约束的适应而无需寻找感觉-运动因子去优化建立新的动作模式[50,53]。在训练中,教练或老师的任务是利用各种任务约束手段来帮助练习者、运动员找到基本的和个性化的动作协调方案[54],以及模拟竞赛条件进行训练。可见,从任务约束结构和组织训练环境的角度看,教练的作用是不可或缺的[54-55]。
使用脚蹼来训练下肢打水动作,满足改变任务型约束而不引起改变基本动作模式的条件。游泳练习者经常使用脚蹼训练,因而脚蹼与腿部打水不同,但熟悉的任务型约束条件,一种新型的穿在小腿上的腿蹼则提供了不同,但为大多数游泳练习者不熟悉的任务型约束条件(见图4)。使用腿蹼可以减小膝关节的受力,同时在打水时使下肢三环节变成了二环节系统。这样,练习者不仅要适应腿部力矩的变化,而且要适应三环节系统压缩为二环节系统的变化,即第2个环节加长(小腿+足)。

图4 腿蹼(a)和脚蹼(b)
SANDERS等[56]研究了下肢打水在无蹼、脚蹼和腿蹼条件下,即任务型约束变化条件下的调节适应过程。9位达到竞赛年龄的少年游泳练习者(11~14岁)随机分为腿蹼组、脚蹼组和无蹼对照组,受试者均有经常使用脚蹼作打水练习的经验。有蹼组在训练前进行5次无蹼打水测试,完成60次带蹼训练(每周10次)后,再进行10次无蹼打水测试。控制组则只参加训练前的5次和训练后的10次无蹼打水测试。所有训练和测试均用测试游泳池装备的数字化水下影像记录系统记录,并采用APAS计算机影像解析系统数字化解析所获得的相关数据。受试者的运动模式与任务约束的契合程度采用对髋、膝、踝关节竖直运动的傅里叶分析,计算出髋-膝和膝-踝波动速度比指数加以判定。结果表明,无论是腿蹼组还是脚蹼组,重建适当的运动模式的调节过程都发生在第一次训练节段,即前10次带蹼训练中,但非熟悉任务型约束变化组(腿蹼组)在10次训练后的打水动作仍未达到优化的程度。
3 小 结
优化的脚蹼击水模式,是身体各部依次波浪形运动传递到末端的脚蹼从而形成卡门涡列并加以有效利用,产生推进力。关键是,一方面需要产生足够的“有用的”涡旋去克服前进的阻力;另一方面,避免产生多余的涡旋以致耗费不能再利用的能量。高效率的推进依赖游进者的身体和脚蹼与流体流动环境相互作用的优化。在此基础上,本文论述了在动态系统理论框架下打水动作模式的优化及动作模式形成和演变的机制。高水平的打水模式是下肢各关节依次运动向足部方向产生正弦波动,振动速度不发生衰减。人体系统无论对熟悉还是不熟悉的任务型约束,在相应的运动模式未建立起来之前,都可以迅速地在训练前期加以调节,但快速的调节并不妨碍肌肉持续微调整合打水动作,即当任务型约束为非熟悉形式时,整合过程会长于熟悉任务型约束。
[1]SANDERS R H.Kinematics,coordination,variability,and noise in the prone flutter kick at different stages of a learn to swim program[J].Journal of Sports Sciences,2007,25:213-227.
[2]ADAMS M.Thoughts on the crawl stroke[J].Swimming technique,2000,37(2):17-23.
[3]ARELLANO R,BROWN P,CAPPAERT J,et al.Analysis of 50-,100-,and 200-m freestyle swimmers at the 1992 Olympic Games[J].Journal of applied biomechanics,1994,10(2):189-199.
[4]KENNEDY P K,BROWN P L,CHENGALUR S N,et al.Analysis of male and female Olympic swimmers in the 100-meter events[J].International journal of sport biomechanics,1990,6:187-197.
[5]HOLLANDER A P,DE GROOT G,VAN INGEN SCHENAU G J,et al.Contribution of the legs to propulsion in front crawl swimming[M].Champaign,IL:Human Kinetics,1988.
[6]MAGLISCHO E W.Swimming Even Faster[M].CA:Mayfield,1993.
[7]ALLEY L E.An analysis of water resistance and propulsion in swimming the crawl stroke[J].Research quarterly,1952,23:257-270.
[8]FUJIWARA H,OGITA F.Effects of the foot frontal area and the lower limbs flexibility on the maximal swimming velocity during leg kick and whole body stroke of front crawl[D].Japan:University of Tokyo,1997.
[9]SHEERAN T J.Electrogoniometric analysis of the knee and ankle of competitive swimmers[J].Journal of human movement Studie,1980,6(3):227-235.
[10]LYTTLE A D,BLANKSBY B A,ELLIOTT B C,et al.Net forces during tethered simulation of underwater streamlined gliding and kicking techniques of the freestyle turn[J].Journal of sport sciences,2000,18:801-807.
[11]CLOTHIER P.Underwater kicking following the freestyle tumble turn[EBOL].http://www.digthesis.ballarat.edu.au/adt/public/adt-ADT2005 1026.161431/index.html.
[12]NEUHAUS P D,O’SULLIVAN M,EATON D,et al.Concept designs for underwater swimming exoskeletons.Proceedings of the 2004 IEEE international conference on robotics and automation,New Orleans,LA April 2004[EBOL].http://www.ieeexplore.ieee.org/iel5/9126/28923/01302493.pdf.
[13]WEBB P W.Body form,locomotion and foraging in aquatic vertebrates[J].American zoologist,1984,24:107-120.
[14]SCHLEIHAUF R E.A hydrodynamic analysis of swimming propulsion[M].Maryland:University Park Press,1979.
[15]TOUSSAINT H,TRUIJENS M.Biomechanical aspects of peak performance in human swimming[J].Animal biology,2005,55(1):17-40.
[16]LINDSEY C C.Form,function and locomotory habits in fish[M].London:Academic Press,1978.
[17]LIGHTHILL M J.Note on the swimming of slender f i sh[J].Journal of fluid mechanic,1960,9:305-317.
[18]WU T Y.Swimming of a waving plate[J].Journal of fluid mechanics,1961,10:321-344.
[19]WEIHS D,WEBB P W.Optimization of locomotion[M].New York:Praeger,1983.
[20]MÜLLER U,VAN DEN HEUVEL B,STAMHUIS E,et al.Fish foot prints:morphology and energetics of the wake behind a continuously swimming mullet[J].Journal of experimental biology,1997,200:2893-2896.
[21]SFAKIOTAKIS M,LANE D M,DAVIES B C.Review of fish swimming models for aquatic locomotion[J].IEEE journal of oceanic engineering,1999,24(2):237-252.
[22]李树屏.人体与运动器械在流体中的阻力和动力[J].浙江体育科学,1998,20(6):34-38.
[23]LI S,HONG Y,LUK T C.The power initiating point and tail vortex in scuba swimming[D].Hong Kong:The Chinese University of Hong Kong,2000.
[24]李树屏.运动生物力学[M].北京:高等教育出版社,2000.
[25]ARELLANO R,PARDILLO S,GAVILAN A.Underwater undulatory swimming:kinematic characteristic,votex generation and application during the start,turn and swimming strokes[D].Spain Universidad de Extremadura,Caceras,2002.
[26]TRIANTAFYLLOU M S,TRIANTAFYLLOU G S.An efficient swimming machine[J].Scientific American,995,272:64-70.
[27]LIGHTHILL M J.Aquatic animal propulsion of high hydromechanical efficiency[J].Journal of fluid mechanics,1970,44:265-301.
[28]WU T Y.Hydrodynamics of swimming propulsion.Part 2.Some optimum shape problem[J].Journal of fluid mechanics,1971b,46:521-544.
[29]TRIANTAFYLLOU M S,TRIANTAFYLLOU G S,YUE D K P.Hydrodynamics of fishlike swimming[J].Annual review of fluid mechanics,2000,32:33-53.
[30]COLGATE J E,LYNCH K M.Mechanics and control of swimming:a review[J].IEEE journal of oceanic engineering,2004,29(3):660-673.
[31]BEAL D N,HOVER F S,TRIANTAFYLLOU M S,et al.Passive propulsion in vortex wakes[J].Journal of fluid mechanics,2006,549:385-402.
[32]LIGHTHILL M J.Large-amplitude elongated-body theory of fish locomotion[J].Proc.R.Soc.Lond.B,1971,179:125-138.
[33]WARDLE C S,REID A.The application of large amplitude elongated body theory to measure swimming power in fish[M].New York:Academic,1977.
[34]WU T Y.Hydrodynamics of swimming propulsion.Part 1.Swimming of two dimensional flexible plate at variable forward speed in an inviscid fluid[J].Journal of fluid mechanics,1971a,46:337-355.
[35]WU T Y.Hydrodynamics of swimming propulsion.Part 2.Some optimum shape problem[J].Journal of fluid mechanics,1971b,46:521-544.
[36]WU T Y.Hydrodynamics of swimming propulsion.Part 3.Swimming and optimum movement of slender fish with side fins[J].Journal of fluid mechanics,1971c,46:545-568.
[37]NEWMAN J N.The force on a slender fish-like body[J].Journal of fluid mechanics,1973,58(4):689-702.
[38]NEWMAN J N,WU T.A generalized slender-body theory for fish-like forms[J].Journal of fluid mechanics,1973,57:673-93.
[39]KATZ J,WEIHS D.Hydrodynamic propulsion by large amplitude oscillations on an airfoil with chordwise flexibility[J].Journal of fluid mechanics,1978,88:485-497.
[40]CHENG J Y,ZHUANG L X,TONG B G.Analysis of swimming three dimensional plates[J].Journal of fluid mechanics,1991,232:341-355.
[41]TONG B G,ZHUANG L X,CHENG J Y.The hydrodynamic analysis of fi sh propulsion performance and its morphological adaptation[J].Sadhana-Academy proceedings in engineering and sciences,1993,18:719-728.
[42]CHENG J Y,BLICKHAN R.Note on the calculation of propeller eff i ciency using elongated body theory[J].Journal of experimental biology,1994,192:169-177.
[43]ZHU Q,WOLFGANG M J,YUE D K P,et al.Three dimensional flow structures and vorticity control in fish-like swimming[J].Journal of fluid mechanics,2002,488:1-28.
[44]LI S.Biomechanical study of fin-swimming and fins[M].Vic:Dept.of Physical Education and Recreation,1989.
[45]UNGERECHTS B E,DALY D,ZHU J P.What dolphins tell us about hydrodynamics[J].Journal of swimming research,1998,13:1-7.
[46]UNGERECHTS B E,PERSYN U,COLMAN V.Application of vortex flow formation to self-propulsion in water[M].Jyvaskla:Gummerus Printing House,1999.
[47]UNGERECHTS B,PERSYN U,COLMAN V.Analysis of swimming techniques using vortex traces[D].Hong Kong:The Chinese University of Hong Kong,2000.
[48]DAVIDS K,GLAZIER P,ARAUJO D,et al.Movement systems as dynamic systems:the functional role of variability and its implications for sports medicine[J].Sports medicine,200,33(4):245-260.
[49]WICKELGREN I.The cerebellum:The brain’s engine of agility[J].Science,1998,281:1588-1590.
[50]NEWELL K M.Constraints on the development of coordination[M].MA:Matinus Nijhof,1986.
[51]CLARK J E.On becoming skillful:patterns and constraints[J].Research quarterly for exercise and sport,1995,66(3):173-183.
[52]NOURRIT D,DELIGNIÈRES D,CAILLOU N,et al.On discontinueities in motor learning:a longitudinal study of complex skill acquisition on a ski-simulator[J].Journal of motor behaviour,2003,35(2):151-170.
[53]DAVIDS K,BENNETT S,HANDFORD C,et al.Acquiring coordination in self-paced,extrinsic timing tasks:A constraints led perspective[J].International Journal of Sport Psychology,1999,30:437-461.
[54]ARAU J O D,DAVIDS K,BENNETT S,et al.Emergence of sport skills under constraints[M].London:Routledge,2004.
[55]SCHMIDT R A,LEE T D.Motor control and learning[M].Champaign,IL:Human Kinetics,1999.
[56]ROSS S,SHUPING L,JOSEPH H.Adjustment to change in familiar and unfamiliar taskconstraints[J].Journal of Sports Sciences,2009,27(6):651-659.
Thrustandcontrolmechanismoflowerextremityunderwaterkickinginswimming
Li Shuping
(Center of Motor Control and Evolution,Hubei University,Wuhan 430062,China)
The core target of lower limbs kicking is to increase swimming thrust.Based on the view of energy analysis,both power(thrust)and resistance are come from the vortex.High efficiency promoting relies on the optimization of interaction between swimmer and fluid flow:the balance of vortex intensity.A high level kicking model is:all joints of the lower extremity generate sine wave to the direction of foot,vibration velocity does not decay.Either to the familiar or to the unfamiliar task constraints,the human system could adjusted quickly in the early stage of training before the corresponding motion mode was estab⁃lished.When a task constraint is unfamiliar,the integration process would be longer.It indicated that there are theoretical and practical values for the study on the time effectiveness of exercise methods and auxiliary training equipments.
lower limb underwater kicking;Thrust;dynamic system theory;constraint;coordination
G 804.6
A
1005-0000(2014)02-100-05
2013-11-15;
2014-02-08;录用日期:2014-02-09
李树屏(1957-),男,湖北武汉人,教授,博士,研究方向为生物力学。
湖北大学动作控制与发展研究中心,湖北武汉430062。
猜你喜欢
装备制造技术(2022年6期)2022-10-02
北京航空航天大学学报(2022年7期)2022-08-06
中国海洋大学学报(自然科学版)(2022年6期)2022-06-03
小天使·三年级语数英综合(2020年10期)2020-12-23
悦游 Condé Nast Traveler(2019年9期)2019-11-25
少年文艺·我爱写作文(2017年5期)2017-06-10
小学阅读指南·低年级版(2017年1期)2017-03-13
中国体育教练员(2015年3期)2015-11-03
人生十六七(2015年6期)2015-02-28
计算机辅助工程(2012年5期)2012-11-21
