
竞技能力增长理论模型及其演进
2016-12-15 01:47胡海旭
体育科学 2016年2期
胡海旭
竞技能力增长理论模型及其演进
胡海旭
主要运用历史研究方法和文献资料法,对20世纪中后期竞技能力增长的理论模型及其演进特征进行揭示和剖析,厘清其理论的科学基础与演进特点。结果表明:1)竞技能力增长模型大抵进路为:以1950年代Selye应激理论提出的GAS模型衍生而来的运动训练“超量补偿循环”模型、“高级适应循环”模型、“超量补偿循环和代偿适应”模型等生理学理论模型,到基于生理学、物理学和数学、计算机科学提出的 CP模型、IR模型、PerPot模型等计算模型;2)生理学是运动训练的重要基础;3)数学模型通过假设与验证,实现对复杂生物系统的有效表达,并可根据个体参数不同实现模型个性化;4)融合计算机信息技术的“PerPot模型”则通过现有数据,即刻优化训练计划和预防过度训练,借助其强大的运算功能,达到运动训练数据与目标成绩之间的高度拟合。研究认为,同其他科学与社会的发展规律一致,如吉姆·格雷在“科学的第4个范式”中所言,数据密集型科学理论或同样是运动训练理论与方法的下一个科学范式,并有待基于此范式,构建下一个竞技能力增长理论模型。
运动训练;竞技能力增长;生理学;数学;计算科学;数据密集型
自从运动训练实践诞生以来,人们一直都试图寻找提高运动训练效果的机制原理来揭示运动训练过程,以便更好地指导运动训练活动。值得注意的是,在经历两次世界大战后,全世界对运动训练的关注似乎更加活跃。战后的世界,面临急剧矛盾冲突后的内部结构调整,同样,压制已久的科学创造成果与社会改革力量迎来释放的时机,科学社会的全面繁荣为体育科学,尤其是为携带国家政治基因的竞技体育运动训练活动注入了全新的动力。事实也表明,不到20年的时间,运动训练相关的现代基础理论全面爆发,如运动生理学、运动心理学、运动生物力学等学科的独立发展均诞生于1960s[54]。而1980s“大科学时代”的来临,同样也带来运动训练科学化的发展。人们愈发意识到运动训练理论已不再是单一学科理论可以应对的。一方面,受到国际大环境趋于和平的诉求,各国将竞技体育视作国际交流的重要舞台;另一方面,此时来之不易的和平为科学发展铺设了肥沃的土壤,推动科学的迅猛发展以及科学对和平守护的多重张力,共同催生了科学创新和学科发展的勃兴。由此,也影响着运动训练的科学化进程。基于此,本文试图对这一时期以来指导运动训练的理论模型进行梳理和解析,拟将这类模型归类为竞技能力增长理论模型,厘清人们认识和解释运动训练活动的科学进路,为推动当前运动训练理论与方法的发展提供一些历史逻辑与科学启示,为运动训练实践提供一些理论参考。
1 生理模型及其演进
20世纪50年代,加拿大籍匈牙利内分泌学家塞里(H·Selye,1907—1982)[36]基于伯尔纳(Claude Bernard)的“内部介质”和坎农(W.B.Cannon)的“内稳态”学说,在1936—1956年期间提出生物的生理“应激理论”。塞里把应激看作是人或动物有机体对环境刺激的一种生物学反应现象,由加在机体上的许多不同需求引起,并且是周身的,非特异性的,统称为一般适应综合症(the general adaptation syndrome,GAS),表现为报警、抵抗、衰竭3个阶段。塞里提出的非特异性,是指多样性、性质不同的刺激引起同样的或类似的全身反应,如果刺激太强,时间太长,机体会逐渐失去应付能力,则出现衰竭,陷入病理状态。随着应激的普遍性及其研究的深入,当前已经发展到如下几种情形[7]:1)应激是指那些使人感到紧张的事件或刺激环境,从这个意义上说,应激对人是外部的;2)应激是指一种主观反映,从这个意义上讲,应激是紧张或唤醒的一种内部心理状态,是人体内部出现的解释性的、情感性的、防御性的应对过程;3)应激是指人体对需要或伤害侵入的一种生理反应。
1.1 三大经典模型的提出
1967年,雅克夫列夫(Yakovlev)根据塞里应激模型提出“运动训练的超量补偿循环”模型[74];1968年,康希尔曼(James Counsilman)也由应激理论提出“运动训练的高级适应循环”[27];20世纪70年代,马特维也夫(Matveyev)则综合雅克夫列夫和康希尔曼的模型提出了“超量补偿循环”和“代偿适应”模型[6,27],并于1981年被翻译成英文出版[58]。
雅克夫列夫和康希尔曼均赞同塞里基础理论,认为物理负荷的反应与报警反应是一致的,并以工作能力下降或疲劳发生的形式出现。雅克夫列夫指出,“适应的发生是一个有次序的循环状态,即‘负荷-无负荷-加大负荷-无负荷’的训练恢复理论模式”,而康希尔曼的高级适应模式是以“负荷上再加负荷”理论为基础的。雅克夫列夫虽然认识到了引发不适应的潜在因素,但认为恢复前又有负荷;而康希尔曼则指出,负荷高到某一特定点后,机体承受负荷时也有适应发生,并不再继续恢复,但没有说明为促进完全恢复,而降低负荷。
直到马特维也夫,才综合了二人的观点,将“负荷-适应循环模式”解释得更为准确,同时,还强调了运动训练过程甚至需要处于一种长期不完全恢复和训练末超量恢复的状态,这种状态只在训练循环末期,通过降低负荷来实现,累积训练的整体效应,进而带来竞技能力更好、更大的增长[6,27],亦即运动训练中的超量恢复模型。然而,超量恢复理论的质疑者提出了新的模型,如1988年,德国学者马德尔(Mader)以适应理论为基础,从细胞代谢的层面提出了“机能储备模型”的训练适应模型,随后,诺依曼( Neumann) 也提出“改变-适应的时间动态”理论[2]。前者认为,运动训练适应过程是对原本存在“适应储备”的挖掘;后者则提出,运动训练刺激是对机体平衡状态的干扰与重建,是一个不断“适应”与“提高”的过程。
可以看出,上述理论均是建立于塞里的“应激理论”而提出的。事实上,应激理论在生理学和心理学领域已得到广泛应用,其实用性解决了不少生理和心理现象,因此,也应用到运动训练活动中。然而,应激理论是否无懈可击?我国生理学奠基人之一的蔡翘院士曾质疑“应激理论”[1]:
1.“应激也可能引起特异性适应症,实际上,内、外环境的不利因素所引起的反应是极端复杂的,视其性质、强度、作用时间及机体技能状态之不同而各有差异,而且很多反应是链锁性的,互相影响,互相调节,甚至互为因果的,很难硬性划分特异性和非特异性。”因此,运动训练引起人体生理和心理反应亦如此。
2.“生理应激过程的3个阶段,即对刺激的直接反应及代偿反应、对刺激的部分或全适应、刺激停止后的恢复过程是互相穿插,互相交联的,恢复和适应很难明确分开,‘神经-肌肉’一面劳动,一面恢复,所以不致于发生疲劳。”就运动训练而言,训练与恢复的节奏控制也充满复杂性,很难具体而明确。
3.“内环境的调节是多方面的,如神经系统调节、‘神经-体液’的调节、单纯体液途径的调节,还有细胞代谢的自我化学调节,因此带来适应有难易之分、应激适应时间的长短之分、应激机制极端复杂等特性。”在运动训练中,训练适应过程同样处于一个极其复杂的调节与控制当中,除了训练方法得当,训练的艺术性同样不容忽视。
4.“生理应激的恢复过程,是指应激反应及适应反应逐渐消失,恢复了体内环境到未被刺激前的情况,包括恢复内环境的稳定性和恢复到原来情况。但任何一种剧烈反应后的恢复过程总在波动中完成,源于需要多方面的调节才能实现,生物的自动控制并不那么精确。应激作用强,时间长和适应的程度好,恢复就慢。但恢复时间并不与适应程度成反比,因为所谓特殊刺激都是寻常所见的,所以其作用的总时间总要比正常条件的刺激短得多。”就运动训练而言,恢复内环境的稳定性是运动训练需要的恢复,恢复到原来情况是运动训练杜绝的恢复。此外,马特维也夫传统分期理论对多年系统训练的有序分期与组织是否更有利于建立一种更长的刺激产生适应而缺乏强度?板块周期理论的大强度但时间短是否会影响训练适应程度等?值得证实[3]。
基于蔡翘院士的上述观点,本文认为,如果以“应激理论”的非特异性原则来指导运动训练亦会存在局限。
1.2 “超量恢复”理论模型研究进展
鉴于超量恢复周期模型相关支持理论的最新研究进展,有必要对其演进史进行重新梳理。早在20世纪40年代,在一些生理、生化的研究论著中就出现类似“超量恢复”现象的研究[2]。但直到Hultman E和 Bergström J(1966)在《自然》杂志上发表《运动后肌糖原的合成:因素存在于人体肌细胞中》一文,才正式明确存在肌糖原的“超量恢复”现象。研究中,两名受试单腿骑功率车(一人用右腿骑车,左腿休息,另一人相反) ,以同样1 200 kpm/ min的负荷,5~20 min一次间歇,直至力竭(无法继续1 min以上的运动)。接下来,3天内安排受试者进补专门的碳水化合物食物(力竭运动后,进食碳水化合物比进食高脂肪和蛋白质及混合食物对糖原恢复更有利[15]),并分别对股四头肌糖原(认为股四头肌糖原代表糖原含量水平)进行检测。结果显示(图1),第1天,测试腿糖原含量就远远超过对照组;第2天,受试者测试腿中糖原高出近2倍,运动导致肌肉糖原耗竭后,反而能提高其糖原重新合成能力。但其机制尚不明确,研究者认为,可能是一个或多个刺激因素,导致肌糖原合成或者细胞膜的影响,刺激了糖原摄取。研究者亦指明,这项研究可用于正常人的体育锻炼(或运动员的运动训练)和糖尿病人碳水化合物饮食的建议。翌年,两人提出,工作肌肉的现有肌糖原含量是维持长期大负荷工作的决定性因素,且这种工作能力及其糖原含量可通过糖原耗竭后进食不同食物来调节[15]。这两篇文献在描述力竭性工作肌肌糖原超量恢复现象及其营养补充领域起到引领性作用。随后,1984年,James D E和Kraegen E W研究运动训练对肌肉和肝脏中的糖原、糖原合成酶、磷酸化酶的影响,也发现运动训练可以对碳水化合物消耗具有保护作用。运动训练引起比目鱼肌的糖原、糖原合成酶增长60%~150%、糖原含量增长50%~70%。于是,肌糖原含量越高,运动疲劳发生的时间越晚,糖原含量尤其对长期、剧烈运动十分重要。因此,可用于指导耐力运动训练获取最佳竞技状态[38]。
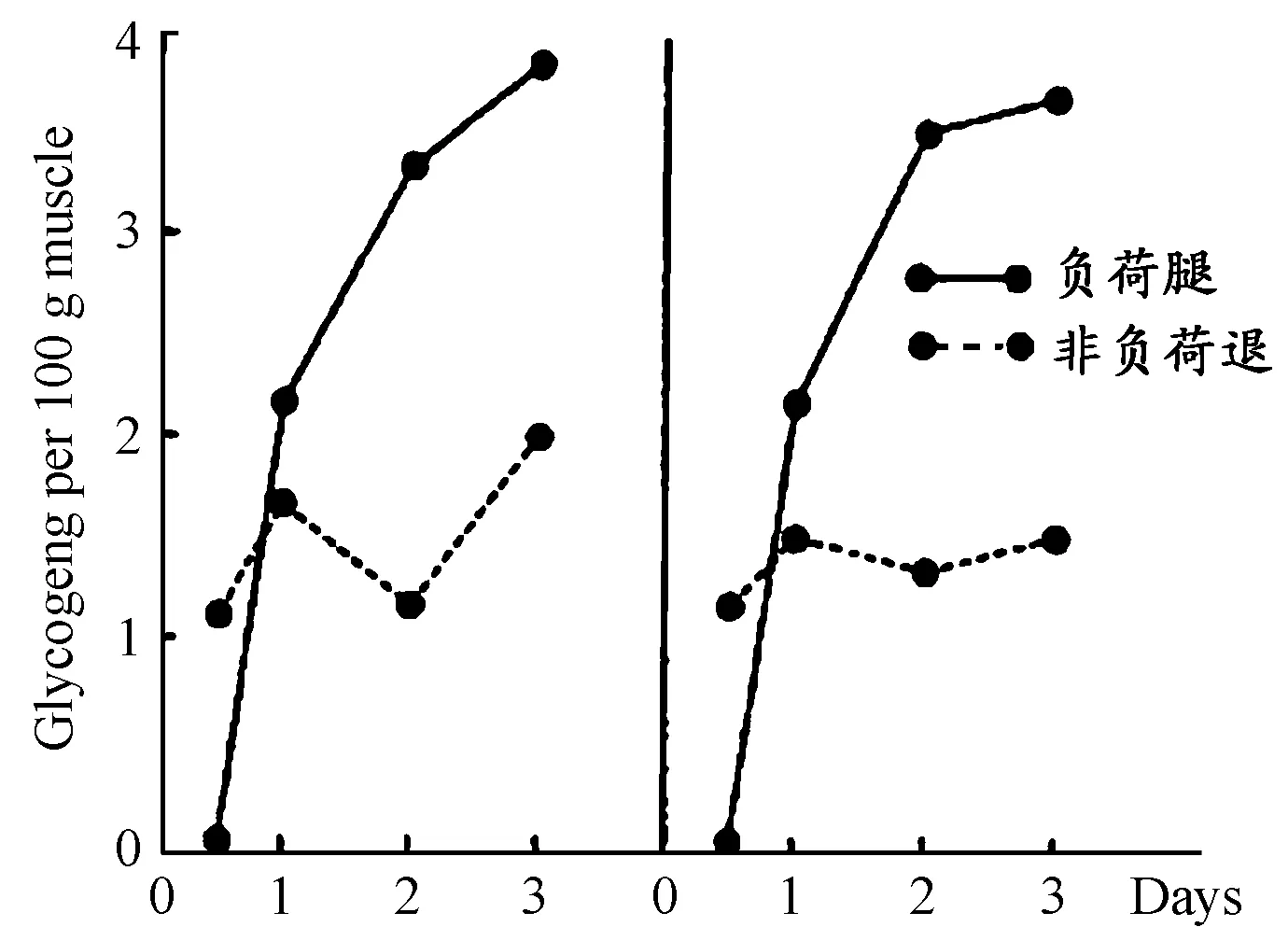
图1 负荷腿与非负荷腿股四头肌糖原含量示意图
根据对ISI文献中的跟踪调研发现,自Hultman E和Bergström J 1966和1967年连续发表文章阐明长时间力竭运动后,补充碳水化合物带来显著的肌糖原超量恢复现象以来,对运动饮食或运动营养补充、肌糖原与竞技能力的关系的研究便不断涌现,且以运动营养补充促进肌糖原更好的恢复方面的研究更为普遍。本文对1966年迄今为止,引用频次在200~500次以上(21世纪选择100次以上)的文献进行筛选,选取不同时期代表性研究结论进行了归纳(表1)。
同时,据现有文献研究发现,将“超量恢复”用以解释运动训练对人体机能能力的影响,源自1972年原苏联列宁格勒体科所的雅克夫列夫(前文提出根据1994年康希尔曼著述是1967年)的研究,1977年首次完整地提出了“超量恢复”学说,并将其作为解释运动训练效果的理论基石。可以看出,雅克夫列夫所提出的运动训练中的超量恢复应用是在Hultman E和 Bergström J发现肌糖原超量恢复现象之后,而马特维也夫更是继承了二者的学说。超量恢复理论自前苏联传至中国并成为我国运动训练的理论支柱之一[5],但在进入21世纪前后,超量恢复理论遍受质疑,认为应被“体力波”训练理论和“适应性”训练理论所替代[4]。
为了解国际最新研究进展,本研究在南京大学杜厦图书馆通过ISI web of knowledge数据库以“supercompensasion”为主题(2015/11/27)进行搜索,共获得964条文献记录,前5个研究方向分别为生物化学与分子生物学、生理学、内分泌代谢、运动科学、解剖形态学,其中,运动科学方向有147篇文献。被引频次最高的两篇是《运动训练与饮食对肌糖原和运动能力的影响研究》[75]和《过度训练和恢复》[50]。而以“supercompensasion”为标题进行搜索,共获得155篇文献,运动科学方向共67篇,分析发现,主要涉及力竭运动后肌糖原超量恢复、碳水化合物超量恢复以及近期关于脑糖原超量恢复的研究[33,56-58,81],且肌糖原和脑糖原超量恢复都是基于长时间力竭性运动之后产生,二者都可以提高运动训练中所需能源储备。

表1 肌糖原超量恢复相关代表性文献研究内容与进展一览表
值得关注的是,脑糖原的研究曾一度因争议被搁置,但21世纪初又再次逐渐引起科学家们的重视,图2是以“主题:(Brain glycogen),时间跨度=所有年份”,分别于2014年1月25和2015年12月27日在ISI web of science中搜索到文献9 171条和11 142条,其中,2002年以来出版文献数和2003年以来文献被引用数急剧递增,近1年半每月平均增加近100篇研究文献。

图2 以“Brain Glycogen”为主题在ISI中搜索到的文献计量示意图
脑糖原比肌糖原和肝糖原浓度相对要低,但却是脑葡萄糖含量的几倍。成年人脑糖原主要存在于脑星形胶质细胞中,是神经元活动的主要能源,它由神经递质和周围葡糖糖浓度来调节。可靠研究表明,低血糖条件下,星形胶质细胞糖原分解为乳酸,并传递到相邻的神经元和突触,部分乳酸为轴突提供能量[14,18],进而延长神经和轴突在高强度活动下的稳定性能[17]。在长时间力竭运动之后,糖原超量恢复,不仅存在于骨骼肌中,而且在脑中也存在,进而提高动作控制(运动耐力)和认知功能[14,57]。同时,脑糖原可能是剧烈运动引起中枢疲劳的部分机制所在[57]。2010—2013年已经有研究提出,“长时间力竭运动后脑糖原出现超量恢复现象”(图3)[56-58]。新近的研究成果表明,“脑糖原”和“运动引起脑糖原超量恢复”的研究结论将为我们重新审视“超量恢复理论”推开新的理论窗口。由此,本文认为,“超量恢复理论”的提出已经过去近半个世纪,对原有理论的质疑本是科学的积极态度,但质疑的同时应汲取和跟进最新的研究动态,同时,我们自身也不能忽视基于西方科学实证传统的同步推进,否则可能容易丢失“传统科学”与“科学的传统”。
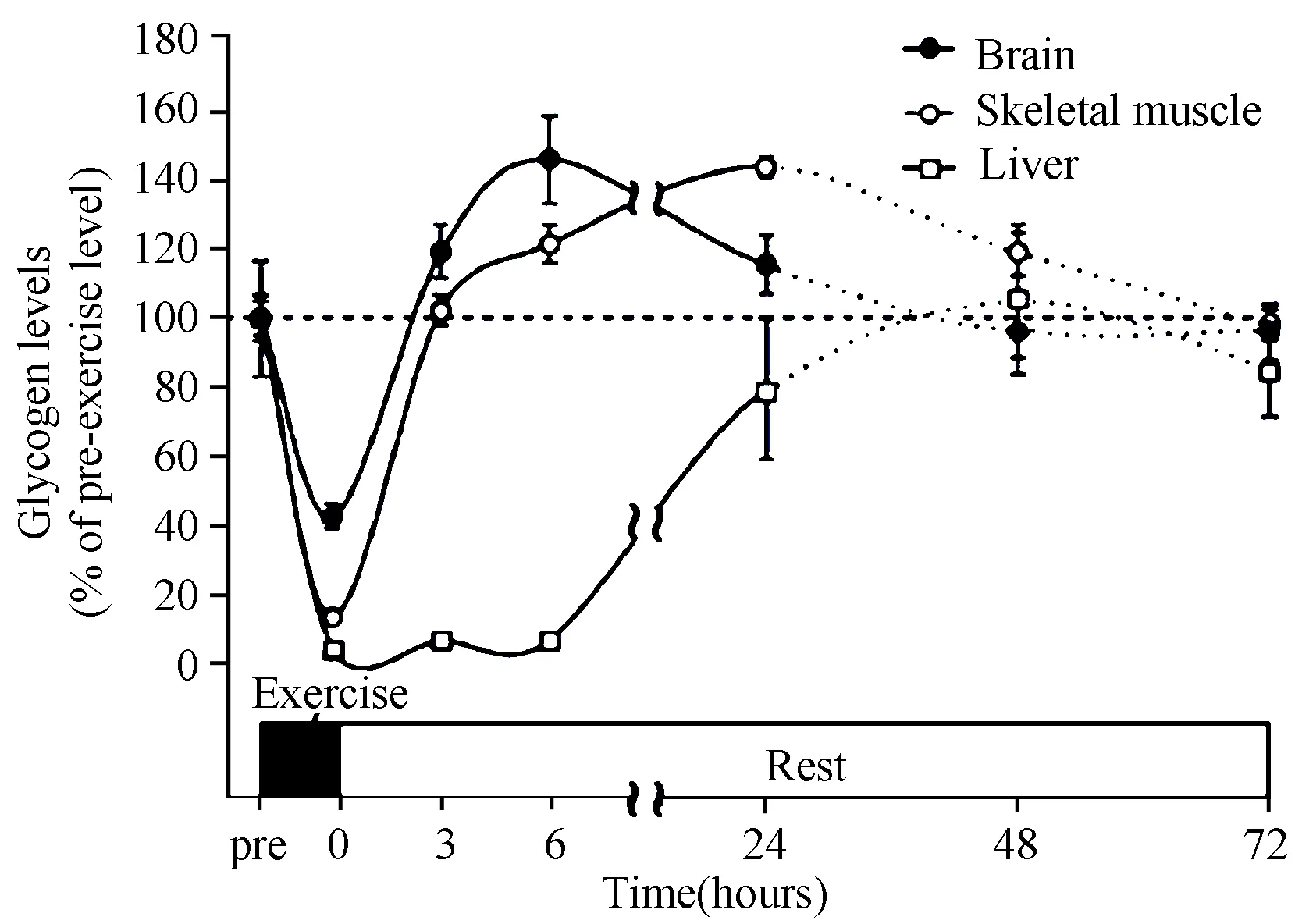
图3 力竭运动后脑糖原、肌糖原和 肝糖原的动态变化示意图
2 生理实证与数学推理相结合模型及其演进
随着生理模型不断提出并应用于运动训练实践,人们也逐渐发现,仅此已不能满足运动训练实践中的复杂性需求。1960—1970年代美苏冷战,激发了美国向“科学和数学”看齐,随后的计算机信息技术更是推动现代科技的数学化和信息化发展,运动训练理论模型也随之改进。本文例举自20世纪60年代至21世纪初的3个经典模型,阐述其进路。
2.1 临界功率(Critical Power,CP)模型
Monod和Scherrer 1965年正式提出用于测试动态和静态肌肉做功的“临界功率(Critical Power,CP)”模型(Scherrer等,1954;Scherrer和Monod,1960;Monod和Scherrer,1965)。CP模型是指最大输出功率(P)和局部肌肉疲劳前保持最大功率的时间(T)存在双曲线关系:W总=CP×T+W′,其中,CP代表有氧做功部分(单位:W),W′是无氧做功部分(单位:J)。并由此推导功率为:P=CP+W′/T,而如果一个项目的无氧功率高于CP,则最大功率持续的时间为:T= W′/(P-CP)(图4)。1980 s早期,Moritani等[59]和Whipp等[81]将此概念扩展到全身的训练功率中,发现不同受试者在功率自行车训练中达到疲劳时,CP与气体交换的无氧阈显著相关,CP和能量储备之和与最大摄氧量也显著相关。
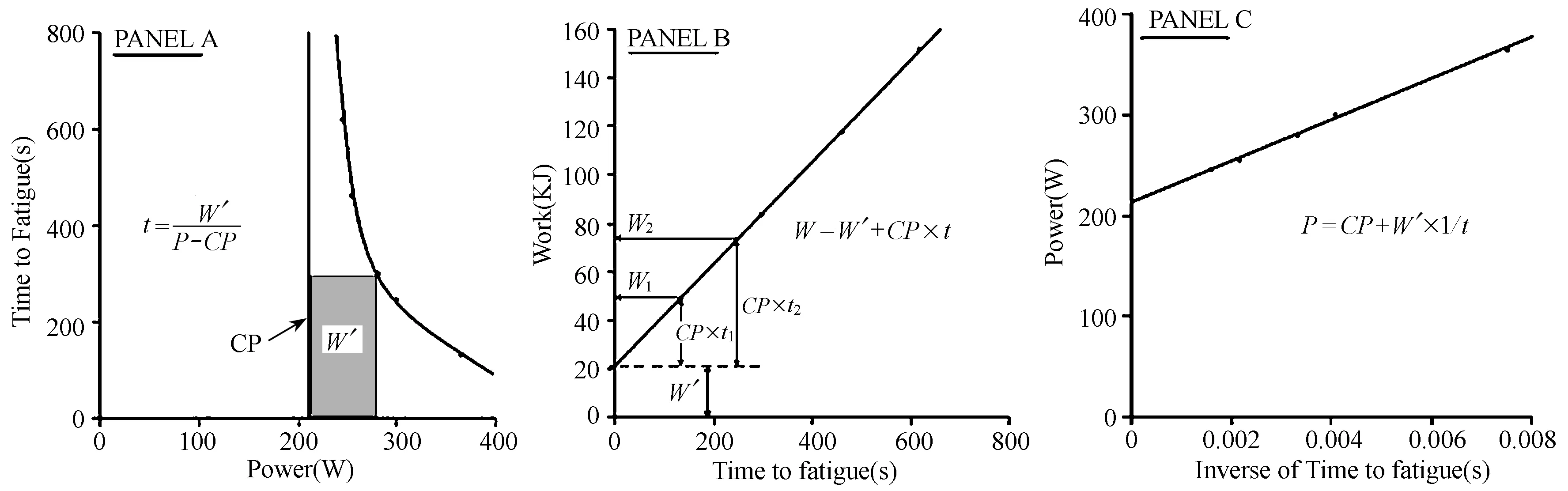
图4 二变量CP模型的3种主要表达形式
CP最初的理念来自于功率P=W/t,以其表示在不同功率时,做功与疲劳出现之前所持续的时间之间的线性关系。模型的两个变量中,P是功率,T是功率持续的时长。对于游泳和跑步项目而言,P和CP可用速度(S)和临界速度(CS)表示,W′用距离D′表示。传统方法是通过对不同持续时间的最大恒定功率测试,获得CP和W′。但是,由于多个单元测试导致不可控的干扰因素会影响其结果。为消除传统测试方法的不足,有学者将其改进为3 min最大强度测试[78]。
Morton[61]将CP模型的假设条件归纳为4点:1)两类能量物质输出功率:有氧和无氧;2)有氧能量是无限供能的(即可以在CP强度或CP以下强度长时间无限运动),但其转化为做功的效率是有限的;3)无氧供能在转化做功效率上也是无限的(即最大输出功率或最大速度是无限的),但其能量是有限的;4) W′耗尽时将出现力竭。目前,CP模型已经在一些运动项目中较好地评估了肌肉能力表现,尤其在自行车、跑步、游泳和划艇[44]等项目中。然而,有研究也提出,这些假设过于牵强,该模型对最大功率持续时间限定2~30 min才适用[61],通过W′产生最大功率也是有限的,因为难以知晓受试者的最大功率工作能力是否足够大且稳定,而W′在力竭时也不可能耗尽[44]。总体而言,会导致CP高估,而W′低估。
为弥补不足,Morton[37]创立了三变量CP模型(图5)。假设最大输出功率是无限的,且力竭发生在W′耗竭之时。修正后放宽了两变量模型的要求,渐近线在t=0时也存在,T无限接近于0,修改后的数学表达为(1):
图5 三变量CP模型
注:与二变量模型相比,同样的数据,三变量模型CP值较低,且W′值较高。三变量模型的第3个变量Pmax代表最大的瞬时功率[37]。
t= W′/(P-CP)+K,(K<0)
(1)
K是渐近线,假设为负值。因为最大功率(Pmax)只能在瞬时产生(即力竭时间为0),则意味着:
t= W′/(P-CP)+ W′/(CP-Pmax)
(2)
Morton进一步假设了训练中可达到的最大输出功率有赖于W′的剩余量大小。通过这样一个数学推理补充,他将上述方程中最大功率变换为“最大瞬时功率”,P且被证明与剩余W′有一个线性函数[37],因此,改良后的CP模型假设力竭时W′耗尽,变为力竭发生在最大功率小于理想输出功率时。此外,t是总的持续时间,n是间歇次数,Tw和Tr分别是每个间歇的功率持续时间和每个间歇的恢复阶段,Pw和Pr分别是做功和休息阶段的输出功率[60]。同时,模型的合理性还需要以下约束条件:
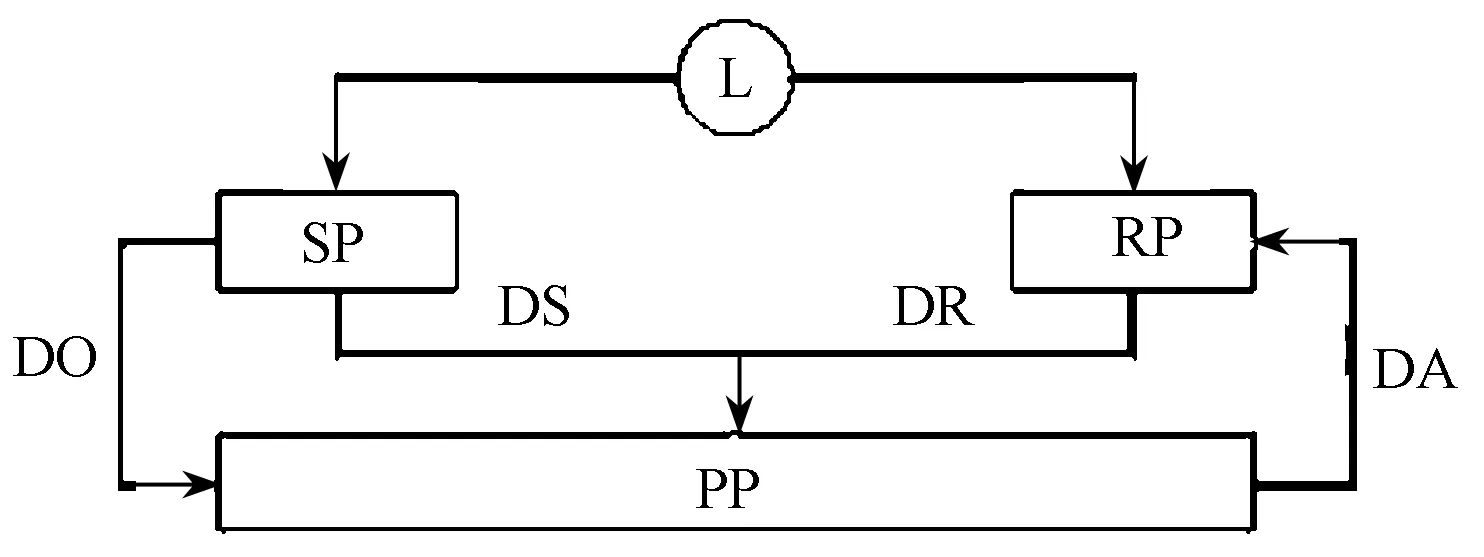
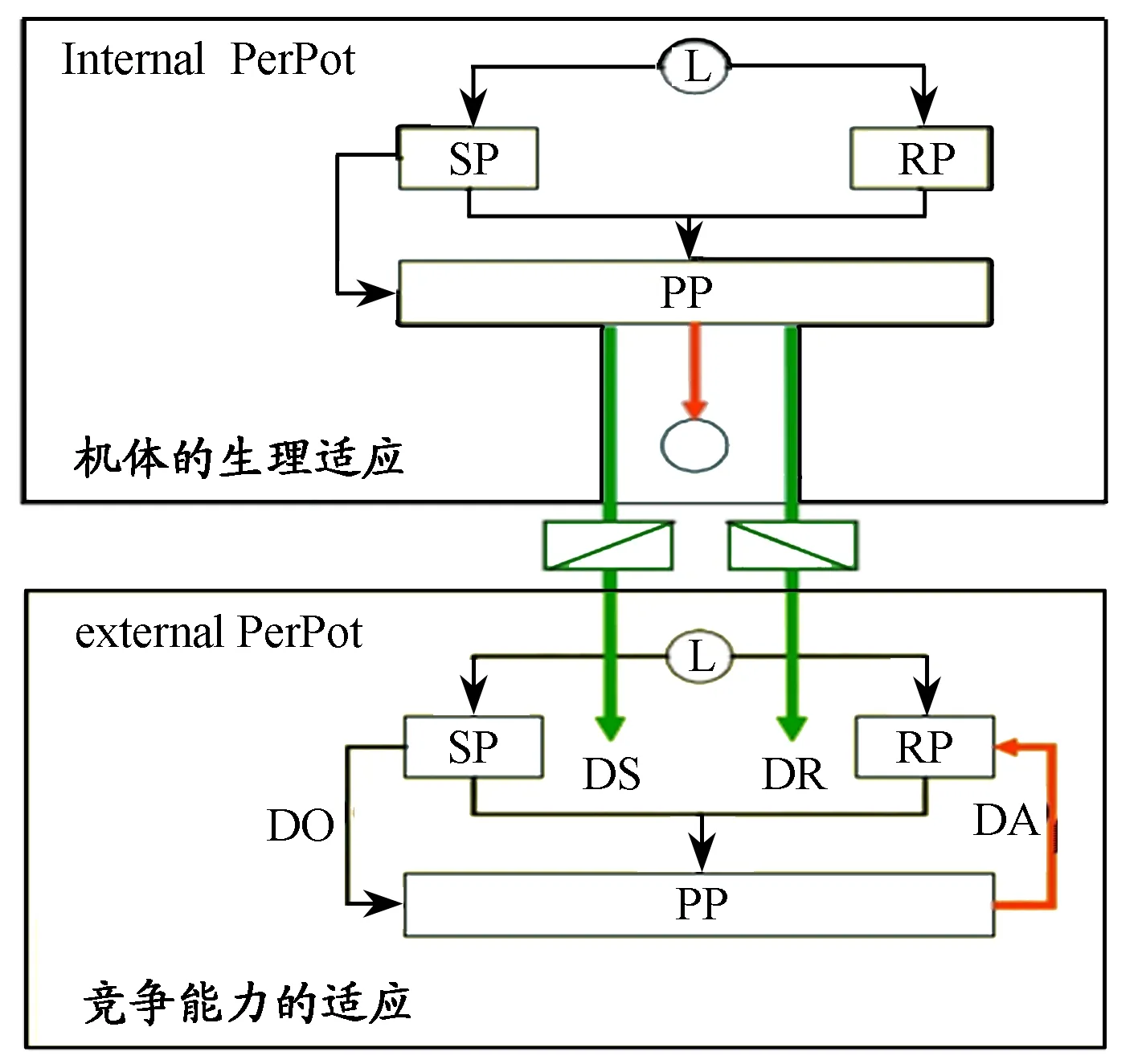
0≤Pr (3) 可见,Morton和Billat[60]将两变量CP模型进行补充,更有利于优化间歇运动的训练处方。概言之,CP模型的突出优点,是将能量输出和机械效率统一于一个变量关系中,且简单易行。此外,一些运动训练实践中难以实现的人体运动参数通过数学转换来实现和优化,使得运动训练负荷与竞技能力增长模型在原有生理模型的基础上更能向指导实践推进一步。 2.2 训练负荷-效应模型(impulse-response,IR) 1975年,Banister E W提出“训练负荷-效应模型(impulse-response,IR)”[11],试图解释运动训练与竞技能力提高的个性化预测问题,该模型是运动训练理论与方法中最具代表性的经典模型之一,陆续得到研究和拓展(Calvert等,1976;Busso等,1997,2002;Busso,2003)[21]。其基本定义:“对前期训练负荷在特定时间产生累积效应的竞技能力量化描述。”[76]通过简洁地描述个体的训练“负荷-效应”关系,将非线性复杂因素个性化地设定在一个单一框架中,其目的是通过运动训练计划定量数据和运动成绩预测运动训练效果,最终实现优化训练质量。原始研究是对游泳训练负荷和成绩效应进行模型化[11]。此后,该模型在其他运动项目中也得到应用(如跑步、游泳、自行车、铁人三项、举重、链球等)。 IR建模之初,为检验运动训练引起竞技能力的变化过程,Calvert等[26]提出,竞技能力的动力学和一阶系统行为类似,系统行为随时间而变化,并可利用常微分方程(ODEs)建模。Calvert等因此提出常微分方程可以用以量化竞技能力的时间动态变化过程,随后用标准数学技术进行了求解[26]。然而,在后期发现方程难以解释近几个月对游泳训练采集的所有竞技能力(成绩)数据,且游泳运动员的竞技能力在负荷增加时反而下降,Calvert等对原方程又进行了修改,根据训练带来积极与消极的两种效应,试图建立一个二元系统,并惊喜地发现,二元系统方程与最初一元系统方程是一种形式。 具体而言,该二元系统模型假设运动训练负荷存在2个相反的函数效应关系:一是正向效应(PTEs),致使运动员机体处于健康状态,并带来运动成绩或竞技能力提高;二是负向效应(NTEs),致使运动员短期或长期处于疲劳状态,并带来运动成绩或竞技能力停滞或下降,且对已知的训练负荷,开始NTE超过PTE而导致竞技能力下降,然而,NTE随时间消减得更快,最终PTE超过NTE,竞技能力增加。其中的PTE和NTE,分别与测得的健康与疲劳生理学参数均存在对应的量化关系,如女性跑步运动员的铁状态生物标志物的动力学、肌细胞损伤的标志物(血清酶升高,如肌酸激酶、乳酸脱氢酶和天冬氨酸氨基转移酶[13])在NTE时活跃;通气阈与PTE显著相关,NTE与个人的情绪状态问卷(反应感觉疲劳)中度相关[82];同样,血清激素水平与NTE、PTE也存在显著相关[20]。IR模型的具体表达见图6、图7。 图6中,A是IR模型的求和方程,B是IR模型的递归方程,两方程中P(t)是时间T时的竞技能力,P(0)是初始的竞技能力水平;K1和K2分别是健康和疲劳强度因子;τ1和τ2分别是健康和疲劳衰减的时间常数;W(s)是已知自第1周到赛前一段时期的周(或日)训练负荷。这些变量都是以个体的反应来解释的[62](Mujika等,1996)。图7是对训练效应定性特征的模拟。模型是以日训练负荷作为输入变量的,因此,定义训练负荷为:训练负荷=训练强度×持续时间。 图7 竞技能力随运动训练负荷变化的IR模型定性描述 注:底部是日常训练负荷的时间函数。运动员在开始的120天中,每天进行100 TRIMPs的负荷训练,随后进行7天减量训练,每天训练负荷为30 TRIMPs,接下来停训。PTE、PNE和竞技能力通过TRIMPs来计算,其参数如下:p(0)=500,K1=1,K2=2,τ1=27,τ2=10。 训练强度一般采用Eric Banister提出的TRIMP,即以每分钟的心率HR来度量强度,同时还考虑到高强度负荷比同时长的低强度负荷的代谢过程更费力[12],具体表达:1)TRIMP=t×k×FHRR;2)FHRR=(HR平均-HR休息)/(HR最大-HR休息)。其中,t是训练单元持续的时间(min),FHRR是心率储备分数,k=0.64e1.92×FHRR(男)或者k=0.86e1.67×FHRR(女)。 此外,还有研究提出,在自行车训练中采用训练强度得分(TSS),这种方法考虑到爬坡、风速等对不同强度训练输出功率的影响,而取平均功率来度量其强度[8]。随着人们对IR模型的深入认识和拓展,IR模型不仅应用于竞技运动,而且可用于个人疾病和损伤康复,如Le Bris和Coauthors用这个模型预测心血管病人在康复计划中断或被干扰时的功能能力损失情况,以及第2阶段康复病人每周2~3次和每周5次康复训练的治疗效果比较[53],Jomenez和Skiba等还运用这一模型预测膝盖手术病人的功能能力恢复周期[32]。 当然,IR模型也存在明显的缺陷:1)需要受试运动员保持激昂的测试动机来全力配合训练数据的采集,且测试比较频繁(至少每周1次),因此经常表现出预测能力偏差[77];2)测试需要购买昂贵的设备;3)IR模型假设健康和疲劳对训练负荷的应答是线性的,但现实中机体对训练的适应是有限的。为反映这一生理现实,Hellard等(2005)提出对IR模型的修订,日训练负荷[w(s)]用饱和HILL方程进行转换,将原模型中对训练负荷的线性自适应刺激调整为非线性(公式4、5)。 (4) (5) Busso及其同事(1997)同样也发现,不随时间改变的参量可能不适合于不同训练计划的竞技能力模型,因此,为找出随时间变化的参量,Busso等用一个递归最小二乘算法使得变量随时间变化,并发现改变后的模型更能与成绩数据拟合。同时,通过增加训练频率来提高训练负荷引起增益变量K1和K2变化,这促使Busso(2003)对IR模型进行了修改,将K2重新定义(公式6),修改后的负面效应可表达为(公式7): (6) (7) 至此,可进一步将IR模型定义描述为:基于前期竞技能力水平加上正向训练效应(PTE)减去负向训练效应(NTE)所得的竞技能力变化值。与CP中的W′不同,模型参数可能不仅仅是单一的生理学变量,而是反映训练效应的动力学多变量聚合影响。并且,通过输入具体运动员数据和变量值,模型可以个性化。修正的二元系统IR模型为运动训练适应的动力过程提供了一个解释窗口,基于此假设,IR模型能解释大多数收集到的成绩数据(一些研究中R2>0.90)[82]。它对制定运动训练计划具有很强的指导意义,具体表现为:1)将训练原则整合到统一的一致性框架中;2)以估测的训练负荷为输入,强调运动训练是一个生理应激过程,训练扰乱了体内平衡,而恢复则需要大量器官系统的适应,如免疫、神经和内分泌等系统对训练应答的协调,如果施加持续过量训练负荷将对这些系统带来破坏,导致过度训练发生,因此,将量化不同阶段的训练负荷作为检查施加适宜负荷的一种途径;3)该模型突出了竞技能力适应是一个训练应激的聚合功能,意味着不是单一训练内容影响竞技能力,而是持续日常训练、超负荷和减量共同带来高峰竞技能力。 然而,运动训练与竞技能力增长关系又表现为强烈的个性化差异[10,64],且受到多种因素的影响,如不同训练背景、生理学因素、技术因素、专项因素等[34]。以实验为导向的理论模型(实验事实的描述归纳),常存在诸如难以形成对训练计划设计的长期纵向研究、无法明确比较不同干预 因素的影响(且干预往往是正常训练的一种补充)等问题。此外,这类研究通常选取的受试较少,缺乏统计力度。任何一个运动训练模型都不是全面而精确的,CP和IP模型将生理学与数学相结合,而其中的数学模型往往正是复杂生物系统的有效表达和假设评估的有效手段,并予以个性化[52],这些优点是此前的生理模型无法实现的。从国际上的发展态势而言,这可作为对原有运动训练理论与方法模型的一种补充,正如恩格斯所言:“任何一门科学的真正完善在于数学工具的广泛应用。” 2.3 竞技潜能元模型(Performance Potential Metamodel,PERPOT) 2000年,德国美因茨大学计算机科学系的Jürgen Perl提出“竞技潜能元模型”(Performance Potential Metamodel,PERPOT)[68,69],一种“训练负荷与竞技能力交互关系”的理论模型。由于超负荷训练会引发超量恢复或者过度训练两种相反的效应,他基于数学分析指出,随时序变化的特殊效果和竞技能力稳定性只能用一个对抗性结构来建模, 模拟一系列潜在变量之间的延迟传输作用线的交互作用,这在自控制神经系统中已得到广泛应用。其首次对运动训练中的定性现象进行描述,通过对抗系统动力学(训练负荷会引起正反两方面的效果:提高竞技能力的应答流和降低竞技能力的应变流),且主要以应答流与应变流的延迟时间(时滞)不同来控制运动训练中生理适应的正负效果;也是首度尝试采取计算机软件模拟训练负荷对竞技能力作用的短期与长期反应的动力过程,用于优化训练计划和预防过度训练的计算模型(图8、图9)[66]。 图8 PerPot模型基本结构图 图9 依据训练负荷(红线)和相关流时滞的竞技能力走势图(蓝线) 其中,L是训练负荷,SP是训练导致的机体应变潜能,RP是训练导致机体的应答潜能,PP是机体表现竞技能力的潜能,或称竞技潜能;DS是应变流时滞,DR是应答流时滞;DO是应变潜能的溢出流时滞(当训练负荷超过机体应变潜能承受限度时产生),DA是机体内部衰减流(作为下一次训练的起始状态)。左侧的DS和DO对竞技能力潜能PP有减法作用,右边的DR对PP有加法作用,一般情况下是同时存在的动力对抗关系。然而,当训练负荷超过机体应变潜能限度(机体存在一个适应的自然限度)而导致应变流溢出时,就表现为过度训练,进而减小竞技能力潜能。为解释两种时滞值的大小对运动训练效果产生的重要影响,图9展示了引发超量恢复与否的作用线。 图9显示,施加的训练负荷一样,应变流时滞(DS=4)大于应答流时滞(DR=2)时的竞技能力表现明显低于应答流时滞(DR=4)大于应变流时滞(DS=2)时。后者才能出现竞技能力的超量恢复效应,因此,当前的竞技能力潜能状态和产生应变流、应答流的时滞将对竞技能力的动力学行为产生极为重要的影响。 对于PerPot元模型如何反映训练负荷、效应时滞和竞技能力之间长期的相互作用与反馈,Perl在原有基本结构的基础上进行了改进:要求模型不仅反映竞技能力输出,而且训练效应时滞值都应该由训练负荷输入来调控。原有的基础模型(图8)显然无法囊括,进而提出“二维水平的PerPot模型”(图10)[66]。该模型中,内部PerPot模型的“竞技能力”PP调控着外部PerPot模型中的时滞DS和DR (即人体器官的竞技能力状况直接影响运动训练引起的两种时滞),并且两个模型均接受同样的训练负荷输入L。这样,运动训练不仅通过优化竞技能力生理表征物质和传输率,而且还通过提高生成和传输组件的性能和速度来提高外在的竞技能力。 图10中,内部(人体器官)竞技潜能(internal PerPot)表示竞技能力生理资源再生能力和运输速度,外部时滞变量(DS、DR)代表器官“竞技能力”,通过如下关系进行的调控:1)如果应变时滞比应答时滞小,系统的第一反应是积极的;2)否则是消极的。比如,为增加训练负荷引起的外部(可测量的)竞技能力的反应的途径,即减小应变时滞,或增加应答时滞。 此外,通过“二维水平的PerPot模型”可知,内、外部衰减模式是不同的。外部模型表示短期行为,其衰减竞技能力可以反馈给系统并可以被重新激活,而内部控制系统模型是长期行为,长期训练导致生理资源会被耗尽,进而影响竞技能力潜能,使其下降,模型中添加了一个衰减分支线(红色带圆圈)表示内部“竞技能力”的亏损。如此,特定的训练方式,如过度训练则会引发延后的竞技能力“崩塌”,如果竞技能力状态因此不见好转,这种竞技状态崩塌现象或将无以修复。 图10 通过内、外方式双向动力时滞控制的 长期效应过程的PerPot模型图 注:红线表示不同类型的内部和外部衰减流;绿线和方盒子表示外部时滞的内部控制 竞技潜能元模型是不同于IR模型的另一种认识方式,它们都是以运动训练引起“正负对抗效应”的假设为前提衍生而来。PerPot元模型的不同之处还在于引用计算机软件对这一生理适应进行模拟,融合了生理学、数学和计算机科学与技术进而将“训练负荷”与“竞技能力增长”模型注入了新的科技元素,难怪对IR和PerPot的对比实验研究指出,PerPot对运动成绩数据具有更高的拟合能力[69]。 塞里应激理论奠定了运动生理学等在运动训练中的领导地位,以生理学为理论基础的模型,揭示了运动训练负荷与竞技能力增长之间的关系。直接由应激理论衍生的模型,有“运动训练的超量补偿循环”模型、“运动训练的高级适应循环”模型、“超量补偿循环”和“代偿适应”模型即“超量恢复”模型等,其中,“超量恢复”模型是后期对前期应激理论的GAS模型在运动训练中的修正与综合。 但是,运动训练的生物学复杂性,使仅采用单一的生理模型已难以进行有效表达。于是,结合数学、物理学和计算机科学来转换和优化不失为一种更加合理的途径,如CP模型、IR模型、PerPot模型。其中的数学模型可以通过假设与验证,实现对复杂生物系统的有效表达,并可根据个体参数不同来实现模型个性化。融合计算机信息技术的“竞技潜能元模型”则通过现有数据,即刻优化训练计划和预防过度训练,借助其强大的运算功能,达到运动训练数据与目标成绩之间的高度拟合能力。生理学、数学、物理学和计算机科学与技术进而将“竞技能力增长”模型的全面性和有效性提升了一个层次。 “竞技能力增长”理论模型的演进,大抵也遵循着图灵奖得主吉姆·格雷(Jim Gray)在“科学的第4个范式”中的预测趋势,即依次经历“实验科学(自然现象描述)→理论科学(模型化,一般常模)→计算科学(通过计算来模拟复杂现象)→数据密集型科学(理论、实验和模拟相结合)”。当前,国际上竞技能力增长模型大都处于理论科学和计算科学阶段。 以奥林匹克运动为引领的运动训练活动,素来也是科学的试金石和科技的展示舞台,回顾历史,它与人类科学的进程惊人同步。本文对其中历经的经典科学理论模型进行了提要与简析,毕竟这些模型的建立与应用都得益于跨学科复合人才的共同努力,仅凭本文所述还远远不够,无论我们是否认识到或跟上这一科学步伐,事实是,倘若没有全体体育科研与运动训练实践者们的集思广益、勇于突破和直面挑战的勇气与魄力,运动训练理论与方法很可能长期处于被动或落后的局面。虽然运动训练理论与方法的“第4个范式”是否即将来临,尚无定论,但大数据研究在各个领域和行业已悄然铺开,趋势或逐渐明朗,就运动训练理论与方法乃至整个体育科学而言,关键还在于我们以何种心态和实际行动来迎接这场科学范式革命的机遇与挑战。 [1]蔡翘.Selye 应激学说与生理应激[J].生理科学进展,1963,5(1):1. [2]陈小平.运动训练的基石——“超量恢复”学说受到质疑[J].首都体育学院学报,2004,16(4):3-7. [3]胡海旭,邱锴,李恩荆,等.论运动训练分期理论与板块周期的关系[J].上海体育学院学报,2013,37(6):90-96. [4]黄巧.我国运动性疲劳与超量恢复理论沿革研究[D].重庆:西南大学,2012. [5]李庆, 李景丽, 顾扬, 等.现代运动训练周期理论的思考和讨论[J].体育科学, 2004,24(6):52-55. [6]陆一帆.游泳训练理论创新与实践[M].北京:北京体育大学出版社,2013. [7]王明辉,张淑熙.应激研究综述[J].信阳师范学院学报(哲学社会科学版),2003,(1):59-62. [8]ALLEN H,COGGAN A.Training and Racing with a Power Meter[M].VeloPress,2010:58-60. [9]ACHESON K J,SCHUTZ Y,BESSARD T,etal.Glycogen storage capacity and de novo lipogenesis during massive carbohydrate overfeeding in man[J].Am J Clin Nutri,1988,48(2):240-247. [10]AVALOS M,HELLARD P,CHATARD J C.Modeling the training-performance relationship using a mixed model in elite swimmers[J].Med Sci Sports Exe,2003,35(5):838-846. [11]BANISTER E W,CALVERT T W,SAVAGE M V,etal.A systems model of training for athletic performance[J].Aust J Sports Med,1975,7(5):61. [12]BANISTER E W.Modeling elite athletic performance[J].Physiol Test Elite Athl,1991:403-424. [13]BANISTER E W,FITZ-CLARKE J R.Plasticity of response to equal quantities of endurance training separated by non-training in humans[J].J Therm Biol,1993,18(5-6):587-597. [14]BARROS L F.Metabolic signaling by lactate in the brain[J].Trends Neurosci,2013,36(7):396-404. [15]BERGSTR M J,HERMANSEN L,HULTMAN E,etal.Diet,muscle glycogen and physical performance[J].Acta Physiol Scand,1967,71(2):140-150. [16]BERGSTRÖM J,HULTMAN E.Muscle glycogen synthesis after exercise:An enhancing factor localized to the muscle cells in man[J].Nature,1966,210(5033):309-310. [17]BROWN A M,RANSOM B R.Astrocyte glycogen and brain energy metabolism[J].Glia,2007,55(12):1263-1271. [18]BROWN A M.Brain glycogen reawakened[J].J Neurochemi,2004,89(3):537-552. [19]BUSSO T,DENIS C,BONNEFOY R,etal.Modeling of adaptations to physical training by using a recursive least squares algorithm[J].J Appl Physiol,1997,82(5):1685-1693. [21]BUSSO T.Variable dose-response relationship between exercise training and performance[J].Med Sci Sports Exe,2003,35(7):1188-1195. [22]BEELEN M,BURKE L M,GIBALA M J,etal.Nutritional strategies to promote postexercise recovery[J].Int J Sport Nutr Exe Metab,2010,20(6):515-532. [23]BLOM P C,HESTMARK A T,VAAGE O,etal.Effect of different post-exercise sugar diets on the rate of muscle glycogen synthesis[J].Med Sci Sports Exe,1987,19(5):491-496. [24]BURKE L M,HAWLEY J A,WONG S H S,etal.Carbohydrates for training and competition[J]. J Sports Sci,2011,29(sup1):S17-S27. [25]BURKE L M,KIENS B,IVY J L.Carbohydrates and fat for training and recovery[J].J Sports Sci,2004,22(1):15-30. [26]CALVERT T W,BANISTER E W,SAVAGE M V,etal.A systems model of the effects of training on physical performance[J].IEEE Transact Sys Man Cybernet,1976,SMC-6(2):94-102. [27]COUNSILMAN J E,COUNSILMAN B E.The New Science of Swimming[M].Prentice-Hall,1994:169-173. [28]CARTEE G D,YOUNG D A,SLEEPER M D,etal.Prolonged increase in insulin-stimulated glucose transport in muscle after exercise[J].Am J Physiol-Endocrinol Metab,1989,256(4):E494-E499. [29]COSTILL D L,SHERMAN W M,FINK W J,etal.The role of dietary carbohydrates in muscle glycogen resynthesis after strenuous running[J].Am J Clin Nutr,1981,34(9):1831-1836. [30]COYLE E F,COGGAN A R,HEMMERT M K,etal.Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate[J].J Appl Physiol,1986,61(1):165-172. [31]COYLE E F,HAGBERG J M,HURLEY B F,etal.Carbohydrate feeding during prolonged strenuous exercise can delay fatigue[J].J Appl Physiol,1983,55(1):230-235. [32]DANIEL D M,STONE M L,DOBSON B E,etal.Fate of the ACL-injured patient a prospective outcome study[J].Am J Sports Med,1994,22(5):632-644. [33]GRUETTER R.Glycogen:The forgotten cerebral energy store[J].J Neurosci Res,2003, 74(2):179-183. [34]HELLARD P,AVALOS M,LACOSTE L,etal.Assessing the limitations of the Banister model in monitoring training[J].J Sports Sci,2006,24(5):509-520. [35]HELLARD P,AVALOS M,MILLET G,etal.Modeling the residual effects and threshold saturation of training:Acase study of Olympic swimmers[J].J Strength Cond Res,2005, 19(1):67-75. [36]HTTP://en.wikipedia.org/wiki/Hans_Selye/2015/7/9. [37]HUGH MORTON R.A 3-parameter critical power model[J].Ergonomics, 1996, 39(4):611-619. [38]IVY J L.Muscle glycogen synthesis before and after exercise[J].Sports Med,1991,11(1):6-19. [39]IVY J L,GOFORTH H W,DAMON B M,etal.Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement[J].J Appl Physiol,2002,93(4):1337-1344. [40]IVY J L,KATZ A L,CUTLER C L,etal.Muscle glycogen synthesis after exercise:Effect of time of carbohydrate ingestion[J].J Appl Physiol,1988,64(4):1480-1485. [41]IVY J L,RES P T,SPRAGUE R C,etal.Effect of a carbohydrate-protein supplement on endurance performance during exercise of varying intensity[J].Int J Sport Nutri Exe Metab,2003,13:382-395. [42]IVY J L.Muscle glycogen synthesis before and after exercise[J].Sports Med,1991,11(1):6-19. [43]JAMES D E, KRAEGEN E W.The effect of exercise training on glycogen,glycogen synthase and phosphorylase in muscle and liver[J].Eur J Appl Physiol Occupat Physiol, 1984, 52(3):276-281. [44]JONES A M,VANHATALO A,BURNLEY M,etal.Critical power:Implications for determination of VO2max and exercise tolerance[J].Med Sci Sports Exe,2010,42(10):1876-1890. [45]JENSEN T E,RICHTER E A.Regulation of glucose and glycogen metabolism during and after exercise.[J].J Physiol,2012,590(Pt5):1069-1076. [46]JENTJENS R,JEUKENDRUP A E.Determinants of post-exercise glycogen synthesis during short-term recovery[J].Sports Med,2003,33(2):117-144. [47]JEUKENDRUP A E.Carbohydrate and exercise performance:The role of multiple transportable carbohydrates[J].Curr Opin Clini Nutr Metab Care,2010,13(4):452-457. [48]JEUKENDRUP A E.Carbohydrate feeding during exercise[J].Eur J Sport Sci,2008,8(2):77-86. [49]JEUKENDRUP A E.Carbohydrate intake during exercise and performance[J].Nutrition,2004,20(7):669-677. [51]KARLSSON J,SALTIN B.Diet,muscle glycogen,and endurance performance[J].J Appl Physiol,1971,31(2):203-206. [52]LAUFFENBURGER D A,LINDERMAN J J.Receptors:Models for binding,trafficking,and signaling[J].Int J Biochem Cell Biol,1996,28(12):1418. [53]LE BRIS S,LEDERMANN B,CANDAU R,etal.Applying a systems model of training to a patient with coronary artery disease[J].Med Sci Sports Exe,2004, 36(6):942-948. [54]MASSENGALE J D,SWANSON R A.The History of Exercise and Sport Science[M].Human Kinetics Publishers,1997:408. [55]MATSUI T,ISHIKAWA T,ITO H,etal.Brain glycogen supercompensation following exhaustive exercise[J].J Physiol,2012,590(3):607-616. [56]MATSUI T,SOYA H.Brain glycogen decrease and supercompensation with prolonged exhaustive exercise[M]//Social Neuroscience and Public Health.Springer New York,2013:253-264. [57]MATSUI T,SOYA S,OKAMOTO M,etal.Brain glycogen decreases during prolonged exercise[J].J Physiol,2011,589(13):3383-3393. [58]MATVEEV L P,ZDORNYJ A P.Fundamentals of Sports Training[M].Moscow:Progress Publishers,1981:56. [59]MORITANI T, NAGATA A, DEVRIES H A,etal.Critical power as a measure of physical work capacity and anaerobic threshold[J].Ergonomics, 1981, 24(5):339-350. [60]MORTON R H, BILLAT L V.The critical power model for intermittent exercise[J].Eur J Appl Physiol, 2004, 91(2-3):303-307. [61]MORTON R H.The critical power and related whole-body bioenergetic models[J].Eur J Applied physiol, 2006, 96(4):339-354. [62]MUJIKA I,BUSSO T,LACOSTE L,etal.Modeled responses to training and taper in competitive swimmers[J].Med Sci Sports Exe,1996,28(2):251-258. [63]MUJIKA I,CHATARD J C,BUSSO T,etal.Use of swim-training profiles and performances data to enhance training effectiveness[J].J Swim Res,1996,13(2):23-29. [64]MACDOUGALL J D,WARD G R,SUTTON J R.Muscle glycogen repletion after high-intensity intermittent exercise[J].J Appl Physiol,1977,42(2):129-132. [65]OLSSON K E,SALTIN B.Variation in total body water with muscle glycogen changes in man[J].Acta Physiol Scand,1970,80(1):11-18. [66]PERL J.On the long-term behaviour of the performance-potential-metamodel PerPot:New results and approaches[J].Int J Comput Sci Sport,2003, 2(1):80-92. [67]PERL J.Per pot:A metamodel for simulation of load performance interaction[J].Eur J Sport Sci,2001, 1(2):1-13. [68]PERL J.PerPot-a meta-model and software tool for analysis and optimisation of load-performance-interaction[J].Int J Perform Anal Sport,2004, 4(2):61-73. [69]PFEIFFER M.Modeling the relationship between training and Performance-A comparison of two antagonistic concepts[J].Int J Comput Sci Sport, 2008, 7:13-32. [70]PHILP A,HARGREAVES M,BAAR K.More than a store:Regulatory roles for glycogen in skeletal muscle adaptation to exercise[J].Am J Physiol-Endocrinol Metab,2012,302(11):E1343-E1351. [71]PIEHL K.Time course for refilling of glycogen stores in human muscle fibres following exercise-induced glycogen depletion[J].Acta Physiol Scand,1974,90(2):297-302. [72]RICHTER E A,GARETTO L P,GOODMAN M N,etal.Muscle glucose metabolism following exercise in the rat:Increased sensitivity to insulin[J].J Clin Invest,1982,69(4):785. [73]ROBINSON T M,SEWELL D A,HULTMAN E,etal.Role of submaximal exercise in promoting creatine and glycogen accumulation in human skeletal muscle[J].J Appl Physiol,1999,87(2):598-604. [74]SCHOLICH M.Circuit Training[M].Berlin:Sportverlag,1986:77-81. [75]SHERMAN W M, COSTILL D L, FINK W J,etal.Effect of exercise-diet manipulation on muscle glycogen and its subsequent utilization during performance[J].Int J Sports Med, 1981, 2(2):114-118. [76]TAHA T,THOMAS S G.Systems modelling of the relationship between training and performance[J].Sports Med,2003,33(14):1061-1073. [77]TAHA T. A new approach to monitoring exercise training.[J].J Strength Cond Res,2001,15(1):109-115.[78]VANHATALO A,DOUST J H,BURNLEY M.Determina- tion of critical power using a 3-min all-out cycling test[J].Med Sci Sports Exe, 2007, 39(3):548. [79]VAN LOON L J C,SARIS W H M,KRUIJSHOOP M,etal.Maximizing postexercise muscle glycogen synthesis:Carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures[J].Am J Clin Nutr,2000,72(1):106-111. [80]WEAVER S A, SHARPE S N, PEDERSON B A.Brain glycogen supercompensation in the mouse after recovery from insulin‐induced hypoglycemia[J].J Neurosci Res, 2011, 89(4):585-591. [81]WHIPP B J,HUNTSMAN D J,STORER T,etal.A constant which determines the duration of tolerance to high-intensity work[J].Fed Proc,1982,41(5):1591. [82]WOOD R E,HAYTER S,ROWBOTTOM D,etal.Applying a mathematical model to training adaptation in a distance runner[J].Arbeitsphysiologie,2005,94(3):310-316. [83]ZAWADZKI K M,YASPELKIS B B D,IVY J L.Carbohydrate-protein complex increases the rate of muscle glycogen storage after exercise[J].J Appl Physiol,1992,72(5):1854-1859. Theoretical Models and Its Evolutionof Athletic Performance Enhancement HU Hai-xu Objective:Later period of the 20th century,through two world wars,and usher in the era of "big science" of society,science and history background,this paper reveals and analyzes the theoretical models and its evolution characteristics of the athletic performance enhancement from that time on,and then clarifies the scientific basis and theory evolution characteristics.Method:mainly using methds of historical research and literature.Result:1) the generally evolution approach of theoretical models of the athletic performance enhancement is as follows:the physiological model of "cycle of suercompensation","cycle of super adaptation","cycle of supercompensation and compensatory adaptation" that based on the Selye's GAS (general adaptation syndrome) model in the 1950 s;to the computational models of CP(critical power),IR(impulse-response),PerPot(The Performance Potential Metamodel)that based on physiology,physics and mathematics,computer science;2)Physiology is an important basis of athletic training;3)Mathematical model by assuming and verification,to achieve effective expression of the complex biological system,and according to different individual parameters personalized implementation models;4)the PerPot that integration of computer information technology can use the existing data to optimize training plans and prevent excessive training immediately,with its strong operation function to achieve a good fit between athletic training data and target results.Conclusion:With other law of development of science and society,and the same to Jim Gray in "the fourth paradigm of science",data-intensive scientific theory or is also a scientific paradigm of athletic training theory and method,and needs to be based on this paradigm,build the next theoretical model of the athletic performance enhancement. athletictrainning;athleticperformanceenhancement;physiology;math;computationalscience;data-intensive 1000-677X(2016)02-0014-11 10.16469/j.css.201602002 2015-09-13; 2016-01-28 江苏省博士后基金项目(1402039C);南京航空航天大学基本业务青年科技创新基金项目(NR2015019)。 胡海旭(1983-),男,湖北黄冈人,讲师,博士,主要研究方向为运动训练理论与方法、人因工程,Tel:(025)84892463,E-mail:hhx100000@163.com。 南京航空航天大学 体育部,江苏 南京 210016 Nanjing University of Aeronautics and Astronautics,Nanjing 210016,China. G808.1 A

3 小结
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
体育科技文献通报(2022年3期)2022-05-23
灌篮(2021年22期)2021-11-23
红领巾·萌芽(2020年5期)2020-06-03
中国医药导报(2019年3期)2019-03-18
现代养生·下半月(2016年6期)2016-10-21
Coco薇(2015年3期)2015-12-24
对联(2011年16期)2011-09-18
求医问药(2009年7期)2009-08-31
中学生物学(2009年1期)2009-02-23
