
不同训练方式建立大鼠空间记忆后海马结构中BDNF表达的变化*
2015-06-05 14:36于玮,谢雯,罗禹,张莉,△
华中科技大学学报(医学版) 2015年1期
于 玮,谢 雯,罗 禹,张 莉,△
1西安医学院生理学教研室,西安 710021
2西安交通大学医学院生理学与病理生理学系,西安 710061
3西安交通大学环境与疾病相关基因教育部重点实验室,西安 710061
不同训练方式建立大鼠空间记忆后海马结构中BDNF表达的变化*
于 玮1,谢 雯2,罗 禹3,张 莉2,3△
1西安医学院生理学教研室,西安 710021
2西安交通大学医学院生理学与病理生理学系,西安 710061
3西安交通大学环境与疾病相关基因教育部重点实验室,西安 710061
目的 研究短期强化训练能否建立可靠的空间长时记忆;用不同训练方式建立空间记忆后,大鼠海马结构中脑源性神经营养因子(BDNF)的表达是否会发生变化。方法 利用Morris水迷宫建立大鼠空间短期记忆、长期记忆和短期强化训练形成的长期记忆3种模型,用激光共聚焦显微镜观察正常(NC)组、短期记忆(ST)组、长期记忆组(LT)和短期强化训练形成的长期记忆(SRT)组大鼠海马各亚区的BDNF的分布情况。结果 Morris水迷宫定位航行实验中LT组和SRT组间寻找站台平均潜伏期和策略差异均无统计学意义;记忆检测发现,LT组大鼠除了在站台所在象限的停留时间明显长于SRT组,差异有统计学意义(P<0.05)外,两组大鼠寻找站台潜伏期和策略以及穿越站台次数差异均无统计学意义。LT组与NC组及ST组比较,海马齿状回和CA1区的BDNF免疫荧光反应明显增强,差异有统计学意义(均P<0.05);SRT组与NC组以及ST组比较,海马齿状回和CA1区的BDNF免疫荧光反应明显增强,差异有统计学意义(P<0.05或P<0.01)。LT组与SRT组之间,NC组与ST组之间,海马各区的BDNF表达差异均无统计学意义。结论 短期强化训练可建立与长期训练基本相同的长期记忆。BDNF与空间长期记忆的形成密切相关,其可能不参与空间短期记忆的形成或调节。
脑源性神经营养因子; 海马; 学习记忆
脑源性神经生长因子(brain-derived neurotrophic factor,BDNF)是一个小的二聚体蛋白质,是神经营养因子家族的一员,由其前体蛋白在细胞内高尔基体和(或)非成熟囊泡内裂解N端而成为成熟的BDNF[1],在中枢和周围神经系统神经元的生存、维持和生长中起重要的作用[2]。越来越多的证据表明,BDNF可以调节短期突触功能和活动依赖的突触可塑性,如长时程增强(long-term potentiation,LTP),BDNF还可快速增强海马神经元突触的传递和递质释放,从而影响学习记忆[3]。许多体外实验已表明,BDNF在海马LTP的诱导和维持中具有重要作用,提示BDNF在学习与记忆中也具有重要作用。已有研究发现,在大脑内注入反义BDNF寡核苷酸可以破坏某些形式的长期记忆的形成[45];而大脑过度表达BDNF的转基因小鼠在长期记忆中也表现出缺陷[6]。也有研究报道,BDNF突变小鼠在复合空间任务训练中表现出学习障碍,而在记忆保持上无障碍[7],因为急性或慢性大脑内注入BDNF在记忆保持上无任何作用。尽管目前关于BDNF在学习与记忆中的作用尚无统一结论,但普遍认为,BDNF可能参与学习和记忆的过程。本研究拟在不同训练方法建立空间记忆的基础上,研究大鼠海马结构中BDNF的分布情况,探讨其在空间短期记忆、长期记忆中的可能作用。
1 材料与方法
1.1 实验动物与分组
1.2 实验方法
1.2.1 空间记忆模型的建立 利用Morris水迷宫建立记忆模型。水迷宫为一直径120cm、高55cm的圆形水池,水池内壁被漆为黑色,池内水深42 cm,水温保持在(22±1)℃,房间内光照恒定,并保证无光线反射在水池内。池壁上以4个等距离点东(E)、南(S)、西(W)、北(N)将水池分为4个象限,分别称SW、NW、SE及NE象限,在NE象限正中,距离池壁35cm处放有一个直径为8cm,高40cm的圆形透明站台,站台低于水面2cm。迷宫上方安置着连接显示系统的摄像机。大鼠的运动轨迹及测试结果的显示和处理均采用中国科学院心理研究所研制开发的动物行为学软件。训练期间迷宫外参照物保持不变。
Morris水迷宫的测试主要包括定位航行实验(Place navigation)和空间探索实验(Spatial probe test)两个部分。定位航行实验用于检测大鼠对水迷宫学习的能力。空间探索实验用于测试大鼠学会寻找站台后,对站台空间位置记忆的能力。根据大鼠在Morris水迷宫中搜索站台的游泳轨迹,可判断其寻找水下站台的搜索策略。一般有4种策略方式:直线式(straight)、趋向式(tendency)、边缘式(marginal)及随机式(random)[810]。
直线式以大鼠入水后找到的第1个点与站台之间的连线为中轴,如果大鼠的所有运动轨迹点与中轴的距离均不超过半径的15%范围,且在设定区域中运动的时间至少占运动总时间的70%,则认定该运动策略为直线式。趋向式与直线式的策略相似,但范围为半径的50%。边缘式是以大鼠的运动区域中心为圆心,取半径的75%作一个圆,如果大鼠70%以上的时间均在该圆外活动,软件系统则判断大鼠此次的运动为边缘式策略。若大鼠的运动策略不能归为以上3种,则系统判定是随机式。
本实验参照Ramirez-Amaya[11]和Meiri[12]等的Morris水迷宫训练方法建立了短期记忆、长期记忆和短期强化训练3种训练模型。①ST组的训练:即定位航行实验,实验历时1d,分2个时间段,中间间隔1h,每个时间段训练5次,共训练10次,训练时将大鼠面向池壁放入水中,入水点是假性随机的,10次训练将大鼠依次从4个不同的入水点入水,每次潜伏期的值作为该训练时段的成绩,记为平均潜伏期,以便和其他两组进行比较。②LT组的训练:即定位航行实验,实验历时5d,每天分上、下午两个训练段,每段训练5次,共50次。每训练段5次潜伏期的平均值作为该训练段的平均潜伏期。每只大鼠的单次训练操作同ST组。③SRT组的训练:即定位航行实验,实验历时1d,与ST组相同。分上、下午两个训练段,每段训练25次,共50次。训练总次数与LT组相同。每相邻5次训练潜伏期的平均值作为该训练时段的成绩,记为平均潜伏期,进行统计分析。每只大鼠的单次训练操作同ST组。以上3组训练中,如果大鼠在60s内未找到站台,则将其引至站台,停留10s,这时潜伏期记为60s。训练采用大循环方式,找到站台者停留5s。每2次训练的最短间隔时间为10min。观察潜伏期,游泳速度,搜索策略。
以上3组大鼠训练结束后,均进行记忆的测试。ST组在训练结束后1.5h检测其短时记忆,LT和SRT组在训练结束后第7天检测其长时记忆[11],检测内容包括定位航行实验和空间探索实验两部分。定位航行实验:选择离站台最远的SW象限为入水点,将大鼠面向池壁放入水中,检测其寻找站台的潜伏期及搜索策略。空间探索实验:在定位航行实验结束最少10min后进行此项实验。将站台撤掉后,选择SW象限为入水点,将大鼠面向池壁放入水中,观察并记录60s内动物在原站台所在象限的停留时间、穿越原站台次数以及搜索策略。
1.2.2 BDNF的免疫荧光染色 在记忆模型建立好之后,即刻挑选学习成绩好的大鼠4只,进行免疫荧光染色。动物用200g/L氨基甲酸乙酯按9.5 mL/kg进行腹腔麻醉,40g/L多聚甲醛灌注固定,取全脑,修块,入20%蔗糖溶液4℃过夜。次日,冠状冰冻连续切片,片厚40μm,采用间接免疫荧光法进行BDNF的免疫荧光漂浮染色:0.3%Triton室温孵育30min;正常羊血清封闭1h;兔抗BDNF抗体(1∶50)孵育72h,4℃;FITC标记羊抗兔抗体孵育24h,4℃;甘油碳酸盐缓冲液封片。封片后立即用Leica激光共聚焦显微镜观察并摄片,全程避光。阴性对照用PBS替代一抗孵育,其它步骤相同。
1.3 统计学分析
第二年春,将甲树与乙树相连处右甲树的枝锯掉。(两棵树都只有小指粗),(如今,甲树的新枝(丙)和乙树都长到围粗20cm多了。)本年岁末,发现甲树B段接近乙树的地方长得很粗,而甲树B段中点以左部分一点也没长粗。
所有数据以¯x±s表示,ST、LT和SRT组间潜伏期的比较采用重复测量数据方差分析,站台所在象限停留时间的比较及积分光密度值组间比较采用单因素方差分析,两组大鼠寻找站台所用策略百分数采用Mann-Whitney秩和检验比较组间差异。数据处理应用SPSS 13.0统计软件,以P<0.05为差异有统计学意义。
2 结果
2.1 各组大鼠空间学习能力的比较
2.1.1 大鼠在训练时逃逸潜伏期的比较 3组动物在训练中,均随着训练时段的增加,在水中逃逸、寻找平台的潜伏期越来越短,整体曲线呈下降趋势,直至达到稳定水平。表明接受训练的3组大鼠通过学习,均可对水下的安全逃逸站台产生空间定位记忆。各组动物在不同训练时段的潜伏期见图1A。3组间的平均潜伏期总体比较差异有统计学意义(P<0.01),在1、2、3、8、10这5个时间段,ST组的潜伏期明显长于LT组,差异有统计学意义(P<0.05或P<0.01);在1、2、8、10这4个时间段,ST组的潜伏期明显长于SRT组,差异有统计学意义(P< 0.05或P<0.01);LT组和SRT组间的平均潜伏期差异没有统计学意义。
2.1.2 搜索策略的比较 ST组、LT组和SRT组的动物在训练中均较多地采用了趋向式和直线式的有效策略。各组动物在训练时的平均搜索策略见图1B。LT组和ST组相比,4种策略差异均有统计学意义(均P<0.05),LT组采用趋向式策略和直线式策略较ST组多,而ST组采用边缘式策略和随机式策略较LT组多。ST组和SRT组相比除趋向式策略差异没有统计学意义以外,边缘式、随机式和直线式策略差异均有统计学意义(均P<0.05),SRT组采用直线式策略较ST组多,而ST组采用边缘式策略和随机式策略较SRT组多。LT组和SRT组相比,4种策略差异均没有统计学意义。
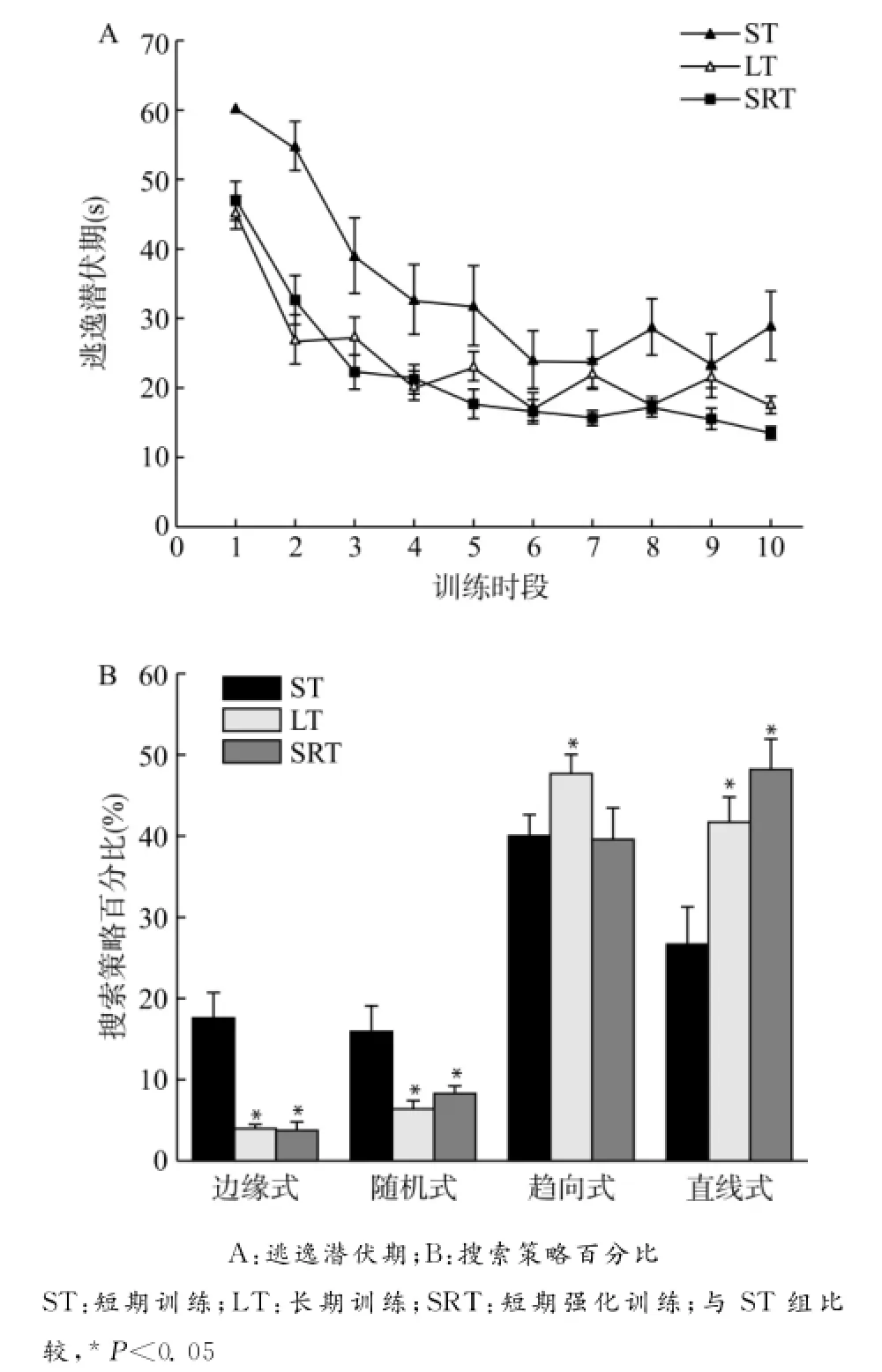
图1 各组大鼠训练时逃逸潜伏期的变化及采用不同搜索策略的百分比Fig.1 Changes of escape latency and the percentage of different target-seeking strategies during training in each group
2.2 各组大鼠空间记忆能力的比较
2.2.1 大鼠搜索站台的潜伏期及搜索策略 在训练结束后1.5h检测ST组大鼠的搜索站台的潜伏期及搜索策略;在训练结束后第7天检测LT组和SRT组大鼠搜索站台的潜伏期及搜索策略,结果发现:3组动物搜索站台的平均潜伏期相比差异均有统计学意义(均P<0.01)(图2A),ST组与SRT组比较,其搜索站台的平均潜伏期和搜索策略差异有统计学意义(均P<0.01)(图2B),SRT组搜索站台的平均潜伏期短,且多使用直线式和趋向式的搜索策略;ST组与LT组比较,其搜索站台平均潜伏期和搜索策略差异有统计学意义(P<0.01),LT组搜索站台的平均潜伏期短,且多使用直线式和趋向式的有效搜索策略;LT组与SRT组比较,其搜索站台的平均潜伏期和搜索策略差异没有统计学意义。
2.2.2 大鼠穿越站台次数以及在原站台所在象限的停留时间 在训练结束1.5h后检测ST组大鼠穿越站台次数以及在原站台所在象限的停留时间,在训练结束后第7天检测LT组和SRT组大鼠穿越站台次数以及在原站台所在象限的停留时间。结果发现:3组相比,大鼠穿越站台次数以及在原站台所在象限的停留时间差异均有统计学意义(均P<0.01);ST组和LT组大鼠比较,穿越站台次数以及在原站台所在象限的停留时间差异均有统计学意义(均P<0.01),ST组比LT组大鼠穿越站台次数以及在原站台所在象限的停留时间均少;ST组和SRT组大鼠比较穿越站台次数差异有统计学意义(P<0.01),但两组站台所在象限停留时间相比差异没有统计学意义。LT组比SRT组在站台所在象限停留时间长,差异有统计学意义(P<0.05),比较两组间站台穿越次数,差异没有统计学意义(图2 C、D)。
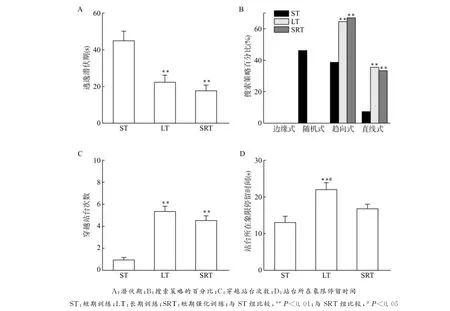
图2 各组大鼠检测时的平均潜伏期、采用不同搜索策略的百分比、穿越站台次数以及站台所在象限停留时间Fig.2 Average latency,percentage of different target-seeking strategies and times of crossing target area and length of stay in each quadrant during detection in each group
2.3 空间记忆对BDNF表达的影响
LT组与NC组以及ST组比较,LT组在齿状回和CA1区的BDNF的积分光密度明显增强,差异均有统计学意义(均P<0.05);CA3区的BDNF积分光密度虽有增强,但与NC组及ST组相比差异没有统计学意义;SRT组与NC组以及ST组比较,其在齿状回和CA1区的BDNF积分光密度明显增强,差异有统计学意义(P<0.05或P<0.01),CA3区的BDNF积分光密度虽有增强,但与NC组及ST组相比差异没有统计学意义。LT组与SRT组比较,海马各个亚区的BDNF积分光密度差异均没有统计学意义;NC组与ST组比较,海马各个区的BDNF积分光密度差异也均没有统计学意义,见表1,图3~5。
表1 各组大鼠海马结构BDNF积分光密度(IOD)比较s,n=4)Table1 The integral absorbance value of BDNF in rat hippocampus in each group(s,n=4)
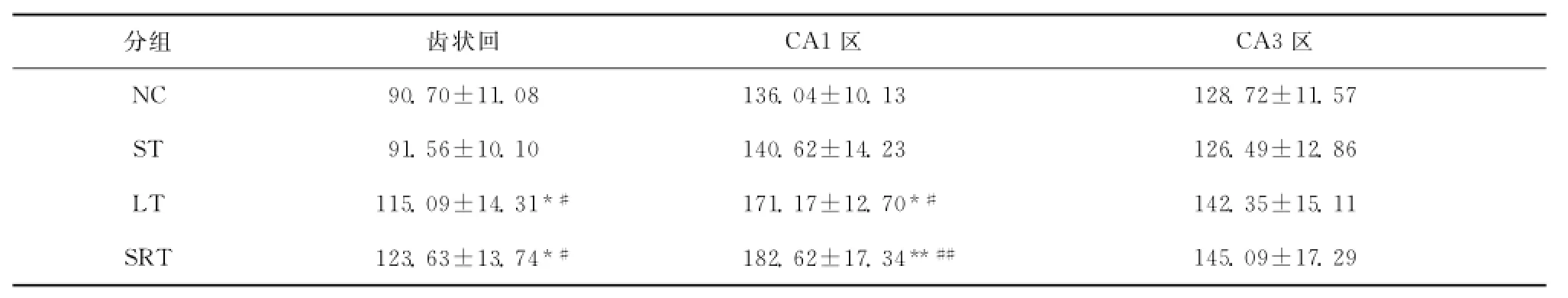
表1 各组大鼠海马结构BDNF积分光密度(IOD)比较s,n=4)Table1 The integral absorbance value of BDNF in rat hippocampus in each group(s,n=4)
与NC组比较,*P<0.05**P<0.01;与ST组比较,#P<0.05##P<0.01
?

图3 各组大鼠齿状回BDNF的免疫荧光染色(标尺=40μm)Fig.3 Immunofluorescence staining for BDNF expression in the dentate gyrus of hippocampus in different groups(Scale bar=40μm)

图4 各组大鼠CA1区BDNF的免疫荧光染色(标尺=40μm)Fig.4 Immunofluorescence staining for BDNF expression in the CA1area of hippocampus in different groups(Scale bar=40μm)
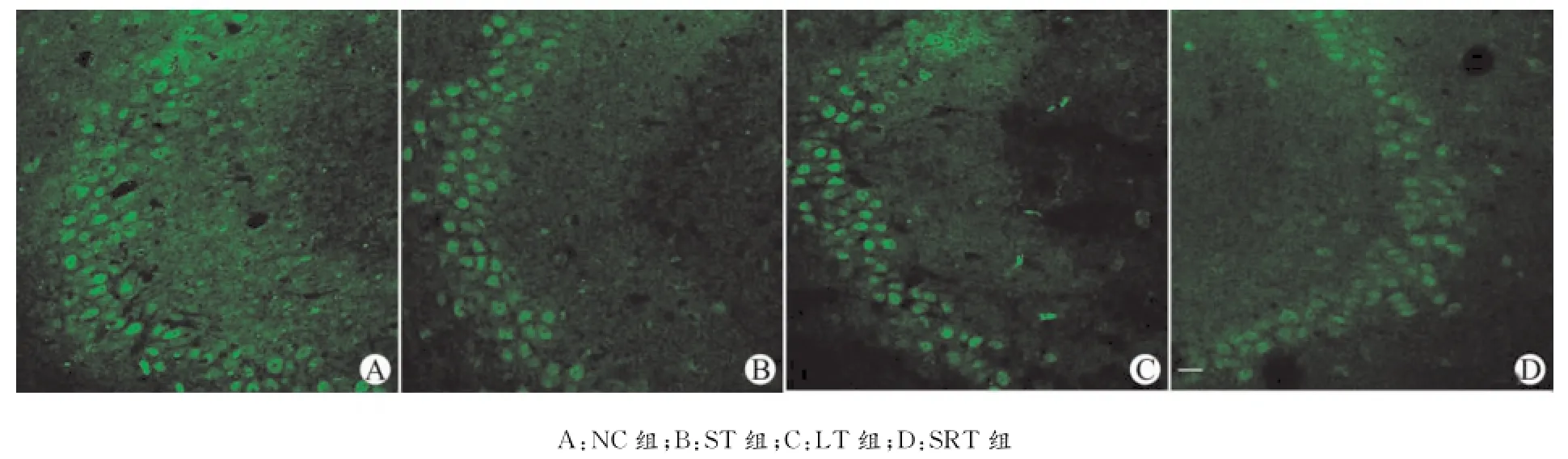
图5 各组大鼠CA3区BDNF的免疫荧光染色(标尺=40μm)Fig.5 Immunofluorescence staining for BDNF expression in the CA3area of hippocampus in different groups(Scale bar=40μm)
3 讨论
3.1 短期强化训练可建立动物空间长时记忆
经典的Morris水迷宫实验一般采用4~6个训练日,训练后可建立空间长时记忆[13]。本研究参照Ramirez-Amaya[11]和Meiri等[12]的Morris水迷宫训练方法分别建立大鼠空间长时记忆和空间短时记忆;并使用短时程、高频率的短期强化训练方法,试图建立大鼠的空间长时记忆。即在1个训练日的10个训练段内完成50次训练,然后间隔7d检测动物对原站台空间位置的记忆能力。结果发现,1个训练日的短期强化训练可使大鼠建立起与经典训练模式中5个训练日所建立的空间记忆基本相同的长时记忆。比较两组动物长时记忆的测试结果发现,两组大鼠的定位航行实验的潜伏期、搜索策略以及空间探索实验的穿越原站台次数差异均无统计学意义,但LT组大鼠在原站台所在象限的停留时间明显长于SRT组。这表明LT组动物所建立的空间长时记忆略优于SRT组。该研究结果表明,对大鼠进行Morris水迷宫短期强化训练可以建立空间长时记忆。提示增加训练频率可以提高学习效率,1个训练日的强化训练可以用作建立动物空间长时记忆的训练模式[14]。在学习的过程中,训练时间分布的方式起重要作用,训练的最小时间间隔对于所获得的信息的完全整合是必需的。学习后储存的信息,如不经常提取和再巩固,大部分信息会随着时间而渐渐消失[15]。一定的训练次数和训练间隔对于长期记忆的形成是必需的,本实验模型建立过程中,各组大鼠每次训练的时间间隔至少10min,这对记忆的形成是有效的[1617]。
3.2 BDNF与空间长期记忆密切相关
BDNF是神经营养因子家族的主要成员之一,在成年哺乳动物大脑中含量较高,尤其是在参与学习、记忆、可塑性的脑区,如海马和皮层等。药理学和基因学的实验已经表明,BDNF和TrkB受体的结合是海马LTP产生的基础,BDNF参与海马LTP的维持。在缺乏BDNF的转基因小鼠,海马LTP不易产生,在出生后脑前区缺乏TrkB受体的选择性敲除小鼠,表现出海马LTP的减弱和学习行为的损害。本研究结果表明:BDNF在海马结构的多个亚区广泛表达,提示BDNF与海马功能密切相关。
BDNF在学习和记忆中的作用可以通过应用抗BDNF抗体等多种方法进行研究。敲除年老大鼠BDNF基因后,其体内BDNF mRNA的表达也降低,而且表现出严重的空间学习记忆损害,这些说明BDNF对于空间学习是必需的。本研究结果表明:形成长期记忆的两组大鼠尽管其训练方式不同,但海马结构的齿状回及CA1区均有BDNF的显著高水平表达,而在NC组和ST组,海马各区BDNF的表达水平无明显变化。因此本研究表明BDNF与空间长期记忆的形成密切相关,可能不参与空间短期记忆的形成或调节。Mu等[4]报道,在水迷宫测试中,给大鼠脑内注入抗BDNF的抗体可引起大鼠空间学习的损害,也有研究指出在被动回避学习中,保持力表现良好的大鼠与保持力表现差的大鼠比较,海马齿状回的BDNF mRNA的水平显著增加[18]。Alonso等[19]研究表明,在抑制性回避训练中,BDNF对于长期记忆的形成是必需的。Linnarsson等[7]报道,用BDNF基因敲除小鼠进行Morris水迷宫训练,结果表明基因敲除鼠与正常鼠相比学习能力明显受损;也有实验表明,在水迷宫训练的第3天和第6天后,学习组和对照组相比,大鼠海马的BDNF mRNA水平升高。这些研究结果均与本研究结果一致。也有研究表明,在抑制性回避训练中,于训练前给大鼠海马CA1区注入抗BDNF抗体可阻断短期记忆的形成[19],这表明在该实验中,BDNF对于短期记忆的形成是必需的。这个研究结果与本研究结果不同,有可能是BDNF在不同形式的学习记忆过程中发挥作用的通路不相同,也可能与检测BDNF水平的时间不同有关。
3.3 参与空间长期记忆的脑区
Becker等[20]用损毁的方法观察不同脑区在空间学习记忆中的作用,发现只要海马系统完好,大鼠就可精确地完成空间迷宫任务,在灵长类动物身上也观察到,海马损害可以导致空间记忆受损[21],所以海马是与空间记忆能力密切相关的重要脑区。本研究结果显示,形成空间长期记忆的大鼠海马齿状回的颗粒细胞内BDNF显著高表达,海马CA1区的锥体细胞内BDNF也显著高表达;海马CA3区的锥体细胞内BDNF未出现表达水平的改变。以上结果表明,海马齿状回可能是空间长期记忆形成的关键脑区,海马CA1区也可能参与了空间长期记忆的形成,海马齿状回和CA1区可能与空间短期记忆关系不大;海马CA3区可能未参与空间短期记忆和空间长期记忆的形成。Okada等[22]和Nakao等[23]的研究表明,与空间学习能力密切相关的脑区主要是齿状回。在被动回避训练中,于记忆巩固之前给大鼠海马齿状回内连续5次注入反义BDNF寡核苷酸,可明显损害大鼠的记忆保持力[19],用原位杂交的方法发现,在依赖海马的某些学习过程中,大鼠海马CA1区的BDNF出现快速和选择性表达[2425]。这些研究结果与我们的研究结果一致。但也有研究表明,在抑制性回避训练中,给大鼠海马CA1区内注入BDNF抗体可阻断短期记忆的形成,提示海马CA1区可能与短期记忆有关,这与本研究结果有矛盾,可能与记忆的形式不同有关。
[1] Cattaneo A,Bocchio-Chiavetto L,Zanardini R,et al.BDNF Val66Met polymorphism and protein levels in amniotic fluid[J].BMC Neurosci,2010,11(2):11-16.
[2] Henderson C E.Role of neurotrophic factors in neuronal development[J].Curr Opin Neurobiol,1996,6(1):64-70.
[3] Musumeci G,Sciarretta C,Rodríguez-Moreno A,et al.TrkB modulates fear learning and amygadalar synaptic plasticity by specific docking sites[J].J Neurosci,2009,29(32):10131-10143.
[4] Mu J S,Li W P,Yao Z B,et al.Deprivation of endogenous brain-derived neurotrophic factor results in impairment of spatial learning and memory in adult rats[J].Brain Res,1999,835(2):259-265.
[5] Johnston A N,Clements M P,Rose S P.Role of brain-derived neurotrophic factor and presynaptic proteins in passive avoidance learning in day-old domestic chicks[J].Neuroscience,1999,88(4):1033-1042.
[6] Croll S D,Suri C,Compton D L,et al.Brain-derived neurotrophic factor transgenic mice exhibit passive avoidance deficits,increased seizure severity and in vitro hyperexcitability in the hippocampus and entorhinal cortex[J].Neuroscience,1999,93(4):1491-1506.
[7] Linnarsson S,Björklund A,Ernfors P.Learning deficit in BDNF mutant mice[J].Eur J Neurosci,1997,9(12):2581-2587.
[8] 隋南,陈双双,匡培梓.海马结构、前额皮层或尾壳核损毁对大鼠空间认知能力的影响[J].心理学报,1992,25(4):81-87.
[9] DiMattia B D,Kesner R P.Spatial cognitive maps:differential role of parietal corte and hippocampal formation[J].Behav Neurosci,1988,102(4):471-480.
[10] Yoerg S J,Kamil A C.Response strategies in the radial arm maze:running around in circles[J].Animal Learn Behav,1982,10(4):530-534.
[11] Ramirez-Amaya V,Balderas I,Sandoval J,et al.Spatial longterm memory is related to mossy fiber synaptogenesis[J].J Neurosci,2001,21(18):7340-7348.
[12] Meiri N,Rosenblum K.Lateral ventricle injection of the protein synthesis inhibitor anisomycin impairs long-term memory in a spatial memory task[J].Brain Res,1998,789(1):48-55.
[13] 胡镜清,温泽淮,赖世隆.Morris水迷宫检测的记忆属性与方法学初探[J].广州中医药大学学报,2000,17(2):117-119.
[14] 张莉,于玮,韩太真,等.不同训练方式建立大鼠空间记忆后海马结构NMDA受体表达的变化[J].生理学报,2006,58(5): 442-448.
[15] Sara S J.Retrieval and reconsolidation:toward a neurobiology of remembering[J].Learn Mem,2000,7(2):73-84.
[16] Yin J,Del Vecchio M,Zhou H,et al.CREB as a memory modulator:induced expression of a dCREB2activator isoform enhances long-term memory in Drosophila[J].Cell,1995,81(1):107-115.
[17] David G,Ursula H,Marlen K,et al.Protein phosphatase 1is a molecular constraint on learning and memory[J].Nature,2002,418(29):970-975.
[18] Ma Y L,Wang H L,Wu H C,et al.Brain-derived neurotrophic factor antisense oligonucleotide impairs memory retention and inhibits long-term potentiation in rats[J].Neuroscience,1998,82(4):957-967.
[19] Alonso M,Vianna M R,Depino A M,et al.BDNF-triggered events in the rat hippocampus are required for both short-and long-term memory formation[J].Hippocampus,2002,12(4): 551-560.
[20] Becker J T,Walker J A,Olton D S.Neuroanatomical bases of spatial memory[J].Brain Res,1980,200(2):307-320.
[21] Redish A D,Touretzky D S.Cognitive maps beyond the hippocampus[J].Hippocampus,1997,7(1):15-35.
[22] Okada T,Yamada N,Tsuzuki K,et al.Long-term potentiation in the hippocampal CA1area and dentate gyrus plays different roles in spatial learning[J].Eur J Neurosci,2003,17(2): 341-349.
[23] Nakao K,Ikegaya Y,Yamada M K,et al.Spatial performance correlates with long-term potentiation of the dentate gyrus but not of the CA1region in rats with fimbria-fornix lesions[J].Neurosci Lett,2001,307(3):159-162.
[24] 张远康,陶连方.电针对脑缺血再灌注大鼠学习记忆能力及血管内皮生长因子表达的影响[J].华中科技大学学报:医学版,2013,42(1):70-73.
[25] Hall J,Thomas K L,Everitt B J.Rapid and selective induction of BDNF expression in the hippocampus during contextual learning[J].Nat Neurosci,2000,3(6):533-535.
(2014-05-15 收稿)
Changes of BDNF Expression in Rat Hippocampus after Establishment of Spatial Memory by Different Trainings
Yu Wei1,Xie Wen2,Luo Yu3et al
1Department of Physiology,Xi’an Medical University,Xi’an 710021,China
2Department of Physiology and Pathophysiology;
3Key Laboratory of Environment and Gene Related Diseases of the Ministry of Education,Medical School of Xi’an Jiaotong University,Xi’an 710061,China
Objective To investigate whether short reinforcing training can establish stable spatial long-term memory and how the brain-derived neurotrophic factor(BDNF)expression in the hippocampus changes after the establishment of spatial memory with different trainings.Methods Long-term memory was established by spatial short-term memory training(ST group),spatial long-term memory training(LT group)or short reinforcing training(SRT group)in Morris water maze.Laser scanning confocal microscopy was used to observe the distribution of BDNF in each area of the hippocampus in normal control group(NC),ST,LT and SRT groups.Results Morris water maze testing showed that there was no significant difference in the mean latency and the target-seeking strategies between the LT and SRT groups.The examination of the memories following different trainings indicated that the mean latency and the target-seeking strategies as well as the times of crossing the target in LT group were not significantly different from those in SRT group,except that the mean time remaining in the platform quadrant in LT group was significantly longer than that in SRT group(P<0.05).The integral absorbance(A)value of BDNF in the dentate gyrus(DG)and CA1area of the hippocampus was significantly increased in LT and SRT groups compared with that in NC and ST groups(P<0.05or P<0.01).In addition,no significant difference in the integral Avalue of BDNF in CA1,CA3and DG of the hippocampus was observed between NC and ST groups or between LT and SRT groups.Conclusion The short reinforcing training can establish long-term spatial memory,which is almost the same as that established by the long-term training.BDNF is closely associated with spatial long-term memory formation and it may not be involved in the formation or regulation of spatial short-term memory.
brain-derived neurotrophic factor; hippocampus; learning and memory
G804.7,R49
10.3870/j.issn.1672-0741.2015.01.003
*教育部中国博士后科学基金资助项目(No.2013M540760)
于 玮,女,1978年生,硕士研究生,讲师,E-mail:marine_wei@163.com
△通讯作者,Corresponding author,E-mail:zhangli1998@mail.xjtu.edu.cn
猜你喜欢
现代食品科技(2022年8期)2022-09-02
作文周刊·小学二年级版(2022年20期)2022-05-05
中学生数理化·高一版(2022年3期)2022-04-05
公民与法治(2020年21期)2020-11-26
初中生学习指导·提升版(2020年10期)2020-09-10
公民与法治(2020年11期)2020-07-25
上海节能(2020年3期)2020-04-13
创新作文(小学版)(2019年10期)2019-09-25
下一代英才(酷炫少年)(2018年6期)2018-07-09
中学生数理化·七年级数学人教版(2017年4期)2017-07-08
