
硝普钠对圣女果种子萌发至幼苗期碳氮代谢及关键酶活性的影响
2017-09-13 09:15张永福郭丽红蒋淑萍姚丽媛赵明方
河南农业科学 2017年9期
张永福,郭丽红,蒋淑萍,靳 松,姚丽媛,赵明方
(1.昆明学院 农学院,云南 昆明 650214; 2.云南省高校特色生物资源开发与利用重点实验室,云南 昆明 650214)
硝普钠对圣女果种子萌发至幼苗期碳氮代谢及关键酶活性的影响
张永福1,2,郭丽红1,2,蒋淑萍1,2,靳 松1,2,姚丽媛1,2,赵明方1,2
(1.昆明学院 农学院,云南 昆明 650214; 2.云南省高校特色生物资源开发与利用重点实验室,云南 昆明 650214)
为了探明硝普钠对圣女果种子萌发及幼苗生长的调控机制,以圣女果品种朱云为试验材料,研究不同浓度硝普钠处理对其碳氮代谢及关键酶活性的影响。结果表明,圣女果种子萌发至幼苗期,淀粉、总氮含量逐渐下降,可溶性糖、还原糖、蔗糖和可溶性蛋白质含量呈先上升后下降的趋势,游离氨基酸含量逐渐上升;硝普钠处理的淀粉、蔗糖及游离氨基酸含量均高于蒸馏水处理(CK);圣女果种子萌发至幼苗期,淀粉酶、蔗糖磷酸合成酶(SPS)和谷氨酰胺合成酶(GS)活性呈下降趋势,硝酸还原酶(NR)和谷氨酸合成酶(GOGAT)活性呈先上升后下降的趋势;CK和0.25 mmol/L硝普钠处理的蔗糖合成酶(SS)活性呈下降趋势,0.50、1.00 mmol/L硝普钠处理SS活性呈先上升后下降的趋势,且硝普钠处理增强了淀粉酶活性;各处理碳含量逐渐下降,碳氮比除CK呈先上升后下降外,其他硝普钠处理均呈下降趋势;此外,硝普钠处理对种子发芽势和发芽率影响不显著,但显著增加了幼苗生物量、根长和苗高。可见,硝普钠处理可影响淀粉的转化和糖的积累,延缓总氮和萌动期前可溶性蛋白质的分解,加速游离氨基酸的积累,促进种子萌发和幼苗生长,其中以0.50 mmol/L 效果最佳。
圣女果; 硝普钠; 种子萌发; 幼苗; 碳氮代谢; 关键酶
圣女果(Lycopersiconesculentumvar.cerasiformeAlef.)又称樱桃小番茄,为茄科番茄属番茄栽培亚种中的一个变种,1年生草本,原产南美洲,因其栽培技术简单,适应性强,现在我国各省均有栽培。圣女果成熟果实含丰富的糖类、有机酸、矿物质、维生素等营养成分,尤其维生素C含量丰富,糖酸比适中,口感好,外形小巧,是颇有市场前景的“水果蔬菜”[1]。一氧化氮(NO)是生物体内的一种重要信号分子,广泛参与植物生长发育和各种逆境胁迫应答调控[2-3],如参与种子萌发、叶片扩张、根系生长、侧根形成、细胞程序性死亡以及植物抗逆反应等生理过程[4]。碳氮代谢是植物最基本的生理过程,其代谢强度及在植物生长发育过程中的动态变化直接影响着光合产物的形成、转化及矿质营养的吸收、蛋白质的合成等[5]。碳氮代谢对圣女果种子萌发、幼苗生长、产量、品质形成均有重大影响。因此,研究NO对圣女果碳氮代谢物质及其相关酶活性的影响,对促进碳氮物质的生产、运输和分配,促进种子的正常萌发和幼苗的健壮生长具有重要意义。
硝普钠作为NO供体已被广泛应用于逆境胁迫下提高种子萌发率及增强抗逆性,如硝普钠可提高盐胁迫下玉米[6]、燕麦[7]、紫苏[8]、高粱[9]、沙葱[10]、水飞蓟[11]种子的萌发率。外源硝普钠处理可提高干旱胁迫下葡萄叶片的可溶性蛋白质含量[12],促进板蓝根[13]和铁皮石斛[14]多糖的积累。此外,硝普钠处理可使黄瓜幼苗可溶性蛋白质含量提高,株高、茎粗和根长均有增加[15]。推测硝普钠对植物的碳氮代谢具有一定的调控作用,但该方面的研究至今还未见报道。鉴于此,以台湾圣女果品种朱云为试验材料,采用不同浓度的硝普钠处理后进行发芽试验,研究硝普钠对圣女果种子萌发至幼苗期碳氮代谢及相关酶活性的影响,旨在为利用硝普钠调控圣女果育苗提供理论依据。
1 材料和方法
1.1材料
选用圣女果品种朱云为试验材料,种子由云南省昆明市蔬菜种子公司提供。该品种极早熟,长势强,产量高,酸甜适中,栽培较广。
1.2试验设计
试验于2015年5月进行,设置3个硝普钠浓度处理,T1—T3分别为0.25、0.50、1.00 mmol/L,硝普钠浓度设置由预试验确定,同时以蒸馏水处理为对照(CK),每个处理重复4次。挑选粒大、饱满的种子,用蒸馏水浸泡2 h后放入垫有3层定性滤纸的培养皿中,每个培养皿中均匀放入100粒种子,然后分别倒入10 mL不同浓度的硝普钠处理液,置于白昼/黑夜温度为26 ℃/16 ℃的光照培养箱中进行培养,培养的昼/夜时间设置为16 h/8 h,白天光量子通量密度日均值约为100 μmol/(m2·s)。种子进入萌发期后,选择萌发健壮者播种于盛有育苗基质的营养钵(规格8 cm×8 cm)中。分别于种子吸胀期、萌动期、萌发期、幼苗形成期和幼苗期5个不同阶段随机选取20粒种子或幼苗测定碳氮营养物质含量及相关酶活性。试验重复3次。
1.3测定指标及方法
淀粉酶活性采用3,5-二硝基水杨酸比色法测定;蔗糖磷酸合成酶(SPS)活性和蔗糖合成酶(SS)活性使用南京建成生物工程研究所生产的试剂盒测定,以产生1 μmol/(mg·min)蔗糖为1个酶活单位(U);硝酸还原酶(NR)活性用磺胺比色法测定[16];谷氨酰胺合成酶(GS)活性和谷氨酸合成酶(GOGAT)活性按照王小纯等[17]的方法测定。
种子取样后于105 ℃烘箱中烘干粉碎后过孔径为0.074 mm的标准筛,筛下干粉末用于测定淀粉、可溶性糖、还原糖、蔗糖、总氮、可溶性蛋白质及游离氨基酸的含量。淀粉含量参照徐昌杰等[18]的方法测定,可溶性糖含量采用苯酚-硫酸比色法测定,还原糖含量采用3,5-二硝基比色法测定,蔗糖含量采用间苯二酚比色法测定,可溶性蛋白质含量采用考马斯亮蓝G250显色法测定,游离氨基酸含量采用酸性茚三酮比色法测定[16]。试材非结构性糖和淀粉中碳元素含量根据糖和淀粉分子中碳元素的比例进行换算而得。
种子进入萌发期时,测定种子发芽指标,发芽势=规定时间内发芽达到高峰期的种子发芽总数/供试种子数×100%;发芽率=规定时间内种子发芽总数/供试种子数×100%。该试验用2 d内发芽总数计算发芽势,用4 d内发芽总数计算发芽率。幼苗总生物量直接用天平(0.001 g)测量,幼苗根长和幼苗高用直尺测量。所有测定重复3次,取平均值。
1.4数据处理
数据采用SPSS 22.0软件进行Duncan氏差异显著性(P<0.05)分析,采用Excel 2010软件作图。
2 结果与分析
2.1硝普钠对圣女果种子萌发至幼苗期碳代谢的影响
2.1.1 淀粉和糖类含量 植物体内的糖类和淀粉含量可分别作为碳转化和积累的重要指标。从图1可以看出,圣女果种子萌发至幼苗期,淀粉含量逐渐下降,可溶性糖、还原糖和蔗糖含量则呈先上升后下降的趋势。吸胀期,T1、T2和T3处理的淀粉含量分别比CK高23.72%、37.25%和27.16%;从吸胀期到萌动期,含量急剧下降,之后缓慢下降;从吸胀期到幼苗期,T1、T2、T3处理和CK的淀粉含量分别下降了92.57%、93.89%、92.91%和90.26%。吸胀期,T1、T2处理和CK的可溶性糖含量分别比T3处理高107.40%、67.09%和57.73%;萌动期,T1、T2、T3处理和CK可溶性糖含量分别较吸胀期上升了13.97%、27.41%、42.99%和22.47%;而从萌动期到幼苗期,T1、T2、T3处理和CK分别下降了86.46%、81.33%、78.26%和80.04%。还原糖含量从吸胀期到萌动期呈急剧上升趋势,T1、T2、T3处理和CK分别上升了190.69%、332.42%、182.30%和116.83%;从萌发期到幼苗期则呈急剧下降的趋势,各处理分别下降了83.59%、81.92%、81.94%和80.56%。蔗糖含量从吸胀期到萌发期呈上升趋势,T1、T2、T3处理和CK分别上升了124.68%、144.24%、164.13%和81.73%,而从萌发期到幼苗期则呈下降趋势,分别下降了68.47%、74.08%、75.24%和75.87%。说明适当浓度的硝普钠处理能够促进淀粉的转化和糖的积累,其中0.50 mmol/L硝普钠处理对促进碳代谢效果最佳。
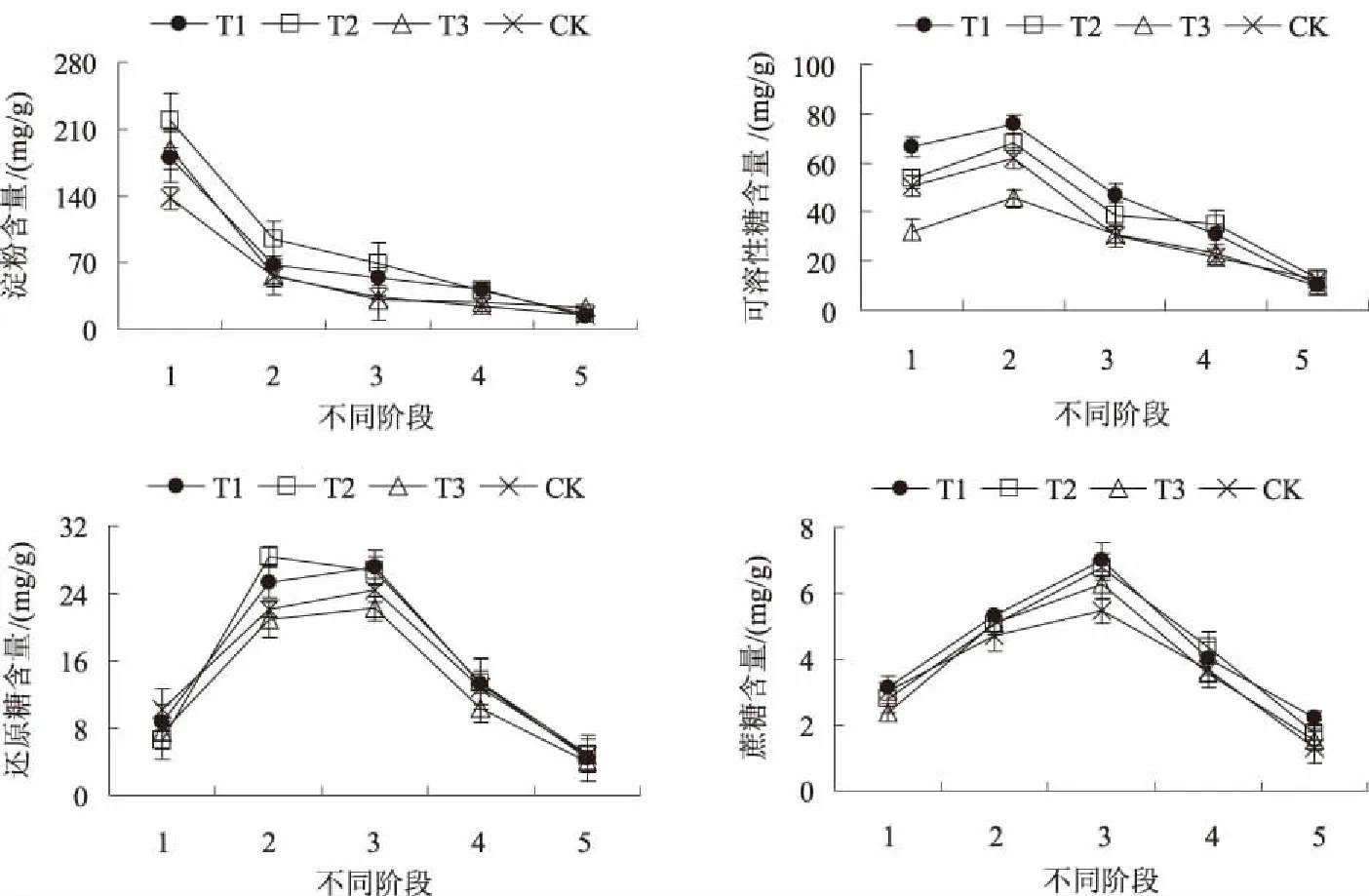
1.吸胀期; 2.萌动期; 3.萌发期; 4.幼苗形成期; 5.幼苗期。下同图1 硝普钠对圣女果种子萌发至幼苗期淀粉和糖类含量的影响
2.1.2 碳代谢关键酶活性 淀粉酶控制圣女果种子萌发至幼苗期淀粉的水解。SPS是控制蔗糖合成的关键酶,调节着光合产物在淀粉和蔗糖间的分配。SS存在于细胞质中,分解蔗糖生成腺苷二磷酸葡萄糖(UDPG)和果糖,用于合成淀粉等碳水化合物。图2显示,从吸胀期到萌动期,各处理淀粉酶活性变化不大,其大小依次为T2>T1>T3>CK;而从萌动期到幼苗期则呈大幅度下降趋势,T1、T2、T3处理和CK分别下降了84.67%、77.74%、84.49%和85.34%,且幼苗期T1、T3处理和CK分别比T2处理低34.33%、43.28%和53.73%。SPS活性呈下降趋势,吸胀期,T1、T2、T3处理分别比CK低25.21%、8.52%和2.53%;从吸胀期到幼苗期,T1、T2、T3处理和CK分别下降了89.87%、94.20%、95.18%和91.04%,且幼苗期T1、T2、T3处理分别比CK降低15.45%、40.79%、47.56%。T2、T3处理的SS活性从吸胀期到萌动期分别上升了24.43%、34.84%;从萌动期到幼苗形成期,则大幅度下降,T1、T2、T3处理和CK分别下降了90.87%、94.55%、97.22%和87.46%。可见,硝普钠对圣女果种子萌发至幼苗期的淀粉酶、SPS、SS活性产生较大影响。
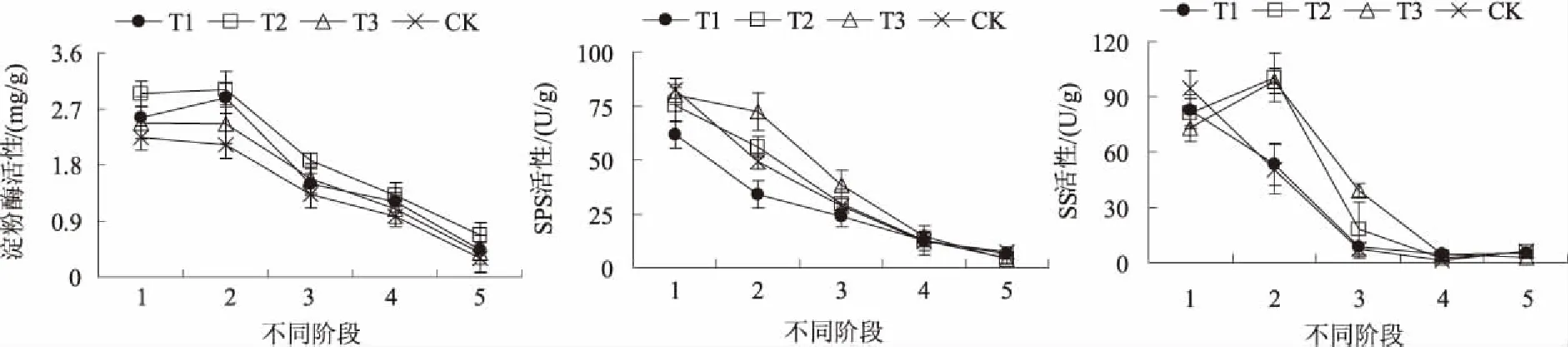
图2 硝普钠对圣女果种子萌发至幼苗期碳代谢关键酶活性的影响
2.2硝普钠对圣女果种子萌发至幼苗期氮代谢的影响
2.2.1 总氮、可溶性蛋白质和氨基酸含量 蛋白质是氮代谢的终产物,氨基酸是合成蛋白质的主要原料,也是蛋白质降解的主要产物。从图3可以看出,各处理的总氮含量均呈下降趋势,萌动期T1、T2、T3处理分别比CK低30.53%、27.67%、1.41%;从吸胀期到幼苗期,T1、T2、T3处理和CK分别下降了67.86%、61.50%、67.69%和84.17%。各处理可溶性蛋白质含量从吸胀期到萌动期呈上升趋势,萌动期T1、T2、T3处理分别比CK高17.76%、32.01%、13.08%;从萌动期到幼苗期呈下降趋势,T1、T2、T3处理及CK分别下降了80.75%、82.83%、81.61%及76.40%。T1、T2、T3处理和CK游离氨基酸含量总体呈上升趋势,CK在各阶段均最低,从吸胀期到幼苗期,T1、T2、T3处理和CK分别上升了60.07%、67.38%、57.46%和60.72%,幼苗期T1、T2、T3处理分别比CK高22.05%、32.84%、46.19%。可见,硝普钠处理能够延缓圣女果种子萌发到幼苗期间氮的降解,促进可溶性蛋白质在萌动期之前的积累和萌动期之后的转化,提高游离氨基酸含量。
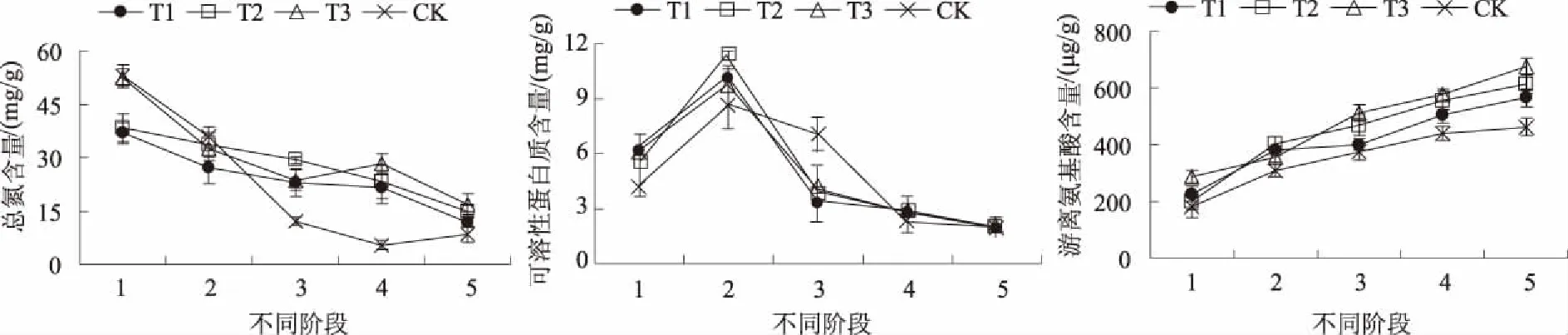
图3 硝普钠对圣女果种子萌发至幼苗期总氮、可溶性蛋白质和氨基酸含量的影响
2.2.2 氮代谢关键酶活性 NR是植物氮同化过程中关键的调节酶和限速酶,GS是催化氨同化的关键酶,GOGAT也是植物体内氮素同化的关键酶。由图4可看出,硝普钠处理后,NR和GOGAT活性均呈先上升后下降的趋势。NR活性的峰值出现在萌动期,此时T1、T2、T3处理分别比CK低16.68%、15.99%、6.60%;从萌动期到幼苗期,T1、T2、T3处理和CK分别下降了43.25%、71.26%、52.09%和80.69%,幼苗期T1、T2、T3处理分别比CK高294.93%、62.54%、204.36%。从吸胀期到幼苗期,GS活性总体呈下降趋势,吸胀期T1、T2、T3处理分别比CK低15.75%、48.47%、34.56%;幼苗期,T1、T2、T3处理及CK分别比吸胀期下降了72.23%、84.87%、84.11%及83.79%。GOGAT活性的峰值除CK在萌发期外,其余处理均在萌动期,此时T1、T2、T3处理分别比CK高8.88%、27.74%、16.77%,到幼苗期T1、T2、T3处理及CK分别比峰值时降低了61.78%、58.34%、64.39%及69.54%。可见,硝普钠处理对提高圣女果种子萌发至幼苗期的氮代谢产生了一定的影响。

图4 硝普钠对圣女果种子萌发至幼苗期氮代谢关键酶活性的影响
2.3硝普钠对圣女果种子萌发至幼苗期碳含量和碳氮比的影响
碳、氮含量是植物的重要营养指标,碳氮比则反映植物主要营养物质的偏向。图5所示,硝普钠处理可延缓圣女果种子萌发到幼苗期的碳损失。吸胀期T1、T2、T3处理的碳含量分别比CK高31.21%、45.58%、18.23%;幼苗期,T1、T2、T3处理及CK分别较吸胀期下降了90.50%、89.57%、84.93%及85.38%。CK的碳氮比在幼苗形成期出现峰值,此时比吸胀期上升了134.19%,之后便迅速下降;T1、T2、T3处理从吸胀期到幼苗期分别下降了70.45%、72.90%、53.35%。显然,硝普钠处理对圣女果种子萌发到幼苗期间的碳含量和碳氮比产生较大影响。
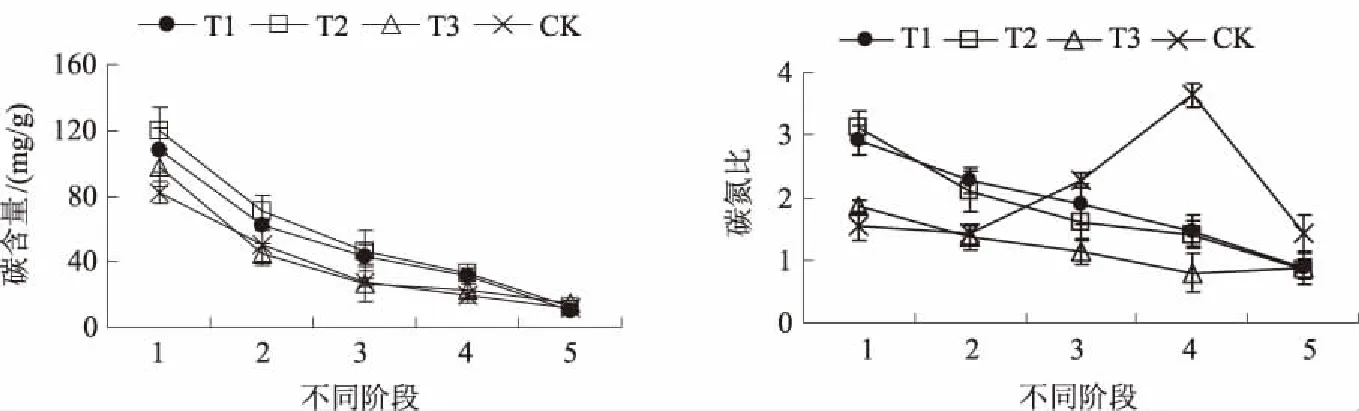
图5 硝普钠对圣女果种子萌发至幼苗期碳含量和碳氮比的影响
2.4硝普钠对圣女果种子萌发和幼苗生长指标的影响
从表1可以看出,硝普钠处理可在一定程度上提高圣女果种子发芽率和发芽势,但差异均不显著;处理后,幼苗生物量、根长和苗高均显著大于CK;其中,T1、T2、T3处理的生物量分别比CK高33.33%、100.00%、108.33%,幼苗根长分别比CK高26.49%、59.90%、56.44%,幼苗高分别比CK高17.35%、35.54%、43.39%。显然,硝普钠处理可促进圣女果幼苗的生长。
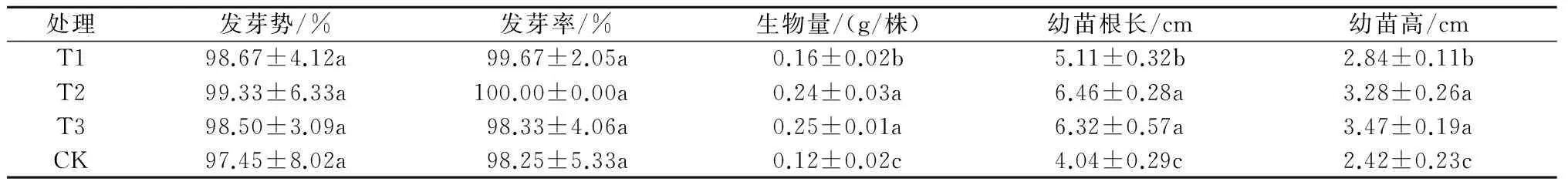
表1 硝普钠对圣女果种子萌发和幼苗生长指标的影响
注:同列不同字母表示在0.05水平上差异显著。
3 结论与讨论
植物的碳代谢包括碳的同化、碳水化合物的运输转化和碳的积累3个阶段。碳的同化是指无机碳在叶绿体中通过光合作用转化为有机碳;碳水化合物的运输转化是指磷酸丙糖通过叶绿体膜运送至细胞质合成蔗糖,并进一步转化为单糖;碳的积累主要是以蔗糖或淀粉的形式进行的。碳代谢的变化直接影响着碳水化合物的形成、转化及蛋白质的合成与分解[19-20]。研究表明,硝普钠[21]、糖[22]、氨基酸[23]等处理均能明显影响作物的碳氮代谢。本研究结果表明,圣女果种子萌发到幼苗期间,淀粉含量逐渐下降,可溶性糖、还原糖、蔗糖含量呈先升高后降低的趋势,硝普钠处理对其变化幅度产生较大影响。本研究还发现,硝普钠种子萌发到幼苗期间,淀粉酶活性在萌动期之前变化缓慢,之后急剧下降,原因是种子吸胀期到萌动期的淀粉含量较高,淀粉酶活性高有利于淀粉的分解,但经过一段时间以后,淀粉含量下降到一个较低水平,此时淀粉酶的活性也随之下降;硝普钠处理延缓淀粉酶活性的下降幅度,加速淀粉的分解。SPS和SS是促使作物蔗糖进入各种代谢途径的关键酶,一般认为SS主要起分解蔗糖的作用[24];SPS可催化蔗糖的合成,其活性与淀粉积累呈负相关,与蔗糖形成呈正相关。圣女果种子萌发到幼苗期,SPS和SS的活性总体呈逐渐下降的趋势,说明种子萌发期间淀粉的分解与蔗糖形成的速率均在减慢,碳代谢速率减缓,但与CK相比,0.50 mmol/L 硝普钠处理有效延缓了这2种酶活性的降低,一定程度上抑制了碳代谢的降低。
氮代谢包括硝态氮的还原、同化及有机含氮化合物的转化、合成等过程。植物吸收的无机态氮在植物体内大多数进入氨基酸,进一步合成蛋白质,少部分参与核酸等含氮物质的代谢。植物氮素同化的主要途径是经过硝酸盐还原为铵后直接参与氨基酸的合成与转化,在这期间,NR、GS和GOGAT等关键酶参与催化和调节。以氨基酸为主要底物在细胞中合成蛋白质,再对蛋白质进行修饰、分类、转运及储存等,成为植物有机体的组成部分,同时与植物的碳代谢协调统一。有研究报道,硝普钠处理植物可使可溶性蛋白质含量下降[12,15]。在本试验中发现,圣女果种子萌发到幼苗期间,总氮含量逐渐下降,GS活性也总体呈下降趋势,游离氨基酸含量逐渐上升,可溶性蛋白质含量及GOGAT、NR活性则呈先上升后下降的趋势,说明在此过程中,氮元素一方面会转化成为蛋白质,另一方面会发生损失;氨基酸含量上升是由于蛋白质的分解导致的,可溶性蛋白质含量到萌发期后下降是由于种子内贮藏的蛋白质逐渐转化为氨基酸供种子萌发和幼苗生长等生命活动所用,而幼苗体内还未合成蛋白质所致;硝普钠处理减缓了总氮的下降,加快了氨基酸含量的上升,在萌动期前加快了可溶性蛋白质含量的上升。可见,硝普钠处理能够促进圣女果种子萌发到幼苗期氮的同化和转化代谢。
碳氮代谢是作物最基本的代谢过程,对作物的生长发育、产量及品质的形成有较大影响。氮代谢需要依靠碳代谢提供碳源和能量,碳代谢则需要氮代谢提供所需的酶和光合色素。碳氮协调对作物的生长发育和产量形成至关重要[25]。研究发现,圣女果种子萌发到幼苗期间,总碳含量逐渐下降,除CK碳氮比呈先上升后下降的趋势外,硝普钠处理碳氮比稳定下降。此外,硝普钠处理能够促进圣女果种子萌发和幼苗的生长,这与郑春芳等[21]的研究结果一致。综上,硝普钠处理可促进圣女果种子萌发到幼苗期碳氮营养的转化,加速物质积累,促进种子萌发和幼苗的生长,其中以0.50 mmol/L处理效果最佳。
[1] 刘尚军,王若兰,姜海峰,等.可食膜果蔬保鲜效果研究[J].郑州工程学院学报,2004,25(4):58-61.
[2] Crawford N W,Guo F Q.New insights into nitric oxide metabolism and regulatory functions[J].Trends in Plant Science,2005,10(4):195-200.
[3] Besson-Bard A,Pugin A,Wendehenne D.New insights into nitric oxide signaling in plant[J].Annual Review of Plant Biology,2008,59:21-39.
[4] 张义凯,韩小娇,金洪,等.外源NO对铜胁迫下番茄光合、生物发光特性及矿质元素吸收的影响[J].植物营养与肥料学报,2010,73(1):172-178.
[5] 唐秀梅,钟瑞春,揭红科,等.间作花生对木薯碳氮代谢产物及关键酶活性的影响[J].中国农学通报,2011,27(3):94-98.
[6] 王芳,王汉宁.外源NO对NaCl胁迫下玉米幼苗氧化损伤的保护作用[J].草地学报,2012,20(6):1117-1122.
[7] 苏桐,龙瑞军,魏小红,等.外源NO对NaCl胁迫下燕麦幼苗氧化损伤的保护作用[J].草业学报,2008,17(5):48-53.
[8] 张春平,何平,杜丹丹,等.外源NO供体硝普钠(SNP)对盐胁迫下紫苏种子萌发及幼苗抗氧化酶活性的影响[J].中药材,2011,34(5):665-669.
[9] 石瑞,杨丽丽,刘树楠,等.外源NO对NaCl胁迫下高粱幼苗生理响应的调节[J].江苏农业科学,2016,44(8):139-142.
[10] 陆莹,王萍,宋兆伟.外源一氧化氮浸种对NaCl胁迫下沙葱种子萌发和生理特性的影响[J].华北农学报,2011,26(2):207-212.
[11] 张晓倩,王康才,张彦南,等.外源NO对NaCl胁迫下水飞蓟种子萌发和幼苗生理特性的影响[J].中草药,2013,44(22):3216-3222.
[12] 张永福,何田方,牛燕芬,等.硝普钠对干旱胁迫下葡萄抗旱生理的影响[J].广东农业科学,2014,41(24):40-44.
[13] 毛亚斌,魏小红.外源NO对干旱胁迫下板蓝根叶片氧化损伤的保护作用[J].草业科学,2010,27(6):97-101.
[14] 蒋天仪,卓宇,唐敏,等.外源一氧化氮(NO)对铁皮石斛类原球茎生长及多糖积累的影响[J].江苏农业科学,2016,44(4):257-260.
[15] 杜桌涛,商桑,朱白婢,等.外源NO对低温胁迫下苦瓜幼苗生长和几个生理指标的影响[J].热带作物学报,2016,37(3):482-487.
[16] 王学奎.植物生理生化实验原理与技术[M].北京:高等教育出版社,2006.
[17] 王小纯,熊淑萍,马新明,等.不同形态氮素对专用型小麦花后氮代谢关键酶活性及籽粒蛋白质含量的影响[J].生态学报,2005,25(4):802-807.
[18] 徐昌杰,陈文峻,陈昆松,等.淀粉含量测定的一种简便方法——碘显色法[J].生物技术,1998,8(2):41-43.
[19] 史宏志,韩锦峰.烤烟碳氮代谢几个问题的探讨[J].烟草科技,1998(2):34-36.
[20] 李志,史宏志,刘国顺,等.施氮量对皖南砂壤土烤烟碳氮代谢动态变化的影响[J].土壤,2010,42(1):8-13.
[21] 郑春芳,姜东,戴廷波,等.外源一氧化氮供体硝普钠浸种对盐胁迫下小麦幼苗碳氮代谢及抗氧化系统的影响[J].生态学报,2010,30(5):1174-1183.
[22] 冯艳,王富国,张相松,等.外源糖调节不同C/N比对苋菜幼苗碳、氮代谢的影响[J].青岛农业大学学报(自然科学版),2008,25(4):272-275.
[23] 曹小闯,吴良欢,马庆旭,等.高等植物对氨基酸态氮的吸收与利用研究进展[J].应用生态学报,2015,26(3):919-929.
[24] Elling L.Effect of metal ions on sucrose synthase from rice grains-a study on enzyme inhibition and enzyme topography[J].Glycobiology,1995,5(2):201-206.
[25] Uhart S A,Andrade F H.Nitrogen deficiency in maize:Ⅱ.Car-bon-nitrogen interaction effects on kernel number and grain yield[J].Crop Science,1995,35:1384-1389.
Effect of Sodium Nitroprussiate on the Metabolites and Key Enzyme Activities of Carbon-Nitrogen Metabolism from Seed Germination to Seedling Period of Cherry Tomatoes
ZHANG Yongfu1,2,GUO Lihong1,2,JIANG Shuping1,2,JIN Song1,2,YAO Liyuan1,2,ZHAO Mingfang1,2
(1.School of Agriculture,Kunming University,Kunming 650214,China; 2.Key Laboratory of Special Biological Resource Development and Utilization of Universities in Yunnan Province,Kunming 650214,China)
The purpose of this paper was to elucidate the regulating mechanism of sodium nitroprussiate on seed germination and seedling growth of cherry tomatoes.After treated by different concentration of sodium nitroprusside,the effect on the metabolites and key enzyme activities of carbon-nitrogen metabolism of cherry tomatoes cultivar Zhuyun as materials were investigated.The results showed that from seed germination to seedling period,the contents of starch,total nitrogen decreased,but the contents of soluble sugar,reducing sugar,sucrose and soluble protein increased firstly and then decreased;free amino acids content increased gradually.Sodium nitroprusside made the contents of starch,sucrose and free amino acids higher than CK.Seed germination to seedling stage in cherry tomatoes,the activities of amylase,glutamine synthetase(GS)and sucrose phosphate synthase(SPS)decreased;the activities of nitrate reductase(NR)and glutamate synthase(GOGAT)were first increased and then decreased;CK sucrose synthase(SS) activity decreased.The sucrose synthase(SS) activity decreased with the treatment of CK and 0.25 mmol/L sodium nitroprusside,and the trend was first increased and then decreased after treatment at 0.50,1.00 mmol/L sodium nitroprusside.Sodium nitroprusside treatment increased amylase activity;carbon-nitrogen ratio of CK increased first and then decreased,while the others tended to decrease.In addition,sodium nitroprusside on seed germination potential and germination rate were not affected,but significantly increased biomass accumulation,root length and height of the seedling.These data suggest that sodium nitroprusside can influence the conversion of starch and sugar accumulation,delay the decomposition of total nitrogen and soluble protein,accelerate the accumulation of free amino acids,promote the seed germination and seedling growth,of which 0.50 mmol/L has the best effect.
cherry tomatoes; sodium nitroprusside; seed germination; seedling; carbon and nitrogen metabolism; key enzyme
2017-03-15
国家自然科学基金项目(31660559);昆明学院科学研究项目(XJZZ1604);云南省高校特色生物资源开发与利用重点实验室开放基金项目(GXKJ201615)
张永福(1981-),男,云南弥勒人,副教授,博士,主要从事园艺植物抗性生理研究。E-mail:123017360@qq.com
S641.2
: A
: 1004-3268(2017)09-0098-06
猜你喜欢
今日农业(2021年13期)2021-11-26
小猕猴学习画刊(2021年12期)2021-01-10
今日农业(2020年22期)2020-12-14
新农民(2020年30期)2020-12-09
中国甜菜糖业(2020年3期)2020-12-08
浙江工业大学学报(2017年5期)2018-01-22
中国糖料(2016年1期)2016-12-01
中国卫生标准管理(2015年1期)2016-01-14
中国卫生标准管理(2015年18期)2015-01-26
中国卫生标准管理(2015年17期)2015-01-26
