
杂交三倍体(4n♀×2n♂)泥鳅染色体组核型多态性研究
2016-12-02 02:10江振华周贺麻天宇沈芾林忠乔钱聪李雅娟
东北农业大学学报 2016年10期
江振华,周贺,麻天宇,沈芾,林忠乔,钱聪,李雅娟*
(1.大连海洋大学辽宁省海洋生物资源恢复及生境修复重点实验室,辽宁 大连 116023;2.北京市水产技术推广站,北京 101105)
杂交三倍体(4n♀×2n♂)泥鳅染色体组核型多态性研究
江振华1,周贺1,麻天宇1,沈芾2,林忠乔1,钱聪1,李雅娟1*
(1.大连海洋大学辽宁省海洋生物资源恢复及生境修复重点实验室,辽宁 大连 116023;2.北京市水产技术推广站,北京 101105)
以杂交三倍体泥鳅(4n♀×2n♂)为研究对象,初步分析对不同发育阶段(胚胎、六月龄及十二月龄)染色体核型。结果表明,胚胎阶段整三倍体(3n=75)染色体数目为3n=75,核型公式:14M+9SM+52T,NF=98;亚三倍体(3n>75)染色体数目为3n=73,核型公式:13M+6SM+54T,NF=92;超三倍体(3n>75)染色体数目为3n=77,核型公式:18M+7SM+52T,NF=102。六月龄阶段整三倍体染色体数目为3n=75,核型公式:15M+5SM+55T,NF=95;亚三倍体染色体数目为3n=73,核型公式:13M+6SM+54T,NF=92;超三倍体染色体数目为3n=76,核型公式:14M+6SM+56T,NF=96。十二月龄阶段整三倍体染色体数目为3n=75,核型公式:16M+6SM+53T,NF= 97;亚三倍体的染色体数目为3n=72,核型公式:13M+6SM+53T,NF=91;超三倍体的染色体数目为3n=77,核型公式:13M+6SM+58T,NF=96。研究表明,杂交三倍体(4n♀×2n♂)泥鳅不同发育阶段染色体核型整倍体及非整倍体均表现出多态性。结论为杂交三倍体泥鳅染色体组不稳定性研究提供细胞遗传学依据。
泥鳅;杂交三倍体;不同发育阶段;染色体核型
江振华,周贺,麻天宇,等.杂交三倍体(4n♀×2n♂)泥鳅染色体组核型多态性研究[J].东北农业大学学报,2016,47(10):34-40.
Jang Zhenhua,Zhou He,Ma Tianyu,et al.Resreach on karyotype polymorphism of hybrid triploid(4n♀×2n♂)loachMisgurnus anguillicaudatus[J].Journal of Northeast Agricultural University,2016,47(10):34-40.(in Chinese with English abstract)
水产动物中鱼类普遍存在多倍体现象,我国有二倍体、三倍体和四倍体3个种群[1-4]。印杰等报道我国长江流域的自然四倍体泥鳅营养价值高、生长速度快、耗氧率低、种质价值高[5]。
本文前期研究证实自然四倍体泥鳅是含有四套染色体组的遗传四倍体,无论雌雄均能产生2n配子。利用二倍体与四倍体泥鳅正、反杂交获得100%杂交三倍体。研究正、反交F1体细胞染色体数目表明,正、反杂交后代在早期胚胎、6月龄及12月龄等不同发育阶段存在大量非整倍体[6]。研究正交(2n♀×4n♂)F1体细胞染色体核型结果表明,染色体核型表现出多态性,非整倍体(超三倍和亚三倍)染色体数量及类型无规律[7]。关于反交(4n♀×2n♂)F1体细胞的染色体核型迄今尚不清楚。因此,本研究以长江流域天然四倍体泥鳅为母本,大连农贸市场购买的二倍体泥鳅为父本倍间杂交获得杂交三倍体泥鳅为研究对象,低渗—空气干燥法制备单个胚胎、6月龄及12月龄鳃细胞染色体标本,分析染色体核型,旨在了解反交组合(4n♀×2n♂)杂交三倍体泥鳅不同发育阶段染色体行为,为杂交三倍体泥鳅染色体组的不稳定性提供细胞学依据。
1 材料与方法
1.1 亲本采集
天然四倍体泥鳅采自长江流域湖北省赤壁市,二倍体泥鳅购于大连市农贸市场。于大连海洋大学细胞与工程实验室水族箱内暂养,温度为(22±1)℃。通过流式细胞仪(Partec PAS-Ⅲ,PARTEC,Münster,Germany)和血红细胞核体积测量方法鉴定倍性。
1.2 亲本的倍性鉴定
1.2.1 红细胞核体积测量
亲本泥鳅为测定对象。鱼体用苯甲醇稀释液(1 m·L-1)麻醉,在臀鳍上方抽取少量血液,迅速滴在玻璃片上。待血液风干后,滴1滴甲醇于载玻片,再用磷酸缓冲液(pH=6.8或7.4)10%吉姆萨染料染色5 min,自然风干后显微镜下拍照,测量细胞核长度。每尾鱼作血涂片1张,每张取50个细胞。分别测量红细胞和红细胞核长径(a)、短径(b),细胞核体积计算公式为V=4/3×π(a/2)(b/2)2,计算结果与前期试验结果对比,鉴定倍性。
1.2.2 流式细胞仪检测
已知二倍体泥鳅作为对照、亲本泥鳅为测定对象。鱼体用苯甲醇稀释液(1 m·L-1)麻醉,取1 mm2尾鳍组织,加入100 μL裂解液于振荡器上震荡。组织充分裂解30 min,期间每隔10 min震荡1次。向裂解完毕的组织中加入500 μL DAPI染料,震荡混匀后静置数秒。将所有液体经500目筛网过滤,获得细胞悬液使用Partec PAS-Ⅲ检测。
1.3 人工催产及授精
选取体表无伤,性腺发育良好的二倍体泥鳅雄鱼2尾与天然四倍体泥鳅雌鱼2尾,注射绒毛膜促性腺激素(HCG)(注射剂量:♀20~25 UI·尾-1,♂减半)人工催产,12 h后确认雌鱼排卵情况,轻压雌鱼腹部若有卵排出,即可用稀释1 000倍的苯甲醇将泥鳅麻醉,雄鱼生殖孔朝上置于湿润纱布,一手持毛细管,平放于生殖孔旁边,一手拇指从胸部以下开始向腹部挤压收集精液,置于碎冰上保持低温,胶头滴管吸1滴置于干燥的载玻片上,加入1滴曝气水激活,光学显微镜下观察精子活力。麻醉雌鱼生殖孔朝上置于湿纱布上,生理盐水冲洗生殖孔,使生殖孔朝下尽量弯曲成弧形,中间腹部最低,用拇指从胸部以下向腹部挤压,可见卵从生殖孔流出,把卵挤到事先用保鲜膜包裹的9 cm培养皿中,精子与卵混合,羽毛按顺时针方向搅匀,加入淡水激活,混匀。配制4n♀× 2n♂杂交组合。
1.4 受精卵、幼鱼培育
受精卵培育温度为(25±1)℃。每天早晚各换2次曝气水。保持室内空气新鲜和流通。受精后每2 h挑1次死卵,避免死卵污染水质影响其他受精卵发育。孵化后幼鱼1 d换水1次。孵化后第3天卵黄吸收完毕,开口后投饵。
1.5 染色体标本制备
1.5.1 单个胚胎染色体标本制备
取发育至眼胞期(受精后约18 h)胚胎30~50个,放入铺有1%琼脂糖培养皿中,加入适量生理盐水,解剖镜下用尖头镊子剥去卵膜和卵黄,胶头吸管去除卵膜将卵黄胚胎吸出,放入盛有0.0025%秋水仙素的青霉素小瓶中避光处理45 min后,吸出秋水仙素,换为0.8%的柠檬酸低渗20 min,吸出柠檬酸,预冷的卡诺固定液(甲醇∶冰醋酸=3∶1)固定3次,每次15~20 min,-20℃冰箱冷冻过夜。
将预冷的四凹槽载玻片放在冰上,取1个胚胎置于凹槽中,吸干周围液体,滴1滴50%冰醋酸,用黄枪头捣碎呈细胞悬液,再加1滴预冷的卡诺固定液,吸出所有液体,液体滴于提前预冷载玻片,在酒精灯上过火,可见载玻片上蓝色火焰,将载玻片倾斜放于托盘中,室温下自然风干。完全干燥的载玻片用磷酸缓冲液(pH=6.8或7.4)配置的10%吉姆萨染液染色45 min,之后用纯水冲洗,自然风干,染色体标本在普通光学显微镜下选择分散均匀、图像清晰的中期分裂相于10×倍物镜下观察,100×倍物镜(油镜)下拍照。
1.5.2 鳃组织细胞染色体标制备
采用修改后鱼类染色体标本快速制备方法制备6和12月龄的泥鳅取鳃组织染色体标本[8],首先腹腔注射植物血球凝集素(PHA)溶液,注射剂量为6 μg·g-1鱼体质量,作用时间为18~20 h,之后第2次注射(剂量同第1次);4~6 h后注射体积分数为0.1%的秋水仙素溶液,剂量为6 μg·g-1鱼体质量;2~3 h后,用稀释1 000倍的苯甲醇溶液将鱼麻醉后在臀鳍后2 mm处剪断尾巴,放血,取出全部鳃组织于淡水生理盐水中轻轻涮洗,去除血细胞,0.8%柠檬酸溶液中低渗40~45 min;用卡诺固定液(甲醇∶冰醋酸=9∶1)固定10 min,用100%冰醋酸处理2 min,预冷卡诺固定液(甲醇∶冰醋酸=3∶1)固定3次,每次15 min;-20℃冰箱冷冻贮藏过夜。样品取出置于5 mL小烧杯中,加入几滴50%冰醋酸,用小剪刀将其剪成细胞悬浊液,此步骤需在冰上完成,加入适量预冷卡诺固定液(3甲醇∶1冰醋酸)用过滤筛将细胞悬浊液过滤到另1个干净烧杯中。冷滴片、染色及观察方法同1.5.1。
1.6 核型分析
选染色体形态适中、分散良好、无重叠中期分裂相5个,测量长臂、短臂,计算染色体的相对长度和臂比。核型分析参照文献[9]。相对臂长=每条染色体长度/一组染色体总长度×100%,臂比=长臂长度/短臂长度,可按照着丝粒位置、臂比范围分为4类:臂比在1.00~1.70为中部着丝点染色体(M);臂比在1.71~3.00为亚中部着丝点染色体(SM);臂比在3.01~7.00为亚端部着丝点染色体(ST);臂比在>7.01为中部着丝点染色体(T);通过染色体核型分析,每个物种有各自的核型公式和臂数(NF),一般将M、SM染色体臂数记为2,将ST、T染色体的臂数计为1,在某些着丝粒中,除去着丝粒(主缢痕)外还有一段稍窄淡染区域称为次缢痕,某些染色体还有随体。
2 结果与分析
2.1 亲本倍性鉴定结果
2.1.1 血红细胞核体积测量
与其他鱼类一样,泥鳅红细胞核呈椭圆球体,具有致密椭圆形细胞核。与高泽霞等结果对比,确定亲本倍性为二倍体和四倍体(见图1A,1B)[10]。泥鳅正常二倍体DNA相对含量200为对照组,在100倍镜下观察泥鳅血涂片结果。图1A、1B分别为二倍体雄性和四倍体雌性亲本泥鳅红细胞照片。
2.1.2 流式细胞仪测定结果
流式细胞仪结果峰图见图1C,1D。分别为二倍体雄性和四倍体雌性亲本DNA含量峰图。
由图1可知,二倍体雄性亲本泥鳅鳍细胞DNA含量在200处出现第1个高峰;四倍体雌性亲本泥鳅的鳍细胞DNA含量在400处出现第1个高峰。
2.2 胚胎染色体核型分析
杂交三倍体泥鳅早期胚胎阶段整三倍体染色体数目为3n=75,核型公式:14M+9SM+52T,NF= 98(见图2A);亚三倍体的染色体数目为3n=73,核型公式:13M+6SM+54T,NF=92(见图2B);超三倍体的染色体数目为3n=77,核型公式:18M+ 7SM+52T,NF=102(见图2C)。
2.3 6月龄染色体核型分析
6月龄阶段的杂交三倍体泥鳅整三倍体染色体数目为3n=75,其中核型公式:15M+5SM+55T,NF=95(见图3A);亚三倍体染色体数目为3n=73,核型公式:13M+6SM+54T,NF=92(见图3B);超三倍体染色体数目为3n=76,核型公式:14M+6 SM+56T,NF=96(见图3C)。
2.412月龄染色体核型分析
12月龄阶段的杂交三倍体泥鳅整三倍体的染色体数目为3n=75,核型公式:16M+6SM+53T,NF=97(见图4A);亚三倍体的染色体数目为3n= 72,核型公式:13M+6SM+53T,NF=91(见图4B);超三倍体的染色体数目为3n=77,核型公式:13M+6SM+58T,NF=96(见图4C)。

图1 亲本倍性鉴定结果Fig.1Ploidy identification of parental

图2 四倍体泥鳅♀×二倍体泥鳅♂F1胚胎染色体中期分裂相及核型Fig.2The metaphase and chromosomes karyotype of hybrid triploid loach(4n♀×2n♂)at early embryonic stage

图3 四倍体泥鳅♀×二倍体泥鳅♂F16月龄鳃细胞染色体中期分裂相及核型Fig.3The metaphaseandchromosomeskaryotypeofsixmonthagegillcellofhybridtriploidloach(4n♀×2n♂)
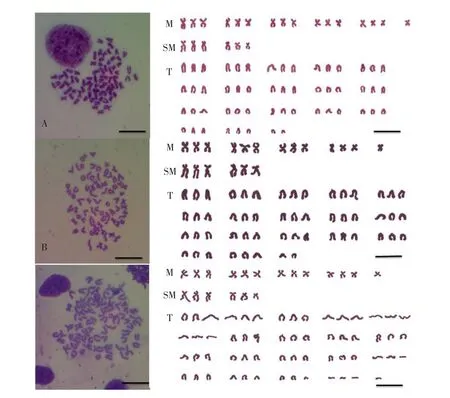
图4 四倍体泥鳅♀×二倍体泥鳅♂F112月龄鳃细胞染色体中期分裂相及核型Fig.4The metaphase and chromosomes karyotype oftwelvemonthagegill cell of hybrid triploidloach(4n♀×2n♂)
4 讨论与结论
染色体标本制备在细胞遗传学研究中具有重要意义[11]。染色体数目、染色体臂数、染色体排列配对方式及同亲缘种比较等作为判断物种倍性的鉴定指标。
染色体核型分析是根据染色体长度、着丝点位置、臂比、随体有无等特征,借助染色体分带技术,对某一生物体细胞分裂中期染色体分析、比较、排序和编号,是细胞遗传学研究基本方法。
李雅娟等采用染色体计数法,对杂交三倍体泥鳅(正交:2n♀×4n♂,反交:4n♀×2n♂)胚胎、6月龄、12月龄等不同发育阶段染色体数目统计分析,结果表明,不同发育阶段均出现大量存活的非整三倍体个体,且非整三倍体率随养殖时间延长逐渐下降[7]。与Zhang等报道超三倍体泥鳅能够存活结果一致[12]。在贝类中曾有关于非整倍体存活的报道,原因可能是贝类是低等生物可耐受非整倍体存在。迄今为止,在太平洋牡蛎中发现有12种可存活的非整倍体,生长正常,且在某些方面表现一定优势[13-14];在皱纹盘鲍和合浦珠母贝中也发现存活的非整倍体[15-16]。无论是鱼类还是贝类产生非整倍体的原因主要是在生殖细胞减数分裂过程中,前期Ⅰ的偶线期同源染色体不能配对、后期Ⅰ同源染色体不分离或分离延迟等,形成(n+1或n-1)的非整倍体配子。这些非整倍体配子与正常配子(n)结合时,形成各种各样的非整倍体[17]。
不同倍性个体间杂交产生非整倍体的原因是细胞遗传学研究热点。Guo等研究显示,太平洋牡蛎四倍体在减数分裂过程中前期Ⅰ同源染色体配对不完全,形成数目不等单价体和三价体,非整倍体形成[18]。Allen等研究表明,三倍体因含有3套染色体组,在减数分裂时有可能失去1套染色体组,造成染色体组的不稳定[19]。姜波等在太平洋牡蛎研究中认为二倍体×四倍体杂交时,非整倍体产生是因父本四倍体在减数分裂时出现错误,形成三价体或单价体精子[20]。Li等研究发现,我国天然四倍体泥鳅无论雌、雄在减数分裂时均产生0~6个四价染色体和38~50个二价染色体[21]。因此,本研究采用四倍体雌性泥鳅与二倍体雄性泥鳅杂交产生非整倍体,原因可能是四倍体母本在减数分裂过程中,由于四价体,造成后期Ⅰ同源染色体不均等分离,产生非整倍体配子。这与姜波等研究的结果一致[20]。本研究分析杂交三倍体(4n♀×2n♂)泥鳅不同发育阶段的染色体核型,结果表明,杂交三倍体泥鳅在胚胎、6月龄及12月龄存在大量非整倍体,不同发育阶段的整三倍体、亚三倍体和超三倍体染色体数目虽相同,但核型公式不同,即染色体核型表现多态性。非整倍体中缺失、重复染色体数目及类型无规律。
本研究结果表明,以四倍体为母本,二倍体为父本产生的杂交三倍体子代不同发育阶段染色体核型表现多态性。说明自然四倍体泥鳅作为母本时产生非整倍体雌配子。为杂交三倍体泥鳅染色体组不稳定性研究提供细胞遗传学依据。
[1] 李康,李渝成,周暾.两种泥鳅染色体组型的比较研究[J].动物学研究,1983,4(1):75-80.
[2] 李渝成,李康.马口鱼和泥鳅的核型研究兼论鱼类染色体数目多态与分类的关系[J].武汉大学学报,1987(1):107-112.
[3] 李雅娟,印傑,王嘉博,等.中国29の地点ドジョウにおける倍数体の分布に関する研究[J].日本水産学会誌,2008,74(2): 177-182.
[4] 李雅娟,隋燚,赵睿,李霞,等.天然二倍体和四倍体泥鳅鳍细胞系染色体组构成研究[J].东北农业大学学报,2015,46(4): 83-88.
[5] 印杰,赵振山,陈小奇,等.二倍体和四倍体泥鳅染色体组型比较[J].水生生物报,2005,4(29):469-472.
[6] 周贺,马海艳,赵睿,等.二倍体泥鳅×四倍体泥鳅F1不同发育阶段染色体核型分析[J].东北农业大学学报,2015,46(7):70-75.
[7] 李雅娟,钱聪,印杰,等.不同倍性泥鳅杂交后代染色体数目组成研究[J].大连海洋大学学报,2012,27(4):326-332.
[8] 张四明.一种制备鱼类染色体的新方法[J].遗传,1993,15(3): 35-36.
[9] Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52(2):201-220.
[10] 高泽霞,王卫民,周小云.2种鉴定泥鳅多倍体方法的比较[J].华中农业大学学报,2007,26(4):524-527.
[11] 楼允东.中国鱼类染色体组型研究的进展[J].水产学报,1997 (1):82-96.
[12] Zhang Q,Arai K.Aberrant meioses and viable aneuploid progeny of induced triploid loach(Misgurnsus anguillicaudatus)when crossed to natural tetraploids[J].Aquaculture.1999,175:63-76.
[13] Wang Z,Guo X,Allen S K,et al.Aneuploid Pacific oyster (Crassostrea gigas Thunberg)as incidentals from triploid produc⁃tion[J].Aquaeulture,1999,173:347-357.
[14] Guo X,Zhang G,Landau B J,et al.Aneuploidy in the Pacific oys⁃ter Crassostrea gigas Thunberg)and its effects on growth[J].Jour⁃nal of shellfish Research,2000,19(1):614-619.
[15] 何毛贤,林岳光,沈琪,等.合浦珠母贝四倍体诱导过程中非整倍体的产生[J].热带海洋,2000,19(4):59-64.
[16] Fujino K,Arai K,Iwadare K et al.Induction of gynogenetic dip⁃loid by inhibiting second meiosis in the Pacific abalone[J].Nip⁃pon Suisan Gakkaishi,1990,56(11):1755-1763.
[17] 王伟伟.谈染色体非整倍体变异[J].中学生物学,2003(4):15-16.
[18] Guo X,Jr A S K.Sex and meiosis in autotetraploid Pacific oyster (Crassostrea gigas Thunberg)[J].Genome,1997,40:397-405.
[19] Jr A S K.Guo X,Burreson G et al.Heteroploid mosaics and rever⁃sion among triploid oysters,Crassostrea gigas:fact or artifact.[J]. Shellfish.Res.,1996,15(2):514-522.
[20] 姜波,王昭萍,于瑞海,等.杂交三倍体太平洋牡蛎群体的染色体数目组成初步观察[J].中国海洋大学学报,2007,37(2):255-258.
[21] Li Y J,Yu Z,Zhang M Z,et al.The origin of natural tetraploid loach Misgurnus anguillicaudatus(Teleostei:Cobitidae)inferred from meiotic chromosome configurations[J].Genetica,2011,139 (6):805-811.
Resreach on karyotype polymorphism of hybrid triploid(4n♀×2n♂) loachMisgurnus anguillicaudatus
JANG Zhenhua1,ZHOU He1,MA Tianyu1,SHEN Fu2,
LIN Zhongqiao1,QIAN Cong1,LI Yajuan1(1.China Key Laboratory of Marine Bio-resources Restoration and Habitat Reparation in Liaoning Province,Dalian Ocean University,Dalian Liaoning 116023,China; 2.Fisheries Technology Extension Station of Beijing,Beijing 101105,China)
This study aimed at analysis chromosome karyotype of hybird triploid loach(tetraploid♀× diploid♂)in different development stages(early embryo,six months old and twelve months old).The results of early embryos loach showed that chromosomes number of triploid(3n=75)was 3n=75,karyotype formula was 14M+9SM+52TandNF=98;chromosomes number of hypotriploid(3n<75)was 3n=73,karyotype formula was 13M+6SM+54T,andNFwas 92;the chromosomes number of hypertriploid(3n>75)was 3n=77, karyotype formula was 18M+7SM+52T,andNFwas 102.The results of six months old loach showed that the chromosomes number of triploid was 3n=75,karyotype formula was 15M+5SM+55T,andNFwas 95;the chromosomes number of hypotriploid was 3n=73,karyotype formula was 13M+6SM+54T,andNFwas 92; the chromosomes number of hypertriploid was 3n=76,karyotype formula was 14M+6SM+56T,andNFwas
Misgurnus anguillicaudatus;hybird triploid;different developmental stages;chromosome karyotype
S917.4
A
1005-9369(2016)10-0034-07
时间2016-10-26 16:38:00[URL]http://www.cnki.net/kcms/detail/23.1391.S.20161026.1638.020.html
2016-07-13
国家自然科学基金项目(31272650)
江振华(1991-),男,硕士研究生,研究方向为水产动物遗传育种。E-mail:jzh9103@163.com
*通讯作者:李雅娟,教授,博士,研究方向为水产动物遗传育种。E-mail:liyajuan@dlou.edu.cn
96.The results of twelve months old loach showed that the chromosomes number of triploid was 3n=75, karyotype formula was 16M+6SM+53T,andNFwas 97;the chromosomes number of hypotriploid was 3n= 72,karyotype formula was 13M+6SM+53T,andNFwas 91;the chromosomes number of hypertriploid was 3n=77,karyotype formula was 13M+6SM+58T,andNFwas 96.The study showed that chromosome karyotype both of euploid and aneuploid showed polymorphism of hybird triploid loach in the different development stages.The conclusion provided a cytogenetic basis for hybrid triploid loach chromosome instability.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
科学养鱼(2021年11期)2022-01-11
热带农业科技(2020年3期)2020-12-15
广东医科大学学报(2020年6期)2020-02-06
中国妇幼健康研究(2019年12期)2019-12-31
食品与生活(2019年12期)2019-05-15
中华肺部疾病杂志(电子版)(2018年1期)2018-03-12
现代园艺(2017年13期)2018-01-19
中成药(2016年4期)2016-05-17
中国卫生标准管理(2015年4期)2016-01-14
