
不同磷营养条件下米槁光响应曲线模型比较分析
2016-12-02 02:10骆畅刘济明王军才李丽霞熊雪柳嘉佳
东北农业大学学报 2016年10期
骆畅,刘济明,王军才,李丽霞,熊雪,柳嘉佳
(贵州大学林学院,贵阳 550025)
不同磷营养条件下米槁光响应曲线模型比较分析
骆畅,刘济明*,王军才,李丽霞,熊雪,柳嘉佳
(贵州大学林学院,贵阳 550025)
以半年生米槁幼苗为试验材料,温室条件下砂培,设置四种不同磷素浓度供给,分别测定不同处理下米槁幼苗光合特征,并利用六种模型拟合米槁叶片光响应曲线,筛选最佳光响应模型。结果表明,直角双曲线修正模型是拟合米槁光响应最优模型。磷素供给与米槁幼苗光合特性相关,在2 mmol·L-1磷浓度处理下,米槁光饱和点(LSP)、最大净光合速率(Pnmax)、表观量子率(AQE)均大于其他处理,光补偿点(LCP)小于其他处理。米槁叶片气孔导度(Gs)、蒸腾速率(Tr)随施磷量增加呈增大趋势,胞间CO2浓度(Ci)呈下降趋势。在一定程度上,增施磷素可改变米槁对光的适应度,提高叶片光合能力。
米槁;磷;光响应模型
骆畅,刘济明,王军才,等.不同磷营养条件下米槁光响应曲线模型比较分析[J].东北农业大学学报,2016,47(10):17-24.
Luo Chang,Liu Jiming,Wang Juncai,et al.Comparison and analysis of fitting models of light response curve ofCinnamomum migaounder different phosphorus levels[J].Journal of Northeast Agricultural University,2016,47(10):17-24.(in Chinese with English abstract)
光合作用是植物吸收光能,同化CO2和水,制造有机物质并释放氧气的生理生化过程,受环境因子和自身生理特征共同影响[1]。光响应曲线测定及其模型拟合可估算植物光合作用表观量子效率(AQE)、最大净光合速率(Pnmax)、初始量子效率(α)、光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率等光合生理参数(Rd),对研究不同生理过程和环境下,植物光合能力及光适应性、光能利用效率等具有重要意义[2]。Bassman建立模型研究植物光响应曲线,包括指数模型[3]、直角双曲线模型[4]、非直角双曲线模型[5]、二次项模型[6]、动力学模型[7]、直角双曲线修正模型等[8-9]。
磷在植物生长代谢过程中具有重要作用。地壳中磷含量丰富,但植物可利用的有效磷(包括H2PO4-和HPO42-)非常少,即使施加磷肥,也会因土壤固定作用而无法利用[10-11]。磷酸盐在土壤中缓慢扩散[12],植物常处于缺磷环境中。磷素对植物光合作用具有重要影响,植物缺磷净光合速率、最大荧光、电子传递速率、PSⅡ光化学效率等降低,叶片光合作用光抑制加重[13]。缺磷还会影响光合作用中同化力形成、光合机构及同化物运输[14-15]。米槁(Cinnamomum migao H.W.Li)是樟科(Lauraceae)樟属常绿乔木,其果实大果木姜子是近年重点发展的民族特色药材之一[16]。目前,关于磷素对落叶松[17-18]等植物光合影响均有研究,但对米槁光响应影响研究鲜有报道。为了解米槁光合特征,采用六种模型拟合不同磷素浓度下米槁幼苗光响应曲线,比较模型间差异。同时,分析施磷量对相关光合参数影响,为研究米槁磷素营养生理和栽培管理提供理论依据。
1 材料与方法
1.1 材料与设计
本试验所用营养液参照霍格兰营养液及Jarkko等营养液配方配制[19],营养液配方如下:NH4NO38 mmol·L-1;KH2PO41 mmol·L-1;KCl 1 mmol·L-1;CaCl2·6H2O 1 mmol·L-1;MgSO4·7H2O 0.6 mmol·L-1;FeCl3·6H2O 0.02 mmol·L-1;MnCl2· 4H2O 6 μmol·L-1;H3BO30.016 mmol·L-1;ZnCl20.3 μmol·L-1;CuCl2·2H2O 0.3 μmol·L-1;NaMoO4·2H2O 0.3 μmol·L-1。通过调节KH2PO4浓度形成4个磷养分梯度,0.125 mmol·L-1(P1)、0.5 mmol·L-1(P2)、1 mmol·L-1(CK)、2 mmol·L-1(P3),KCl调节K浓度,CaCl2调节离子平衡,其他成分相同。
1.2 试验方法
2015年9月7 ~25日选取晴天,于上午采用LI-6400XT光合作用测定系统测定各处理植株叶片光响应曲线。每个处理选取长势一致健康植株3株,以中部叶位叶片作为测定对象。测定时,采用开放气路,控制叶室温度25℃,CO2浓度400 μmol·mol-1,气体流速500 μmol·s-1,用02B-LED红蓝光源,设定光合有效辐射(PAR)梯度为:2 200、2 000、1 800、1 500、1 200、1 000、800、500、300、200、100、80、50、20、0 μmol·m-2·s-1。仪器自动记录净光合速率Pn、气孔导度Gs、胞间CO2浓度Ci、蒸腾速率Tr等生理参数。重复3次,取平均值。采用96%乙醇提取、分光光度法[20]测定叶绿素含量。
1.3 数据分析
测定数据在Excel 2013软件中初步处理,以统计软件SPSS 22.0中的非线性回归功能拟合数据,以非线性最小二乘法的Marquardt迭代原理求解模型参数。
2 结果与分析
2.1 光响应曲线模型参数拟合效果分析
由图1A可知,Pn随PAR增大而增大,无下降趋势,不考虑曲线凸度,直角双曲线模型是无极值函数,导致该模型无法拟合植物出现光抑制现象的光响应曲线。由图1B可知,非直角双曲线模型和直角双曲线模型一样,无法拟合实测值下降趋势。由图1D可知,指数模型拟合值在PAR为200~800 μmol·m-2·s-1时,各处理拟合值大于实测值,拟合光响应曲线时未考虑曲线弯曲程度,
拟合值偏离实测值。
古卷的名字,叫做《天下图》,起先用一幅地图,粗简地勾勒出中州各地的行政区划。从中,他得知了中州之下共分九州,地域广阔,山高水长。而自己所在的云浮山,就位于中州西侧的横断山脉中,向东翻越数道山脉,便是中州治下的巴州地界。
由表1可知,以上三种模型得出光合参数Pnmax、LSP和Rd均与实测值相差较大,拟合决定系数R2相对较高,仅说明模型与数据间拟合程度较高,无法说明拟合结果一定与实际相符[21]。由图1E可知,二次多项式模型拟合值与实测值相差较大,且得到的LCP、Rd为负值(见表1),整体拟合效果差,无法反映各处理光响应特征。动力学模型和直角双曲线模型在不同光强下拟合图完全一致(见图1B),由动力学模型拟合得到的Rd为负值(见表1),与生物学常识不符。
综合对比不同模型给出的各磷素处理光合参数(见表1),仅修正模型得出光合参数与实测值接近,更符合米槁实际生理情况。同时,由图1C可知,修正模型能拟合米槁叶片光抑制现象。综上所述,各模型优劣为修正模型>非直角模型>指数模型>直角模型>动力学模型>二次多项式模型。分析不同氮磷素处理下米槁幼苗各光合参数采用修正模型的拟合值。

A-直角双曲线模型(动力学模型)A-Right angle hyperbolic model(dynamic model)
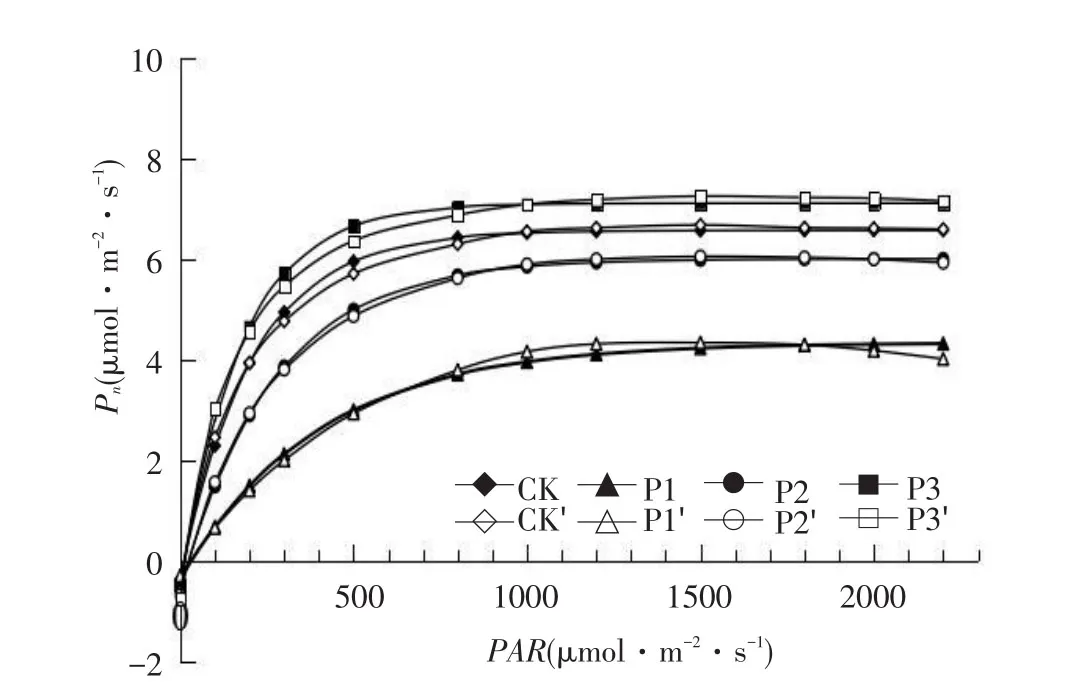
B-非直角双曲线模型B-Non rectangular hyperbolic model

C-直角双曲线修正模型C-Right angle hyperbolic correction model

D-指数模型D-Exponent model

图1 不同模型下拟合的光响应曲线Fig.1Light response curve of Cinnamomum migao fittedby the rectangular hyperbola modified model
The superscript sign'is measured value,the same as below.

表1 各磷素处理米槁光响应模型拟合参数值与实测值Table 1Light response model fitting parameters and measured values one of phosphorus the treatments for Cinnamomum migao
2.1.1 光饱和点和光补偿点
P1处理下LSP最小(1 483.56 μmol·m-2·s-1),LCP最大(29.50 μmol·m-2·s-1),而P3处理下LSP最大(1 600.417 μmol·m-2·s-1),LCP最小(13.638 μmol·m-2·s-1),即随施磷量增加,米槁幼苗LSP逐渐增大,LCP逐渐减小。植物叶片LCP和LSP体现植物光合能力,反映植物对光照适应性,LCP反映对弱光适应性,LSP反映对强光适应性,LCP低、LSP高的植物对光照适应性强[22]。试验表明增施磷素显著增强米槁幼苗对光适应幅度,尤其是对强光辐射的吸收利用率。
2.1.2 表观量子效率
表观量子效率AQE是弱光阶段的光响应曲线斜率,反映植物对弱光利用效率。由表1可知,米槁幼苗AQE随施磷量增加逐渐增大,但值均小于0.03,表明米槁耐阴能力差,增施磷素可提高米槁利用弱光能力。
2.1.3 最大净光合速率和暗呼吸速率
最大净光合速率是衡量叶片光合潜力重要指标,反映植物对强光利用能力,缺磷或少磷使植物净光合速率降低[23],随施磷量增加,米槁幼苗最大净光合速率递增,表明增施磷素可提高米槁幼苗叶片光合潜力。暗呼吸速率可反映植物消耗光合产物速率,主要受植物生长状态和温度影响。本试验中米槁幼苗叶片Rd对磷素响应总体呈上升趋势。
2.2 光合参数响应
2.2.1 气孔导度和胞间CO2浓度
气孔是植物叶片与外界气体和水分交换重要通道,光强直接影响气孔开度,气孔开度及速率随光强增加而增大[24]。随PAR增加,米槁净光合速率(图1)和对空气中CO2需求量不断增大。因此,米槁幼苗叶片气孔导度也逐渐增大(见图2),但P3、P2与CK间Gs相差不大,表明增施磷素一定程度上可提高米槁叶片气孔导度,增强米槁叶片对CO2同化能力。
胞间CO2浓度会影响光合作用暗反应阶段羧化速率,受气孔导度、外界CO2浓度及叶片光合消耗等因素影响。随PAR增加,各处理叶片Ci均呈先急速下降再趋于稳定后缓慢升高变化过程。在PAR为0~200 μmol·m-2·s-1时,因净光合速率迅速增加,消耗CO2量增加,叶片Ci急速下降,气孔导度较小(见图2)。在200<PAR<1 800 μmol·m-2·s-1时,随光强增大,气孔开放程度加大(见图3),外界CO2进入细胞间隙阻力减少,光合消耗CO2量与CO2进入量达到动态平衡,此时Ci总体保持稳定。在PAR>1 800 μmol·m-2·s-1时,出现光抑制现象,光呼吸增强,CO2释放量增加,而Pn略有下降(见图1C),消耗CO2量减少,造成外界CO2进入细胞间隙速度减小,Ci缓慢增加。在同一光强下,Ci均表现为:P3<CK<P2<P1。与Pn、Gs呈负相关,说明增施磷素可减少米槁叶片Ci值。

图2 气孔导度的光响应Fig.2Light responses of Gs

图3 胞间CO2浓度的光响应Fig.3Light responses of Ci
2.2.2 蒸腾速率
由图4可知,随PAR增大,不同处理下米槁幼苗叶片蒸腾速率(Tr)均增大。各处理叶片蒸腾速率先随PAR增加迅速增加,而后增大趋势减缓,最后趋于稳定,当PAR>1 800 μmol·m-2·s-1时,各处理Tr缓慢下降,与Gs变化相似(见图2)。分
析认为Tr下降主要是Gs下降导致气孔阻力增大所致。增施磷素可提高米槁叶片Tr,但P3与CK差异不显著,增施磷素可改变米槁叶片蒸腾速率但作用不明显。
2.2.3 叶绿素
叶绿素是植物光合作用重要介质,其含量可衡量植物光合作用能力。
不同施磷处理对米槁幼苗叶片中叶绿素含量有一定影响,P3组叶绿素含量较高,说明高磷可促进米槁叶片叶绿素合成,但在同一时间下,各处理间差异不大(见表2),说明磷素影响程度较小。

图4 蒸腾速率的光响应Fig.4Light responses ofTr

表2 不同处理下米槁幼苗叶片叶绿素含量Table 2Chlorophyll content in the leaves of Cinnamomum migao under different treatments (mg·g-1FW)
3 讨论与结论
不同植物最适光合作用光响应曲线模型不同,综合比较拟合光合参数与实测值差异,优选最适米槁的光响应曲线模型。直角双曲线模型和非直角双曲线模型的Pnmax均高于实测值,且无法拟合植物光抑制现象,由于这两种模型均是无极值渐近线,植物光合速率在饱和光强后未下降,无法直接估算植物饱和光强,得到的最大净光合速率远大于实测值,也无法解释光抑制现象[25-26]。在桉树无性系(Eucalyptus)[27]研究中也得到相同结果。指数模型中一部分拟合值大于实测值,指数模型光响应曲线未考虑曲线弯曲程度,也无法拟合饱和光强之后随光强增加光合速率降低这一段光响应曲线[28],二次多项式模型得到的LCP、Rd为负值,由动力学模型拟合得到Rd也为负值,与生物学常识不符。直角双曲线修正模型弥补了其他几种模型未考虑曲线凹度缺陷,可拟合植物在饱和光强后,随光强增加光合速率下降光响应曲线,且其估算出LSP和Pnmax与实测值最为接近[7,29]。直角双曲线修正模型较好拟合米槁在不同磷素处理下光响应曲线,得到光合参数与实测值相差小,该结果已在黄山栾树(Koelreuteria bipinnata var.Integrifoli⁃ola)[30]、欧美杨107(Populus×euramericana cv'.Ne⁃va')[31]等植物光合研究中证实。因此,可将修正模型作为研究米槁光响应曲线首选模型。
光合产物运输离不开磷,当植株培养在缺磷或少磷溶液中,叶片光合速率明显降低[32],缺磷时气孔导度下降[33]。根据试验修正模型拟合结果分析,P3处理下,Pnmax最大,而P1处理下Pnmax、气孔导度和Ci最小。缺磷导致净光合速率下降主要是由于磷不足影响代谢过程,磷素缺乏导致米槁利用光能能力降低,磷元素作为光合作用中ATP的重要原料,缺乏则可能对ATP再生产生影响,导致光合能力下降。P3处理中,米槁叶片在不同光强下Pnmax和α最大,气孔导度、蒸腾速率、净光合速率均最高。说明高磷有利于米槁光合作用合成更多
有机物。随施磷量增加,各处理米槁LSP、AQE、α、Pnmax均提高,LCP下降。植物LCP和LSP分别体现其对弱光和强光适应能力,反映其光照条件需求范围[34],LSP高,光合速率不容易达到饱点,光合作用在较低水平上进行,典型阳生植物LCP为9~27 μmol[35],说明米槁是阳生植物,施磷显著增加米槁对光适应幅度。而AQE表现植物对弱光利用能力,反映植物吸收与转化光能色素蛋白质复合体多寡及利用弱光能力强弱,间接反映Rubisco羧化酶活性[36]。综上所述,增加施磷量能改变米槁对光适应幅度,增强对光环境适应力,提高米槁叶片光合能力和光能利用率。本研究中供磷对叶绿素含量无显著影响,但随磷浓度增加,叶绿素含量仍有一定增加,与落叶松研究结果相同[18]。施肥对米槁幼苗生长长期调控及幼苗对其长期适应机制尚需进一步研究。
[1] Robert E S,Mark A,John S B.Kok effect and the quantum yield of photosynthesis[J].Plant Physiology,1984,75(1):95-101.
[2] Gardiner E S,Krauss K W.Photosynthetic light response of flood⁃ed cherry bark oak(Quercus pagoda)seedlings grown in twolight regimes[J].Tree Physiology,2001,21(15):1103-1111.
[3] Bassman J,Zwier J C.Gas exchange characteristics of Populus trichocarpa,Populus deltoids and Populus trichocarpa×P.del⁃toids clone[J].Tree Physiology,1991,8(2):145-159.
[4] Baly E C.The kinetics of photosynthesis[J].Proceedings of the Royal Society Series B(Biological Sciences).1935,117(104):218-239.
[5] Thornley J H M.Mathematical models in plant physiology[M]. London:Academic Press,1976,86-110.
[6] 刘锦春,钟章成,何跃军,等.重庆石灰岩地区十大功劳(Maho⁃nia fortunei)的光合响应研究[J].西南师范大学学报:自然科学版,2005,30(2):316-320.
[7] Broadley M R,Escobar-Gutierrez A J,Burns A,et al.Nitrogenlimited growth of lettuce is associated with lower stomatal conduc⁃tance[J].New Physiologist,2001,151(1):97-106.
[8] Ye Z P.A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa[J].Photosynthetica, 2007,45:637-640.
[9] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,
[10] Barrow N J.Evaluation and utilization of residual phosphorus in soils[J].Role of Phosphorus in Agriculature,1980,333-355.
[11] Marschner H.Mineral nutrition of higher plants[J].London:Aca⁃demic Press,1995:345-364
[12] Smith F W,Mudge S R,Rae A L,et al.Phosphate transport in plants[J].Plant Soil,2003,248:71-83
[13] 于海秋,彭新湘,曹敏建.缺磷对不同磷效率基因型大豆光合日变化的影响[J].沈阳农业大学学报,2005,36(5):519-522.
[14] 樊卫国,王立新.纽荷尔脐橙幼树对不同供磷水平的光合响应[J].果树学报,2012,29:166-170.
[15] 袁军.油茶低磷适应机理研究[D].北京:北京林业大学,2013.
[16] 邱德文,李鸿玉,赵山,等.米槁的本草学研究[J].中国医药学报,1993,8(2):19-21.
[17] 郭盛磊.氮、磷营养供应对落叶松幼苗生长和光合作用的影响[D].哈尔滨:东北林业大学.2004
[18] 吴楚,王政权,孙海龙.氮磷供给对长白落叶松叶绿素合成、叶绿素荧光和光合速率的影响[J].林业科学,2005,41(4):31-36.
[19] Jarkko U,Toini H.Influence of nitrogen and phosphorus availabil⁃ity and ozone stress on Norway spruce seedlings[J].Tree Physi⁃ology,2001,21:447-456.
[20] 张立军,樊金娟.植物生理学实验教程[M].北京:中国农业出版社,2007:36-39.
[21] Herrick J D,Thomas R B.Effects of CO2enrichment on the photo⁃synthetic light response of sun and shade leaves of canopy sweet⁃gum trees(Liquidambar styraciflua)in a forest ecosystem[J].Tree Physiology,1999,19:779-786.
[22] 刘玲,刘淑明,孙丙寅.不同产地花椒幼苗光合特性研究[J].西北农业学报,2009,18(3):160-165.
[23] Ciereszko I,Janonis A,Kociakowska M.Growth and metabolism of cucumber in phosphate-deficient conditions[J].Journal of Plant Nutrition,2002,25:1115-1127.
[24] 许大全.气孔运动与光合作用[J].植物生理学通讯,1984(6):6-12.
[25] Yu Q,Zhang Y Q,Liu Y F,et al.Simulation of the stomatal con⁃ductance of winter wheat in response to light,temperature and CO2changes[J].Annals of Botany,2004,93,435-441.
[26] Posada J M,Lechowicz M J,Kitajima K.Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of indi⁃vidual leaf angles and nitrogen content[J].Annals of Botany, 2009,103:795-805.
[27] 侯智勇,洪伟,李键,等.不同桉树无性系光响应曲线研究[J].福建林学院学报,2009,29(2):97-102.
[28] Prado C H B A,Moraes J A P V.Photosynthetic capacity and spe⁃cific leaf mass in twenty woody species of Cerrado vegetation un⁃der field condition[J].Photosynthetica,1997,33(1):103-112.
[29] 叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
[30] 叶子飘,康华靖,陶月良,等.不同模型对黄山栾树快速光曲线拟合效果的比较[J].生态学杂志,2011,30(8):1662-1667.
[31] 马永春,方升佐.欧美杨107不同冠层光合特性的研究[J].南京林业大学学报:自然科学版,2011,35(4):39-42.
[32] Sicher R C,Kremer D F.Effects of Phosphate deficiency on as⁃ similate partitioning in barley seedlings[J].Plant Science,1988; 57:9-17
[33] 张翠萍,孟平,李建中,等.磷元素和土壤酸化交互作用对核桃幼苗光合特性的影响[J].植物生态学报,2014,38(12):1345-1355.
[34] 黄成林,赵昌恒,傅松玲,等.安徽休宁倭竹光合生理特性的研究[J].安徽农业大学学报,2005,32(2):187-191
[35] 张振文,张保玉,童海峰,等.葡萄开花期光合作用光补偿点和光饱和点的研究[J].西北林学院学报,2010,25(1):24-29.
[36] 靳甜甜,刘国华,胡婵娟,等.黄土高原常见造林树种光合蒸腾特征[J].生态学报,2008,28(11):5758-5765.
Comparison and analysis of fitting models of light response curve of Cinnamomum migaounder different phosphorus levels
LUO Chang,LIU Jiming,WANG Juncai,LI Lixia,XIONG Xue,LIU Jiajia(School of Forestry,Guizhou University,Guiyang 550025,China)
The experiment was conducted in green house,the half-year-oldCinnamomum migao seedlings were cultured in sand and supplied with different phosphorus levels.The photosynthesis characteristics ofCinnamomum migaoseedlings on the condition of different levels of phosphorus supply were measured,and using six models to fit the light response curve under different phosphorus treatments ofCinnamomum migaoleaves.The optimal model was screened out from these models.The results showed that the rectangular hyperbola modified model was the optimal model that fittedCinnamomum migaolight response response curve.Different levels of phosphorus supplied and photosynthetic characteristics of seedlings correlated.Which under the concentration of phosphorus 2 mmol·L-1treatment,the light saturation point,the maximum net photosynthetic rate,apparent quantum rate were higher than other treatments,light compensation point was less than other treatments.With the increasing of phosphorus fertilizer,stomatal conductance and transpiration rate increased,intercellular CO2concentration decreased.Thus,increasing the amount of phosphorus fertilizer could changeCinnamomum migaoto light adaptation scope,improve Cinnamomum migaoleaf energy utilization rate.
Cinnamomum migao;phosphorus;light response model
Q945;S184
A
1005-9369(2016)10-0017-08
时间2016-10-26 16:38:00[URL]http://www.cnki.net/kcms/detail/23.1391.S.20161026.1638.016.html
2016-06-22
贵州省林业厅重大项目“喀斯特地区特色经济林树种培育与示范”(黔林科合[2010]重大04号);贵州省科技计划“大果木姜子生物生态学特征及育苗技术”(黔科合SY[2015]3023-1)
骆畅(1991-),女,硕士研究生,研究方向为野生植物资源保护与利用。E-mail:luochang_96@163.com
*通讯作者:刘济明,教授,博士生导师。研究方向为植物生态学。E-mail:karst0623@163.com。
猜你喜欢
农业工程学报(2022年5期)2022-06-22
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
中学生数理化·七年级数学人教版(2017年9期)2017-08-15
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
土壤与作物(2013年4期)2013-03-11
植物营养与肥料学报(2012年3期)2012-10-26
