
电针对脑缺血大鼠海马神经元microRNA132和p250GAP蛋白表达的影响
2016-03-13 08:46郑彩霞黄晓琳陆敏张凤霞郭雅碧韩肖华
中国康复 2016年6期
郑彩霞,黄晓琳,陆敏,张凤霞,郭雅碧,韩肖华
血管性痴呆(vascular dementia, VaD)是指由缺血性卒中、出血性卒中和造成脑区低灌注的脑血管疾病所致的严重认知功能障碍综合征。在全世界范围内,VaD是仅次于阿尔茨海默氏病(Alzheimer's disease, AD)的第二大痴呆类型,约占痴呆总数的20%~30%[1]。近年来,越来越多的临床研究及实验研究证实针灸对VaD引起的学习记忆功能下降具有明显的保护和改善作用,但其具体作用机制尚不明确[2-3]。韩肖华等[4]发现电针(electroacupuncture, EA)刺激百会穴和大椎穴可以提高脑缺血大鼠Morris水迷宫成绩,机制可能与EA刺激可能激活海马区PKA/CREB信号通路,上调其下游的Bcl-2靶基因转录水平,使翻译成的Bcl-2蛋白量增多,进而发挥抗神经元凋亡作用相关;Li等[5]在后来的研究中也表明EA可能是通过cAMP/PKA/CREB通路改善多发性脑梗塞大鼠的学习记忆功能。研究表明,miR-212/132可能受转录因子CREB(cAMP-response element binding)[6]和转录抑制因子REST/NRSF(Repressor Element 1 silencing transcription factor/neuron-restrictive silencer factor)调控[7],其中CREB存在于神经细胞中,而REST存在于小鼠胚胎成纤维细胞中。miR132和miR212串联基因的正常表达是神经元发育、成熟、发挥正常功能的必要条件,它们的异常表达会引起一些神经系统疾病,如AD和tau疾病[8]。由于海马区miR-212/132成熟产物主要是miR132,miR132被认为在成年小鼠海马区新生神经元的突触生长和分支中发挥重要作用[9]。miR132可能通过抑制GTP酶活化蛋白p250GAP而促进树突棘成熟,影响突触形成[10]。本实验旨在初步探讨EA是否通过激活PKA/CREB信号通路影响miR132及其下游p250GAP蛋白表达,为EA改善学习记忆障碍的内在机制提出新的证据。
1 材料与方法
1.1 材料 ①实验动物:健康成年SPF(Specefic pathogen Free)级SD(Sprague-Dawley)雄性大鼠90只,体质量250±20g,由湖南斯莱克景达实验动物有限公司提供(许可证号:SCXK湘2011-0003)。②主要试剂:H89(Sigma-Aldrich, Shanghai, China)、山羊抗大鼠p250GAP多克隆抗体(1∶200, Santa Cruz Biotechnology, Santa Cruz, CA, USA),小鼠抗大鼠β-tubulin单克隆抗体(1∶1000, Affinity Bioscience, USA)、辣根过氧化物酶(HRP)标记抗体(1∶5000; Proteintech Group Inc., Wuhan, China)、增强化学发光系统(an enhanced chemiluminescence system, ECL, Millipore, Billerica, MA, US)、反转录试剂盒(TOYOBO First Strand cDNA Synthesis Kit,USA)、定量PCR试剂盒(TOYOBO THUNDERBIRD SYBR qPCR Mix,USA)。③主要材料与仪器:电针仪(G6805-II型,上海)、毫针(华佗牌,30号)、脑立体定位仪(深圳沃瑞德科技有限公司)、微量进样器、PVDF膜(Merck Millipore, Germany)、荧光定量PCR仪(StepOneTMReal-Time PCR System, Life Technologies, New York, USA)。
1.2 方法 ①动物分组及处理:90只动物随机分为假手术组、模型组、EA组、EA+H89组和EA+NS组,每组18只,每组又分为7,14,21d 三个观察时间点,即每个时间点6只SD大鼠。②侧脑室注射:在制作2VO模型前1h,EA+H89组和EA+NS组的大鼠分别侧脑室注射H89(2μg/μL)或NS 10μL。侧脑室定位坐标:前囟后0.8mm,前囟右侧1.5mm,硬脑膜下约4.0mm。14、21d组的大鼠每间隔1周重复侧脑室注射H89或NS一次。③模型制备:腹腔注射10%水合氯醛(3ml/kg)麻醉大鼠后,行颈部正中约2cm切口,并游离一小段颈总动脉,分别结扎其近端和远端并剪断中部,以完全阻断双侧颈总动脉血流;假手术组大鼠双侧颈总动脉只暴露,不接扎和剪断。缝合切口,消毒皮肤,术后常规饲养。④EA参数:GV20位于顶骨正中,GV14位于背部正中第7颈椎与第1胸椎间。以30号1寸毫针斜刺入GV20 10mm,直刺入GV14 5mm。将针柄分别连接至电针仪上,GV20接负极,GV14接正极,选取连续波,频率20Hz,电流强度以观察到大鼠眼睑轻微颤动为宜。每天1次,每次20min。
1.3 评定标准 ①RT-PCR:直接断头法取出大脑后快速在冰上分离出双侧海马组织。取100mg组织,加入1ml Trizol Reagent彻底匀浆,随后加入氯仿和异丙醇,抽提总RNA。取10μl水溶RNA,依次加入特异性反转录引物U6-RT(10mM),miR-132-5p-RT(10mM)各1μl,及4μl 5×buffer,2μl 10mM dNTPs,1μl RNA inhibitor和1μl 反转录酶,用枪抽吸混匀后于PCR仪上进行逆转录反应。配制10μl反应体系,每个反转录产物配制3管,进行PCR扩增。选用U6 RNA作为内源参考基因。采用2-ΔΔCT法计算miR132表达的相对变化。引物由Invitrogen Biotechnology Co., LTD中国公司合成。miR132的引物:miR-132-5p-S:5′-ACCGTGGCTTTCGATTGTTAC-3′,通用下游:5′-TGGTGTCGTGGAGTCG-3′;U6的引物:R-U6-S:5′-CCTGCTTCGGCAGCACA-3′,R-U6-A:5′-AACGCTTCACGAATTTGCGT-3′。②Western blot:取100mg组织,与裂解液混合并匀浆,提取总蛋白液。每孔总蛋白上样量约60μg,行十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electropheresis, SDS-PAGE),电泳完成后将蛋白质转移至PVDF膜上。将PVDF膜用5%脱脂奶粉/TBS封闭液室温振荡封闭1h后,加入适当稀释的一抗4℃下过夜。滴加辣根过氧化物酶标记二抗,在37℃下孵育1h,并采用ECL显色系统显色。采用Image-J软件分析目标蛋白及内参蛋白的灰度值并计算比值。所有测定重复至少3次。

a代表假手术组;b代表模型组;c代表EA组;d代表EA+H89组;e代表EA+NS组
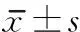
2 结果
2.1 大鼠海马区miR132的表达 同时间点各组大鼠间的miR132表达量相比,7、14、21d时,模型组大鼠miR132的表达量较假手术组均显著下降(P<0.05);EA组较模型组miR132的表达量均显著增多(P<0.05);EA+H89组较EA+ NS组miR132的表达量均显著下降(P<0.05)。同组别3个不同时间点之间miR132表达量相比,模型组、EA组随着时间延长,miR132的表达量有逐渐下降的趋势,但各时间点间差异无统计学意义;EA+NS组大鼠在14d时miR132的表达量较7d显著增加,但21d时miR132的表达量较14d显著减少(P<0.05)。见表1。
2.2 大鼠海马区p250GAP蛋白的表达 同时间点各组大鼠间的p250GAP蛋白表达量相比,7、14、21d时,模型组与假手术组大鼠相比,海马区p250GAP蛋白量均显著增多(P<0.05),EA组与模型组大鼠相比,海马区p250GAP蛋白量均显著减少(P<0.05),且14d时,EA组与假手术组大鼠相比,海马区p250GAP蛋白量亦显著减少(P<0.05);EA+H89组与EA+ NS组比较,海马区p250GAP蛋白量均明显增多(P<0.05)。见表1。同时间点各组大鼠间的p250GAP蛋白上样顺序及典型条带见图1。

组别时间miR132p250GAP蛋白假手术组7d1.00±0.010.16±0.0114d1.00±0.070.35±0.0221d1.00±0.030.18±0.02模型组7d0.67±0.04a0.31±0.04a14d0.61±0.03a0.56±0.04a21d0.59±0.06a0.32±0.04aEA组7d1.08±0.08b0.18±0.01b14d1.07±0.02b0.21±0.02ab21d0.90±0.07b0.19±0.01bEA+H89组7d0.77±0.04c0.39±0.04c14d0.84±0.03c0.50±0.01c21d0.66±0.04c0.35±0.07cEA+NS组7d1.05±0.070.22±0.0414d1.52±0.04d0.32±0.0421d0.91±0.02e0.20±0.01
与同时间点假手术组比较,aP<0.05;与同时间点模型组比较,bP<0.05;与同时间点电针+NS组比较,cP<0.05。与同组7d组比较,dP<0.05;与同组14d组比较,eP<0.05
3 讨论
miRNA主要通过降解靶mRNA或抑制靶mRNA翻译参与细胞转录后调节,在细胞增殖、分化、凋亡、代谢、胚胎发育等一系列生命过程中起重要作用[11],同时参与许多疾病的病理过程。Dhar等[10]发现肥胖基因Leptin可以通过促进海马神经元miR132表达抑制GTP酶活化蛋白p250GAP而促进树突棘成熟,影响突触形成。另一项研究也表明,BDNF可能通过调控miR132及p250GAP蛋白的表达促进视网膜神经节细胞(retinal ganglion cells, RGC)的轴突分枝[12]。miR132的上调会特异性的定位于动态突触,在长时程突触激活和记忆过程中发挥重要作用[13]。本研究发现,在7、14、21d时,脑缺血会引起大鼠海马组织miR132水平显著下降,同时伴有p250GAP蛋白显著增多;EA治疗可以明显促进miR132上调, p250GAP蛋白下调,提示EA可能通过对miR132及p250GAP蛋白的调控影响突触形成,这可能是电针改善脑缺血后学习记忆功能障碍的作用机制之一。模型组、EA组三个不同时间点相比时,随着时间延长,miR132的表达量有逐渐下降的趋势,提示随着缺血缺氧时间延长,miR132的下降会进行性加重,尽管EA组较同时间点模型组大鼠miR132的下降的严重程度已显著缓解,但并不能完全逆转或遏制miR132的下降水平。在western blot实验中,电泳条件及曝光时间对免疫沉淀复合物的相对光密度影响较大,此次实验并不能提供同组别不同时间点标本的电泳图,若直接使用同时间点不同组别样品上样后的实验结果进行比较没有实际意义,这也是下一步有待完善之处。
蛋白激酶A(protein kinase A,PKA)、Ca2+/钙调蛋白激酶IV(Calcium/calmodulin-dependent protein kinase type IV, CaMK IV)等都能通过磷酸化CREB参与学习记忆形成机制[14],其中,大量研究表明,PKA/CREB信号通路对学习记忆过程至关重要[15]。为了验证PKA/CREB信号通路激活是否可能涉入EA对miR132的调节作用,我们给予PKA特异性抑制剂-H89后观察EA的上述作用是否会被抑制。结果表明,侧脑室注射H89的大鼠,虽然按同样方法在手术次日开始进行EA治疗,但在7、14、21d时, 相比侧脑室注射NS的大鼠,EA对miR-132、p250GAP蛋白有利的调节作用均受到抑制,提示PKA/CREB信号通路可能是EA调控miR132表达及作用的内在机制之一。
综上所述,电针刺激百会穴和大椎穴7、14或21d时均可以调控miR-132及p250GAP蛋白的表达,其内在机制可能与PKA/CREB信号通路激活相关。但EA能否因此调控海马神经元树突生长和分枝,以及动态突触形成等,进而发挥改善突触可塑性和学习记忆功能的作用还有待下一步深入研究。
[1] Gorelick P B, Nyenhuis D. Understanding and treating vascular cognitive impairment [J]. Continuum (Minneap Minn), 2013, 19 (2): 425- 437.
[2] Shi G X, Liu C Z, Guan W, et al. Effects of acupuncture on Chinese medicine syndromes of vascular dementia [J]. Chin J Integr Med, 2014, 20 (9): 661-666.
[3] Yu J, Zhang X, Liu C, et al. Effect of acupuncture treatment on vascular dementia [J]. Neurological Research, 2006, 28 (1): 97-103.
[4] Han X, Zhao X, Lu M, et al. Electroacupuncture Ameliorates Learning and Memory via Activation of the CREB Signaling Pathway in the Hippocampus to Attenuate Apoptosis after Cerebral Hypoperfusion [J]. Evidence-Based Complementary and Alternative Medicine, 2013, 20(1): 1-8.
[5] Li Q Q, Shi G X, Yang J W, et al. Hippocampal cAMP/PKA/CREB is required for neuroprotective effect of acupuncture [J]. Physiol Behav, 2015, 13(9): 482-490.
[6] Vo N, Klein M E, Varlamova O, et al. A cAMP-response element binding protein-induced microRNA regulates neuronal morphogenesis [J]. Proc Natl Acad Sci U S A, 2005, 102 (45):16426-16431.
[7] Conaco C, Otto S, Han J J, et al. Reciprocal actions of REST and a microRNA promote neuronal identity [J]. Proc Natl Acad Sci U S A, 2006, 103 (7):2422-2427.
[8] Wang W X, Huang Q, Hu Y, et al. Patterns of microRNA expression in normal and early Alzheimer's disease human temporal cortex: white matter versus gray matter[J]. Acta Neuropathol, 2011, 121 (2): 193-205.
[9] Magill S T, Cambronne X A, Luikart B W, et al. microRNA-132 regulates dendritic growth and arborization of newborn neurons in the adult hippocampus [J]. Proc Natl Acad Sci U S A, 2010, 107 (47): 20382-20387.
[10] Dhar M, Zhu M, Impey S, et al. Leptin Induces Hippocampal Synaptogenesis via CREB-Regulated MicroRNA-132 Suppression of p250GAP[J]. Molecular Endocrinology, 2014, 28 (7):1073-1087.
[11] Fasanaro P, Greco S, Ivan M, et al. microRNA: emerging therapeutic targets in acute ischemic diseases [J]. Pharmacol Ther, 2010, 125(1): 92-104.
[12] Marler K J, Suetterlin P, Dopplapudi A, et al. BDNF Promotes Axon Branching of Retinal Ganglion Cells via miRNA-132 and p250GAP[J]. Journal of Neuroscience, 2014, 34 (3): 969-979.
[13] Nudelman A S, Dirocco D P, Lambert T J, et al. Neuronal activity rapidly induces transcription of the CREB-regulated microRNA-132, in vivo[J]. Hippocampus, 2010, 20 (4): 492-498.
[14] Bell K F S, Bent R J, Meese-Tamuri S, et al. Calmodulin Kinase IV-dependent CREB activation is required for neuroprotection via NMDA receptor-PSD95 disruption [J]. Journal of Neurochemistry, 2013, 126 (2):274-287.
[15] Abel T, Nguyen P V, Barad M, et al. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory [J]. Cell, 1997, 88 (5): 615-626.
猜你喜欢
上海针灸杂志(2022年10期)2022-10-28
锦州医科大学报(2022年3期)2022-06-06
作文周刊·小学二年级版(2022年20期)2022-05-05
世界科学技术-中医药现代化(2021年8期)2021-12-21
中华养生保健(2020年4期)2020-11-16
中国医学影像技术(2020年8期)2020-09-25
中国实验诊断学(2020年1期)2020-02-07
创新作文(小学版)(2019年10期)2019-09-25
中国妇幼健康研究(2019年2期)2019-03-26
汽车观察(2009年1期)2009-02-18
