
超级杂交晚稻不同生育阶段氮素亏缺与补偿效应
2015-05-15 05:59陈小荣黄文贇贺浩华
江西农业大学学报 2015年1期
陈小荣,黄 磊,钟 蕾,黄文贇,贺浩华
(江西农业大学 作物生理生态与遗传育种教育部重点实验室/江西省作物生理生态与遗传育种重点实验室/农学院,江西 南昌 330045)
氮素是影响作物生长发育及产量形成最重要、最活跃的可控养分因子之一,在作物生长过程中对氮素的动态管理一直是生产中的重要措施[1-3]。水稻是我国的主要粮食作物,氮肥的使用对稻谷生产曾发挥过巨大作用,但是当前氮肥利用率降低及增氮增产效果不再明显的问题日益突出。FAO资料表明,中国2002年以占世界1/5的水稻种植面积用去氮肥1/3以上[4],我国单季稻区施氮量(纯氮)平均为180 kg/hm2,比世界平均用量高75%左右[5],双季稻区农户施氮量一般也达到150~180 kg/hm2,高者超过225 kg/hm2[6],统计表明2005年我国氮肥当季利用率仅为30%~35%,损失高达40%~50%[7],氮肥过度施用已在中国引发严重环境问题[8]。众所周知,解决农作物氮肥利用率及施用效率问题必须确定氮肥施用的高效期和迟钝期,前提是查明作物氮素需求关键期及亏缺敏感期[9]。不难发现,以往对水稻氮素需求关键期及亏缺敏感期的研究通常采用相等总氮量在不同生育阶段的分配方案,即氮素运筹试验,也有采用氮肥分期叠增的方法,其共同缺点是未控制土壤本身肥力,且将氮肥大部分作基肥,剩余小部分在不同生育阶段进行分配,无法排除土壤固有速效氮和基础氮对不同生育阶段追氮效果的干扰,而研究表明土壤基础地力或土壤背景氮(indigenous nitrogen supply)对水稻产量和氮肥利用率的影响不容忽视[10]。鲜见有对不同生育阶段氮素亏缺及前一阶段亏缺后一阶段常量、加倍供应补偿效应的研究报道。也就是说,十分有必要引入作物亏缺补偿效应理论与方法来研究水稻氮素利用的科学问题。补偿生长是自然界普遍存在的一种生物现象,学者们经过大量的研究,认为补偿或超补偿效应是指作物受阈值内的胁迫压力后,在结构和生理水平上产生的一种有利于作物生长发育和产量形成的能力[11]。近年来人们发现玉米、大豆、早熟禾、小麦等作物在氮素营养上均存在明显的亏缺补偿效应[12-16],尤其是对小麦氮素亏缺补偿效应进行了较为深入的研究[9,15-16]。
长江流域双季稻区是我国稻谷主产区之一,近年来推广了一批超级杂交稻品种,科技人员对其需氮规律进行了一定的探讨[17-19],提出的前氮适当后移,适度降低基追肥比例,重视穗粒肥等施氮理论与技术得到普遍认可。但是,总体看来,由于超级稻品种有其自身生长发育及栽培生理特点[20],迄今对超级稻高产高效的调控途径尚不十分清楚,因而目前超级稻水肥管理仍大多沿用常规高产水稻品种的灌溉和施肥技术,这也是限制超级稻产量潜力发挥的一个重要原因[21]。据此,本研究以超级杂交晚稻品种为材料,选用速效氮含量较低的土壤,将氮肥平均分配到各个主要生育阶段,研究不同生育阶段氮素亏缺,前一生育阶段亏缺后一生育阶段等量、加倍恢复供应对其生长发育的影响,以期查明超级杂交晚稻氮素亏缺敏感期及补偿有效期,为提高氮肥利用率及增加产量提供依据,也为水稻氮素利用研究提供一个生态学新视角。
1 材料与方法
1.1 水稻材料
本试验于2012年在江西农业大学科技园进行。五丰优T025,2010年通过国家审定并被农业部认定为超级稻,多年多点观察发现产量性能优异,大田一般产量为 8 000 kg/hm2,高产田块达1 100 kg/hm2,是近年来江西等双季稻区主栽晚稻品种之一。
1.2 试验设计
浸种催芽后于6月30日播种于大田,4叶1心期挑选整齐一致幼苗人工移栽入桶中,即7月15日移栽,桶高为24 cm、上部内径30 cm、底部直径23.5 cm塑料桶,每桶栽插3穴,每穴1粒谷苗,10月27日收割。选取全氮及速效氮含量低,近几年来一直撂荒红壤稻田0~20 cm耕层土为供试土壤,土壤经自然晒干,用土壤粉碎机粉碎过网筛,移栽前预先淹水1周。土壤理化性质:pH5.73,有机质37.72 g/kg,全氮4.60 g/kg,碱解氮 50.89 mg/kg,有效磷 9.81 mg/kg,速效钾 76.73 mg/kg。幼苗期肥、分蘖期肥、幼穗分化期肥、抽穗期肥、乳熟期肥分别于7月8日(移栽前7 d)、7月30日(移栽后15 d)、8月29日(移栽后45 d)、9月16日(移栽后61 d)、10月1日(移栽后75 d)按试验设计量施用。氮素供应试验方案具体如下:参考翟丙年等[15]对冬小麦氮素亏缺补偿效应的研究方法,结合水稻总需氮特点设计各处理,共12个,每个处理重复10次,总共120桶,按随机区组排列。移栽前每桶施钙镁磷矿粉6.0 g,分蘖期和抽穗期每桶分别施氯化钾 1.5 g 和 1.5 g;氮肥(N)用尿素,其中 4.0 g/桶约合纯 N 160 kg/hm2,5.0 g/桶约合纯N 200 kg/hm2,根据试验方案幼苗期肥在装桶时混入,其余均在各生育阶段开始时结合灌水施入,施肥后2 d内密切关注天气变化,遇雨则将其移入防雨棚,雨后及时移回。
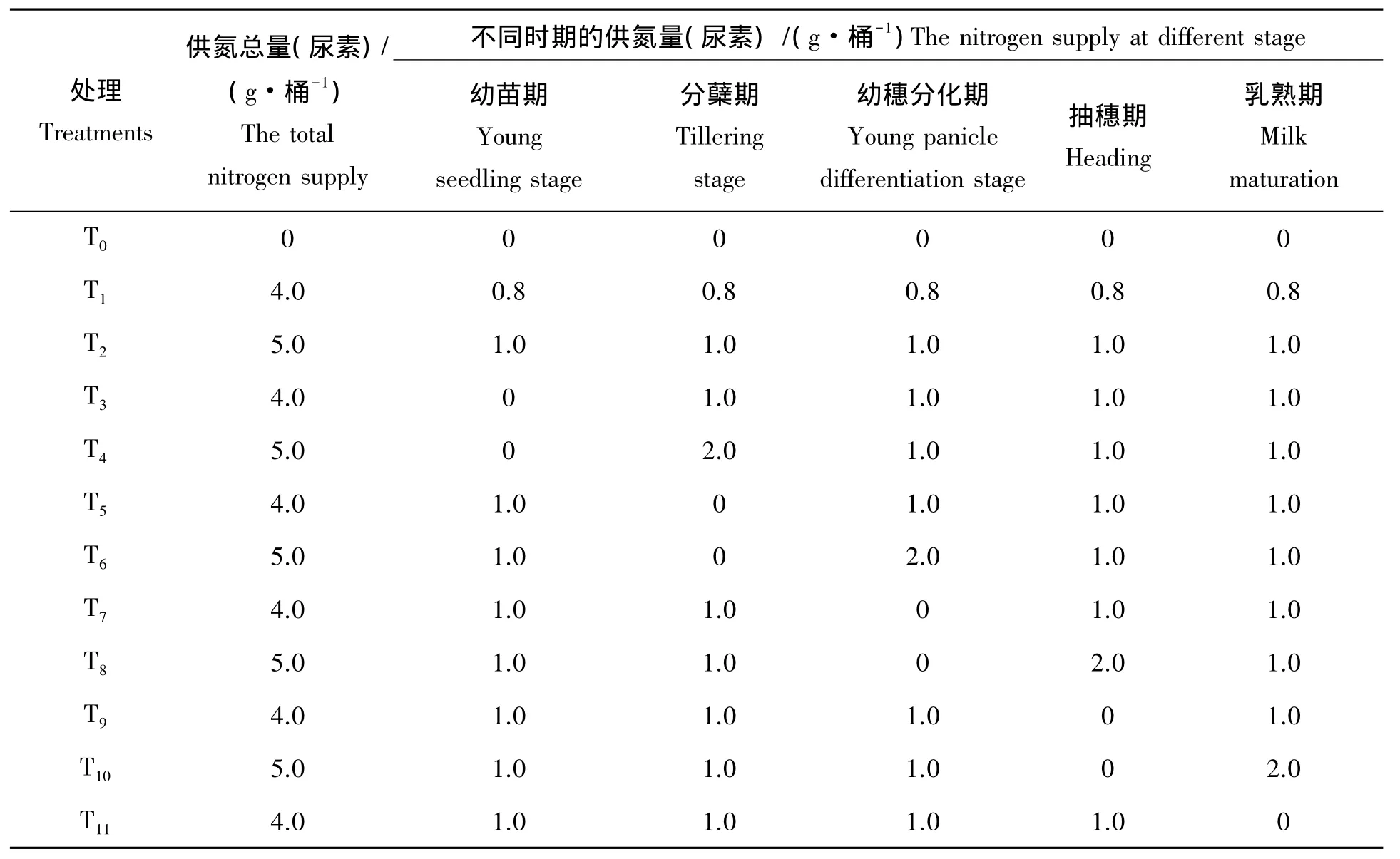
表1 氮素供应试验方案Tab.1 Experiment design for N supply
1.3 取样方法与测定项目
1.3.1 分蘖 移栽后各处理定5株观察分蘖动态,每隔5 d计数1次,本文选取分蘖盛期、穗分化中期和抽穗期单株茎蘖数进行分析。
1.3.2 干生物质量 于成熟期按平均茎蘖数法取3桶即9株,分桶收获,每桶中3株计为1次重复,剪除根,将稻株分为叶片、茎鞘和穗3部分洗净包装,于烘箱105℃杀青20 min,然后保持80℃烘干至恒质量,最后称量,计为干生物质量。
1.3.3 氮素含量 将上述干物质测定用样株按单株分成茎鞘、叶及籽粒3部分,然后粉碎,采用全自动凯氏定氮法测定全N。
1.3.4 产量及其构成 成熟后每个处理取5桶即15株,分桶收获,每桶中3株计为1次重复,晒干计产,并进行产量构成因素指标调查。
1.4 数据处理及计算方法
氮肥农学利用率(ANUE,g/g)=(施氮区产量-空白区产量)/施氮量 (1)
氮收获指数(NHI%)=籽粒中氮素累积量/植株氮素累积量×100 (2)
氮肥生理利用率(NPE%)=(施氮区产量-空白区产量)/(施氮区植株氮素累积量-空白区植株氮素累积量)×100 (3)
氮肥偏生产力(PFP)=施氮处理产量/施氮量 (4)
氮肥吸收效率(NUPE)=植株地上部氮素累积量/施氮量 (5)
数据处理采用Excel 2007,统计分析使用DPS软件完成。
2 结果与分析
2.1 分蘖
由表2可知,超级杂交晚稻所有11个施氮处理分蘖盛期、穗分化中期及抽穗期单株茎蘖数均显著高于对照T0不施氮,而T2高于T1表明随着施氮量增加单株分蘖数将上升。至抽穗期,幼苗期和分蘖期氮素亏缺处理T3、T4、T5、T6显著低于幼穗分蘖期、抽穗期及乳熟期氮素亏缺处理,而处理T8显著高于其余各处理。幼苗期氮素亏缺,分蘖期氮素加倍供应处理T4较等量恢复供氮处理T3,分蘖期氮素亏缺,穗分化期氮素加倍恢复供氮处理T6较等量处理T5幼穗分化期亏缺,抽穗期氮素加倍恢复供氮处理T8较等量处理T7在抽穗期的单株茎蘖数均更高,显示出氮素对茎蘖生长的亏缺补偿效应。从表2进一步可看出,就氮素亏缺时期来看,幼苗期、分蘖期氮素亏缺对超级杂交晚稻分蘖的影响最大,抽穗后氮素亏缺的影响程度降低,且补偿效果也更差。综上,幼苗期至分蘖期为超级杂交晚稻分蘖生长需氮关键期和亏缺敏感期,其补偿的有效临界期为抽穗期。
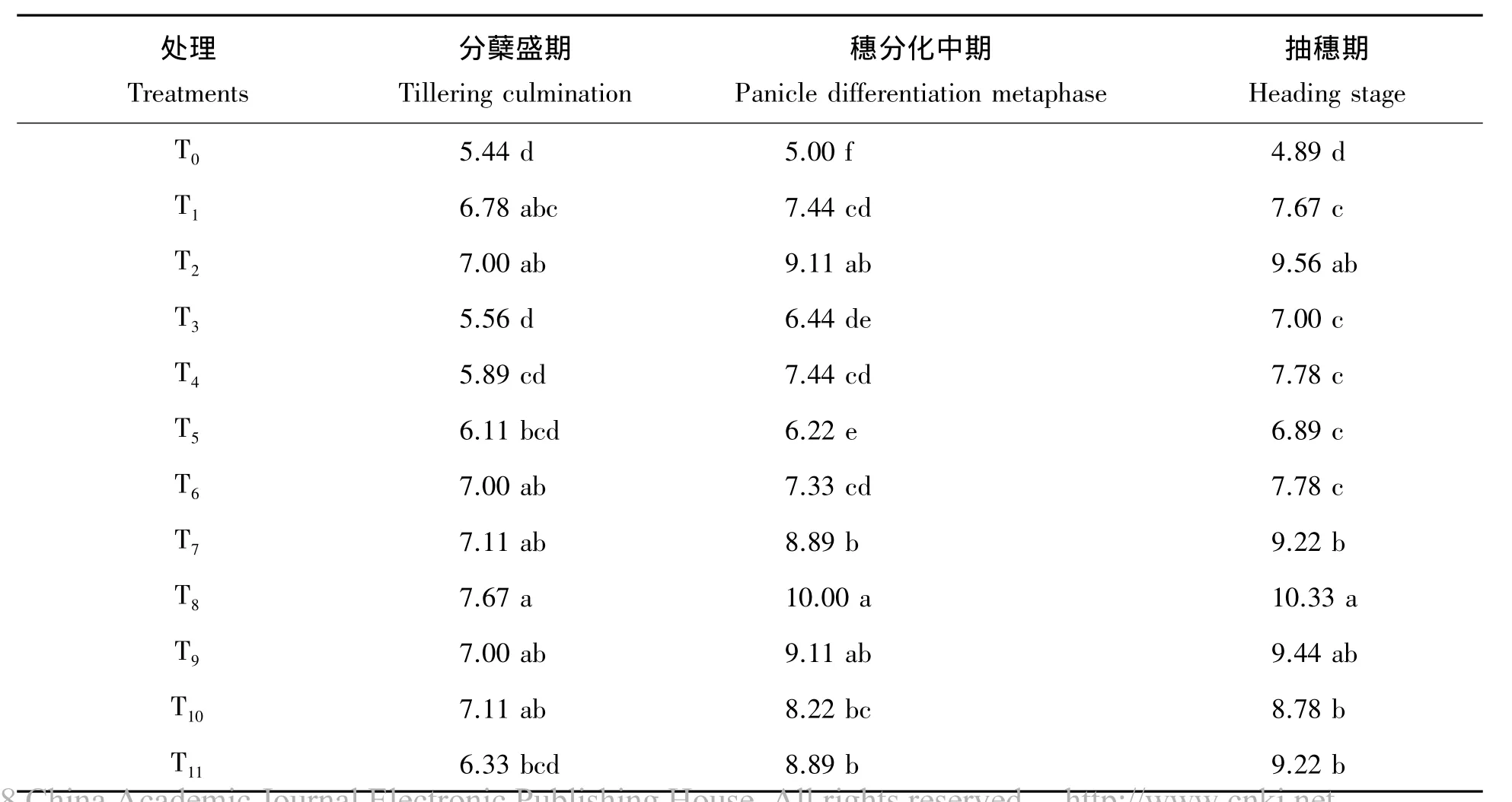
表2 不同氮素亏缺处理下超级杂交晚稻单株茎蘖数Tab.2 Tillers per plant under the different nitrogen deficiency treatments for the super hybrid late rice
2.2 物质生产
由表3可看出,施氮促进了超级杂交晚稻茎、叶和穗干物质积累,所有施氮处理的茎、叶和穗部器官干生物质量均高于不施氮处理T0,每个生育阶段等量施氮量下施氮多的处理均高于少的处理,尤其是穗部干生物质量随着施氮量的增加,这种促进作用越明显。另外,所有前一生育阶段缺氮,后一生育阶段加倍恢复供氮处理下的地上部总干生物质量和穗部干生物质量均高于等量处理,其中幼苗期缺氮分蘖期加倍、分蘖期缺氮幼穗分化期加倍、幼穗分化期缺氮抽穗期加倍、抽穗期缺氮乳熟期加倍处理较等量处理下的穗部干生物质量分别增加35.86%、3.47%、3.80%和11.94%,地上部总干生物质量分别增加33.19%、18.70%、3.34%和-0.25%,显示出氮素对超级杂交晚稻干生物质量具有较为明显的亏缺补偿效应。抽穗后氮素亏缺各处理对地上部总干生物质量基本无影响,也就是说,分蘖期至幼穗分化期氮素亏缺对超级杂交晚稻物质生产及其在穗部的积累影响大,为其干生物质量生产与积累的需氮关键期和亏缺敏感期,其补偿的有效临界期为抽穗期。
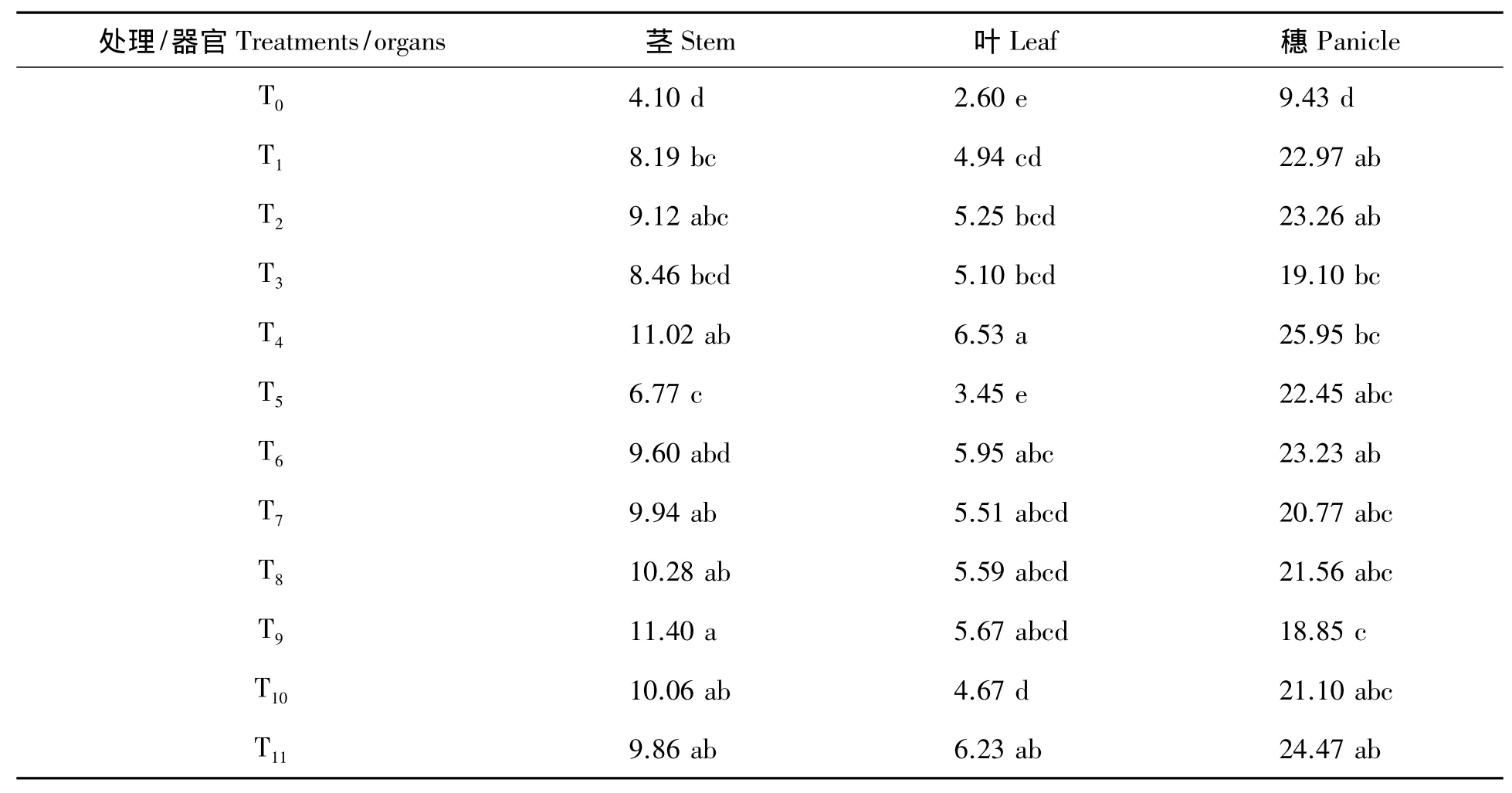
表3 不同氮素亏缺处理下超级杂交晚稻成熟期单株茎、叶、穗器官干生物质量Tab.3 Dry matter weight of the stem,leaf,panicle organ per plant at ripening under the different nitrogen deficiency treatments for the super hybrid late rice g
2.3 产量及产量构成
由表4可知,单株产量均以全生育期不施氮处理T0最低,以乳熟期氮素亏缺处理T11最高,每个生育阶段等量施氮量下施氮多的处理单株产量均高于少的处理。所有前一生育阶段缺氮,后一生育阶段氮素恢复加倍处理均高于等量处理,其中幼苗期亏缺分蘖期加倍、分蘖期亏缺幼穗分化期加倍、幼穗分化期亏缺抽穗期加倍、抽穗期亏缺乳熟期加倍处理较等量处理下分别增加4.42%、10.81%、0.98%和3.89%,显示出氮素对超级杂交晚稻产量具有明显的亏缺补偿效应,尤以分蘖期缺氮幼穗分化期补偿效应最强。就氮素亏缺时期来看,处理T5、T6、T7、T8单株产量均较低,为分蘖期至幼穗分化期氮素亏缺处理,而T3、T4、T9、T10、T11单株产量明显较高。也就是说,分蘖期前和抽穗后氮素亏缺对超级杂交晚稻单株产量影响较小,表明分蘖期至幼穗分化期为其需氮关键期和氮素亏缺敏感期,其补偿的有效临界期为抽穗期,与干生物质量结果基本一致。
表4表明,单株有效穗数均以全生育期不施氮处理T0最低,以幼苗期氮素亏缺处理,分蘖期加倍恢复供氮处理T4最高。就氮素亏缺时期来看,分蘖期及幼穗分化期氮素亏缺对超级杂交晚稻单株有效穗数影响最大,该时期氮素亏缺较幼苗期、抽穗期及乳熟期氮素亏缺处理下降的幅度更大。由此可见,超级杂交晚稻单株有效穗数氮素亏缺敏感期及需氮关键期为分蘖期至幼穗分化期。幼苗期亏缺,分蘖期加倍恢复供氮处理下的单株有效穗数明显高于等量处理,表明超级杂交晚稻在该时期内表现出了较强的氮素亏缺补偿效应,分蘖亏缺,幼穗分化期加倍恢复供氮处理下的单株有效穗数略高于等量处理;而幼穗分化期后等量和加倍恢复供氮间单株有效穗数基本持平,说明超级杂交晚稻单株有效穗数氮素补偿的有效临界期为幼穗分化期。
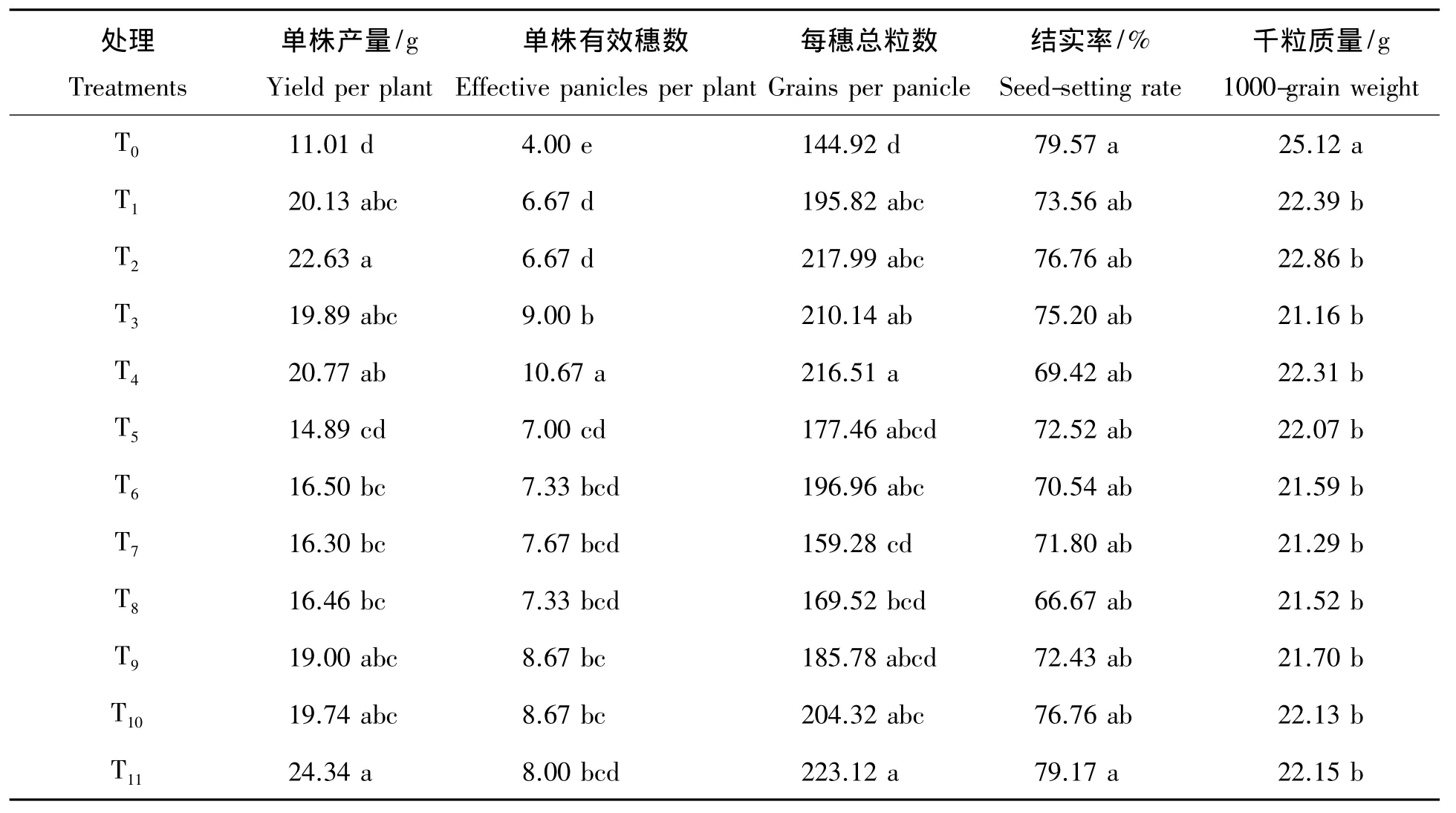
表4 不同氮素亏缺处理下超级杂交晚稻产量及其构成因素Tab.4 Yield and its components under the different nitrogen deficiency treatments for the super hybrid late rice
表4可看出,每穗总粒数均以全生育期不施氮处理T0最低,以乳熟期氮素亏缺处理T11最高,每个生育阶段等量施氮下,施氮多的处理均高于少的处理。所有前一生育阶段缺氮,后一生育阶段氮素恢复加倍处理均高于等量处理,其中幼苗期亏缺分蘖期加倍、分蘖期亏缺幼穗分化期加倍、幼穗分化期亏缺抽穗期加倍、抽穗期亏缺乳熟期加倍处理较等量处理下分别增加3.03%、10.99%、6.43%和9.98%,显示出氮素对超级杂交晚稻每穗总粒数亏缺补偿效应。就氮素亏缺时期来看,处理T7、T8每穗总粒数均较低,表明幼穗分化期氮素亏缺对超级杂交晚稻每穗总粒数影响大,为其需氮关键期和氮素亏缺敏感期,其补偿的有效临界期为抽穗期。
表4表明,超级杂交晚稻结实率以全生育期不施氮处理T0最高,幼穗分化期氮素亏缺、抽穗期加倍恢复供氮处理T8最低,但所有处理间差异均不显著。超级杂交晚稻全生育期中任一生育阶段氮素亏缺,下一生育阶段加倍恢复供氮处理下的结实率基本低于等量处理,未发现该指标具有氮素亏缺补偿效应。
表4还可看出,超级杂交晚稻以全生育期不施氮肥T0千粒质量最高,各氮素亏缺处理间差异不显著。幼穗分化期氮素亏缺处理T7、T8与其他各时期氮素亏缺处理相比较千粒质量较低,即在此时期氮素亏缺对超级杂交晚稻的千粒质量影响最大,为其氮素亏缺敏感期、需氮关键期。此外,各期氮素亏缺后加倍恢复供氮处理千粒质量与等量处理间差异无一致规律,表明超级杂交晚稻千粒质量指标无明显氮素亏缺补偿效应。
2.4 氮肥利用
表5可知,相比全生育期不施氮处理T0,处理T11ANUE增加幅度最高,穗分化期氮素亏缺处理T7、T8最低,表明穗分化期缺氮对超级杂交晚稻氮肥ANUE影响最为明显;抽穗期缺氮,乳熟期恢复供氮后,加倍供氮处理较等量供氮处理的ANUE高,表明抽穗期补追氮肥对超级杂交晚稻穗分化期缺氮下的ANUE具有一定的补偿效果。NUPE上,以幼苗期氮素亏缺处理T3、T4最高,抽穗期氮素亏缺处理T9、T10最低,说明幼苗期氮素亏缺对超级杂交晚稻NUPE的影响最小,抽穗期氮素亏缺的影响最大;抽穗期亏缺乳熟期恢复供氮后,加倍供氮处理较等量供氮处理的NUPE更高,表明乳熟期补追氮肥对超级杂交晚稻抽穗期缺氮下的NUPE具有一定的补偿效果。NHI上,以处理T7、T8最低,说明幼穗分化期氮素亏缺对超级杂交晚稻NHI的影响最大。NPE上,以处理T5、T6最低,即分蘖期氮素亏缺处理,表明分蘖期氮素亏缺对超级杂交晚稻的NPE影响最为明显。表5还可看出,PFP上,就亏缺时期来看,分蘖期氮素亏缺处理T8和T6小,即分蘖期至幼穗分化期氮素亏缺对PFP影响最为明显。
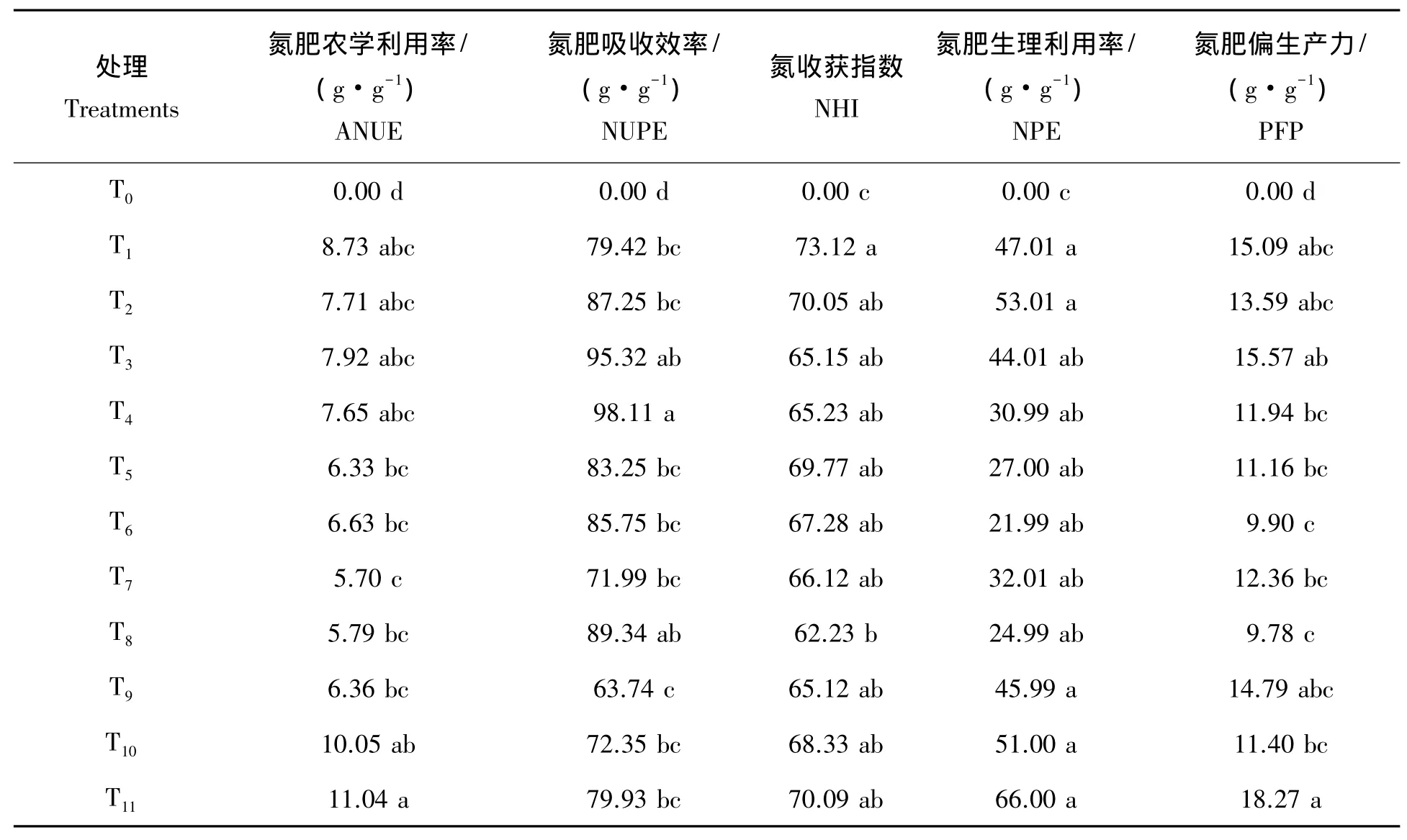
表5 不同氮素亏缺处理下超级杂交晚稻氮素吸收和利用效率Tab.5 Nitrogen uptake and utilization efficiency under the different nitrogen deficiency treatments for the super hybrid late rice
3 讨论
翟丙年等[15]研究表明,越冬期不施氮能明显减少冬小麦的分蘖数和地上、地下部生物量,越冬期和拔节期不施氮能显著降低冬小麦的产量,而后期缺氮对产量的影响不大。翟丙年等[16]进一步研究指出,越冬期和拔节期氮素亏缺不仅可以明显减小根长、根条数和根体积,而且可以减少根吸收总面积、活跃吸收面积、活跃吸收面积百分比、根比表面及根系活力,以越冬期更为显著。冬小麦对氮素的亏缺敏感期在越冬期和拔节期,补偿有效期在拔节期[15-16]。同为禾本科的水稻,本研究表明,从各生育阶段氮素亏缺处理对产量的影响上看,分蘖期、幼穗分化期氮素亏缺处理下产量最低,而幼苗期及抽穗期后氮素亏缺对其产量影响小,其中以乳熟期氮素亏缺在所有12个处理中产量最高。说明,分蘖期、幼穗分化期是超级杂交晚稻单株产量需氮关键期、氮素亏缺敏感期,生产上氮素运筹时应特别注意在该时期施足氮肥。分蘖期、幼穗分化期氮素亏缺后,其下一生育阶段加倍供氮处理较等量供氮处理下产量更高,表现出了较为明显的氮素亏缺补偿效应,而抽穗后缺氮未发现存在补偿效应,说明抽穗期是超级杂交晚稻氮素亏缺补偿的有效临界期,生产上对生育前期缺施进行补追时,应不晚于抽穗期。冬小麦和超级杂交晚稻两研究结果说明,依靠分蘖成穗的禾本科作物,其氮素亏缺的敏感期和需求的关键期为分蘖期至幼穗分化期,氮素亏缺的补偿有效临界期为抽穗期,我们对超级杂交早稻的研究结果也支持了该结论(另文报道)。但对于生育期长的品种(中稻或一季稻)来说,氮素亏缺的敏感期、需求的关键期及补偿有效期是否应延后,值得进一步研究。
水稻产量由单株有效穗数、每穗总粒数、千粒质量和结实率4个因素构成,各因素在生育进程中先后形成,其间存在着相互制约、相互补偿的关系。有研究认为保持结实率和单株有效穗数稳定在较高水平对品种产量稳定性能十分关键,而每穗总粒数在一定范围内的波动将对双季杂交稻品种产量性能的自动调节性产生影响,千粒质量是一个遗传相对稳定的指标,其可塑性小[22-24]。各生育阶段恰当的施氮比例能在一定程度上协调产量构成因素之间的关系,提高水稻单产[25]。对冬小麦的研究表明,越冬期不施氮能明显降低冬小麦的有效穗数,拔节期不施氮对千粒质量影响较大,而抽穗期和成熟期不施氮则对穗粒数具有显著的影响,而影响冬小麦产量最为重要的构成因素为有效穗数,因而越冬期缺氮对冬小麦产量影响最大,为其氮素亏缺敏感期和需氮关键期[15]。表明冬小麦产量构成因素形成上对氮素亏缺响应存在生育阶段的差异性,且影响产量的关键因素为单株有效穗数。从本试验超级杂交晚稻品种五丰优T025不同生育阶段氮素亏缺对产量构成因素的影响来看,以分蘖期氮素亏缺处理下单株有效穗数最少,以幼穗分化期氮素亏缺处理下每穗总粒数最少,且该两指标在幼穗分化期前显示出较强的氮素亏缺补偿效应。结实率和千粒质量两指标均以全生育期不施氮处理最高,未发现结实率存在氮素亏缺补偿效应,而千粒质量氮素亏缺后补偿效应亦较小。从产量形成上仔细分析导致超级杂交晚稻不同生育阶段氮素亏缺处理下产量出现差异的原因可发现,单株有效穗和每穗总粒数最为重要,与产量相关最为紧密,相关系数分别为0.828 3*和0.911 6**,千粒质量和结实率与产量均无明显相关关系。表明高产稳产型超级杂交晚稻品种选育时应首先注重单株有效穗数,前提是必须确保具有较强的分蘖力,在此基础上可适当兼顾每穗总粒数,也就是一定穗型,即应取穗粒兼顾之径,这一点与冬小麦极为相似。不可过度追求大穗大粒,否则产量稳定性能得不到保障,特别是生产上氮肥管理不当时易出现单产大起大落现象。栽培上,超级杂交晚稻氮素管理上应注意生育前、中期氮素亏缺的敏感性及补偿效应至抽穗期之后的无效性,不能过分强调氮素后移,一般意义上讲,抽穗后不应再施氮肥。然而,在保障前、中期氮素供应充足的基础上,也应注意超级杂交晚稻后期脱氮问题,从本试验单株有效穗数结果可知,幼穗分化期氮素亏缺、抽穗期加倍恢复供氮较等量处理更高,进一步分析发现是其成穗率更高,群体质量更优。另外,需要指出的是,由于生长补偿效应需要一定的时间积累[9,11,15-16,26-28],超级杂交晚稻补偿效应总体上有一定的局限性,生产上既要合理利用补偿效应来弥补管理不当带来的损失,更要加强全程优化调控。
如前所述,总体看来,超级杂交晚稻各生育阶段氮素亏缺处理的ANUE、NUPE、NHI、NPE、PFP最低值均出现在处理T5至T10之间,说明保证分蘖期至抽穗期,尤其是幼穗分化期氮素的充分供应,控制籽粒乳熟后氮肥的使用对超级杂交晚稻ANUE、NUPE、NHI、NPE、PFP可起到优化的效果,这与当前认为的超级稻品种前氮适当后移,适度降低基追肥比例,重视穗粒肥等施氮理论与技术[17-19]基本一致。
致谢:朱昌兰,彭小松,贺晓鹏,傅军如,欧阳林娟,边建民,胡丽芳等课题组成员参与了本研究的试验工作,谨致感谢!
[1]Bond J A,Walker T W,Ottis B V,et al.Rice seeding and nitrogen rate effects on yield and yield components of two rice cultivars[J].Agronomy Journal,2008,100(2):393-397.
[2]杨京平,姜宁,陈杰.水稻吸氮量和干物质积累的模拟试验研究[J].植物营养与肥料学报,2002,8(3):318-324.
[3]夏晓亮,石祖梁,荆奇,等.氮肥运筹对稻茬小麦土壤硝态氮含量时空分布和氮素利用的影响[J].土壤学报,2010,47(3):490-496.
[4]樊剑波,张亚丽,王东升,等.水稻氮素高效吸收利用机理研究进展[J].南京农业大学学报,2008,31(2):129-134.
[5]Peng S B,Buresh R J,Huang J L,et al.Strategies for overcoming low agronomic nitrogen use efficiency in irrigated rice systems in China[J].Field Crops Research,2006,96(1):37-47.
[6]吴建富,潘晓华,石庆华,等.江西双季水稻施肥中存在的问题及对策[J].中国稻米,2012,18(5):33-35.
[7]庞桂斌,彭世彰.中国稻田施氮技术研究进展[J].土壤,2010,42(3):329-335.
[8]Ju X T,Xing G X,Chen X P,et al.Reducing environmental risk by improving N management in intensive Chinese agricultural systems[J].Proceedings of the National Academy of Sciences of the USA(PNAS),2009,106(9):3041-3046.
[9]翟丙年,李生秀.氮素对冬小麦产量的亏缺及补偿效应[J].西北农林科技大学学报:自然科学版,2001,29(3):53-56.
[10]曾祥明,韩宝吉,徐芳森,等.不同基础地力土壤优化施肥对水稻产量和氮肥利用率的影响[J].中国农业科学,2012,45(14):2886-2894.
[11]赵明,李建国,张宾,等.论作物高产挖潜的补偿机制[J].作物学报,2006,32(10):1566-1573.
[12]刑英英,张富仓,王秀康.不同生育期水分亏缺灌溉和氮营养对玉米生长的影响[J].干旱地区农业研究,2010,28(6):1-6.
[13]褚丽丽,张忠学.氮素营养与水分胁迫对大豆产量补偿效应的影响[J].生态学报,2010,30(10):2665-2670.
[14]马银山,杜国祯,张世挺.施肥和刈割对冷地早熟禾补偿生长的影响[J].生态学报,2010,30(2):279-287.
[15]翟丙年,李生秀.氮素对冬小麦生长发育及产量的亏缺和补偿效应[J].植物营养与肥料学报,2005,21(3):308-313.
[16]翟丙年,孙春梅,王俊儒,等.氮素亏缺对冬小麦根系生长发育的影响[J].作物学报,2003,29(6):913-918.
[17]蒋鹏,黄敏,Md Ibrahim,等.“三定”栽培对双季超级稻养分吸收积累及氮肥利用率的影响[J].作物学报,2011,37(12):2194-2207.
[18]陈爱忠,潘晓华,吴建富,等.氮素施用比例对双季超级稻产量和氮素吸收、利用的影响[J].中国土壤与肥料,2011,(3):40-44.
[19]邹应斌,陈玉枚,徐国生,等.双季超级稻产量和氮磷钾吸收的基因型差异[J].作物研究,2008,22(4):225-229.
[20]付景,杨建昌.超级稻高产栽培生理研究进展[J].中国水稻科学,2011,25(4):343-348.
[21]Peng S B,Tang Q Y,Zou Y B.Current status and challenges of rice production in China[J].Plant Prod Sci,2009,12(1):3-8.
[22]陈小荣,孙嘉,肖自京,等.不同产量潜力杂交早稻品种的群体自动调节力差异及其生理机制研究[J].江西农业大学学报,2012,34(5):851-858.
[23]陈小荣,肖自京,孙嘉,等.不同产量晚稻品种分蘖期动态密度稀化下群体自动调节力的差异与生理机制[J].中国水稻科学,2013,27(4):405-412.
[24]陈小荣,石庆华,潘晓华,等.金优463在赣中北作双季早稻的产量、产量构成因素分析与生产对策[J].江西农业大学学报,2006,26(2):161-163.
[25]凌启鸿,张洪程,戴其根,等.水稻精确定量施氮研究[J].中国农业科学,2005,38(12):2457-2467.
[26]陈小荣,刘灵燕,钟蕾,等.抽穗后干旱复水对双季杂交晚稻产量形成和叶片δ13C及内源激素水平的影响[J].核农学报,2013,27(2):240-246.
[27]陈小荣,孙嘉,肖自京,等.双季杂交早稻秧苗素质对产量及内源激素水平的影响[J].核农学报,2012,26(9):1298-1304.
[28]陈小荣,孙嘉,肖自京,等.2个杂交晚稻品种不同秧苗素质下产量及内源激素水平的差异性[J].江西农业大学学报,2014,36(3):463-469.
猜你喜欢
华北农学报(2022年4期)2022-09-14
中国农业科学(2022年8期)2022-05-16
东北农业大学学报(2020年12期)2021-01-15
江苏农业科学(2019年10期)2019-07-08
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
江苏农业科学(2016年11期)2017-03-21
湖南农业(2017年1期)2017-03-20
江苏农业科学(2014年8期)2014-10-23
作物研究(2014年6期)2014-03-01
