
渤海鱼类的食物关系*
2018-06-19 03:00张波
渔业科学进展 2018年3期
张 波
(1. 农业部海洋渔业可持续发展重点实验室 山东省渔业资源与生态环境重点实验室中国水产科学研究院黄海水产研究所 青岛 266071;2. 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
渤海作为我国的内海,是黄、渤海主要渔业种类的产卵场和索饵场,也是我国海洋渔业生产的重要渔场。从20世纪50年代以来,我国长期在渤海进行渔业资源与环境调查、监测,开展了大量科学研究,对渤海鱼类种类组成、资源结构和数量分布的变动特征和规律有了比较全面系统的了解(邓景耀等, 1988;金显仕等, 1998、2001; Tang et al, 2003; 单秀娟等, 2012;李忠义等, 2017)。对渤海鱼类的食物关系也开展了较多研究,邓景耀等(1986、1997)对 20世纪 80年代和90年代渤海鱼类食物关系进行了研究;李军(1990)对渤海重要食物主线蓝点马鲛(Scomberomorus niphonius)食物链结构进行了研究;张波等(2012)对渤海鱼类群落的营养功能群进行了研究;Zhang等(2007)、许思思等(2014)和林群等(2016)的研究均发现渤海渔获物的营养级呈下降趋势。本研究拟通过分析渤海鱼类的食物组成、食性类型、饵料的生境宽度和营养级,了解当前渤海鱼类的食物关系及其变化,为进一步构建渤海生态系统模型,探讨环境变化对渔业种群动态的耦合关系及其资源效应提供基础资料。
1 材料与方法
2010~2011年用双拖渔船对渤海进行5次大面综合调查,取样站位如图 1所示。调查网具网口高度6 m,网口宽度22.6 m,网目周长1740目,网目63 mm,囊网网目20 mm。定点站位拖网1 h,拖速为3.0 kn。共收集了27种鱼类10156个胃含物样品(表1)。取样个体经生物学测定后,取出消化道速冻保存。胃含物分析时,将其解冻用吸水纸吸去水分后,再在双筒解剖镜下鉴定饵料生物的种类并分别计数和称重,食物重量精确到0.001 g,并尽量鉴定到最低分类单元。
根据胃含物分析结果,综合饵料生物的重量百分比(W%)、个数百分比(N%)和出现频率(F%)来评价摄食的各种饵料种类的重要性。采用营养级来评价各鱼种在渤海生态系统的营养生态位,采用 Shannon-Wiener多样性指数(H′)来评价各鱼种的饵料生境宽度。为了使各种鱼的Shannon-Wiener多样性指数具有可比性,将各种鱼的食物组成归为以下饵料类群:浮游植物、桡足类、磷虾类、毛虾类、糠虾类、虫戎 类、甲壳类幼体、底层虾类、蟹类、口足类、涟虫类、蛇尾类、腹足类、瓣鳃类、多毛类、端足类、头足类和鱼类后计算 Shannon-Wiener多样性指数。由于各鱼种摄食的饵料生物个体差异较大,因此本研究选用饵料的出现频率百分比组成 FO%来计算各鱼种的营养级(TL)和Shannon-Wiener多样性指数。

图1 取样站位Fig. 1 Sampling stations

表1 渤海鱼类的胃含物样品Tab.1 Stomach samples of fish species in the Bohai Sea

式中Fi%为各饵料的出现频率;FOi%为各饵料的出现频率百分比组成;TLi为各饵料的营养级。饵料类群的营养级参考张波等(2004)和程济生等(1997)。根据邓景耀等(1986、1997)将鱼类按营养级划分为低营养级鱼类(营养级≤3.8)、中营养级鱼类(营养级3.9~4.4)和高营养级鱼类(营养级≥4.5)。
2 结果
2.1 各种鱼类的食物组成
胃含物分析结果(表 2)显示:青鳞沙丁鱼和斑摄食植物性和动物性饵料,均属杂食性鱼类。摄食的植物性饵料以圆筛藻(Cosinodiscus sp.)为主;摄食的动物性饵料,青鳞沙丁鱼以桡足类为主,斑以瓣鳃类和腹足类为主。赤鼻棱鳀、黄鲫和小带鱼均以摄食浮游动物饵料为主,兼食一定比例的底层虾类。其中赤鼻棱鳀共摄食12类饵料,浮游动物饵料以桡足类和磷虾类为主;黄鲫以毛虾类、糠虾类、磷虾类和甲壳类幼体为主;小带鱼以磷虾类、甲壳类幼体和毛虾类为主。方氏云鳚则兼食浮游动物、底栖动物和底层虾类饵料。这6种鱼所摄食的主要饵料种类有太平洋磷虾(Euphausia pacifica)、中国毛虾(Acetes chinensis)、小拟哲水蚤(Paracalanus parvus)、中华哲水蚤 (Calanus sinicus)和细螯虾(Leptochela gracilis)。
胃含物分析结果(表 3)显示:这 6种鱼均主要以底栖动物为食,长绵鳚主要摄食瓣鳃类和端足类,其次是虾蟹类;绯主要摄食端足类,其次是多毛类、瓣鳃类和腹足类;矛尾虾虎鱼摄食的食物范围较广,共12类饵料;六丝矛尾虾虎鱼主要摄食底层虾类、瓣鳃类和端足类;长吻红舌鳎和短吻红舌鳎均主要摄食端足类和瓣鳃类,其次是多毛类和底层虾类。饵料中日本鼓虾(Alpheus japonicus)和细螯虾是主要的底层虾类饵料。
胃含物分析结果(表 4)显示:3种石首鱼类中小黄鱼摄食的食物范围较广,共9类饵料,以桡足类、糠虾类、底层虾类和鱼类为主;白姑鱼以底层虾类和鱼类为主,其次是口足类和头足类;皮氏叫姑鱼则以底层虾类为主,其次是多毛类和鱼类。细纹狮子鱼以底层虾类为主,其次是鱼类。大泷六线鱼摄食的食物范围较广,共9类饵料,其中多毛类、底层虾类和口足类是主要的饵料。绒杜父鱼主要摄食鱼类饵料。饵料中日本鼓虾、葛氏长臂虾(Palaemon gravieri)、脊腹褐虾(Crangon affinis)和细螯虾是主要的底层虾类饵料,六丝矛尾虾虎鱼和矛尾虾虎鱼是主要的鱼类饵料。
胃含物分析结果(表5)显示:这 3种鱼均以底层虾类和鱼类为主要饵料,其中褐菖鲉摄食的食物范围较广,共10类饵料。饵料中日本鼓虾和葛氏长臂虾是主要的底层虾类饵料,六丝矛尾虾虎鱼、矛尾虾虎鱼、皮氏叫姑鱼和小黄鱼是主要的鱼类饵料。
胃含物分析结果(表6)表明,这 6种鱼除鲬主要摄食底层虾类、口足类和鱼类外,其余5种均主要摄食鱼类饵料,其中油还摄食较多头足类饵料,蓝点马鲛还摄食较多毛虾类和口足类饵料。饵料中六丝矛尾虾虎鱼、矛尾虾虎鱼、小黄鱼、皮氏叫姑鱼和黄鲫是主要的鱼类饵料。
2.2 各种鱼类的营养级和饵料生境宽度
本研究中渤海的27种鱼类的平均营养级为3.88,包括了12种低营养级鱼类,12种中营养级鱼类和3种高营养级鱼类(表7)。其中12种低营养级鱼类平均营养级为3.36,包括了杂食性鱼类(斑、青鳞沙丁鱼),主要摄食浮游动物的鱼类(包括赤鼻棱鳀、黄鲫和小带鱼),主要摄食底栖动物的鱼类(包括、短吻红舌鳎、长吻红舌鳎、长绵鳚、矛尾虾虎鱼和六丝矛尾虾虎鱼)和主要兼食浮游动物和底栖动物的鱼类(方氏云鳚)。12种中营养级鱼类平均营养级为4.25,其中的8种(包括白姑鱼、皮氏叫姑鱼、细纹狮子鱼、绒杜父鱼、花鲈、鲬、许氏平鲉、绿鳍鱼)主要兼食底栖动物(以底层虾类为主)和鱼类饵料,4种(包括小黄鱼、大泷六线鱼、褐菖鲉和蓝点马鲛)主要兼食浮游动物、底栖动物和鱼类饵料。3种高营养级鱼类(长蛇鲻、黄和油)平均营养级为4.51,主要摄食鱼类饵料。从饵料生境宽度看,高营养级鱼类、中营养级鱼类和低营养级鱼类的平均饵料生境宽度值分别为 0.34、1.16和1.47。3种高营养级鱼类饵料生境宽度值均很低,属于狭食性鱼类;中营养级的小黄鱼和低营养级的矛尾虾虎鱼是渤海饵料生境宽度最大的2种鱼。

?

?

?
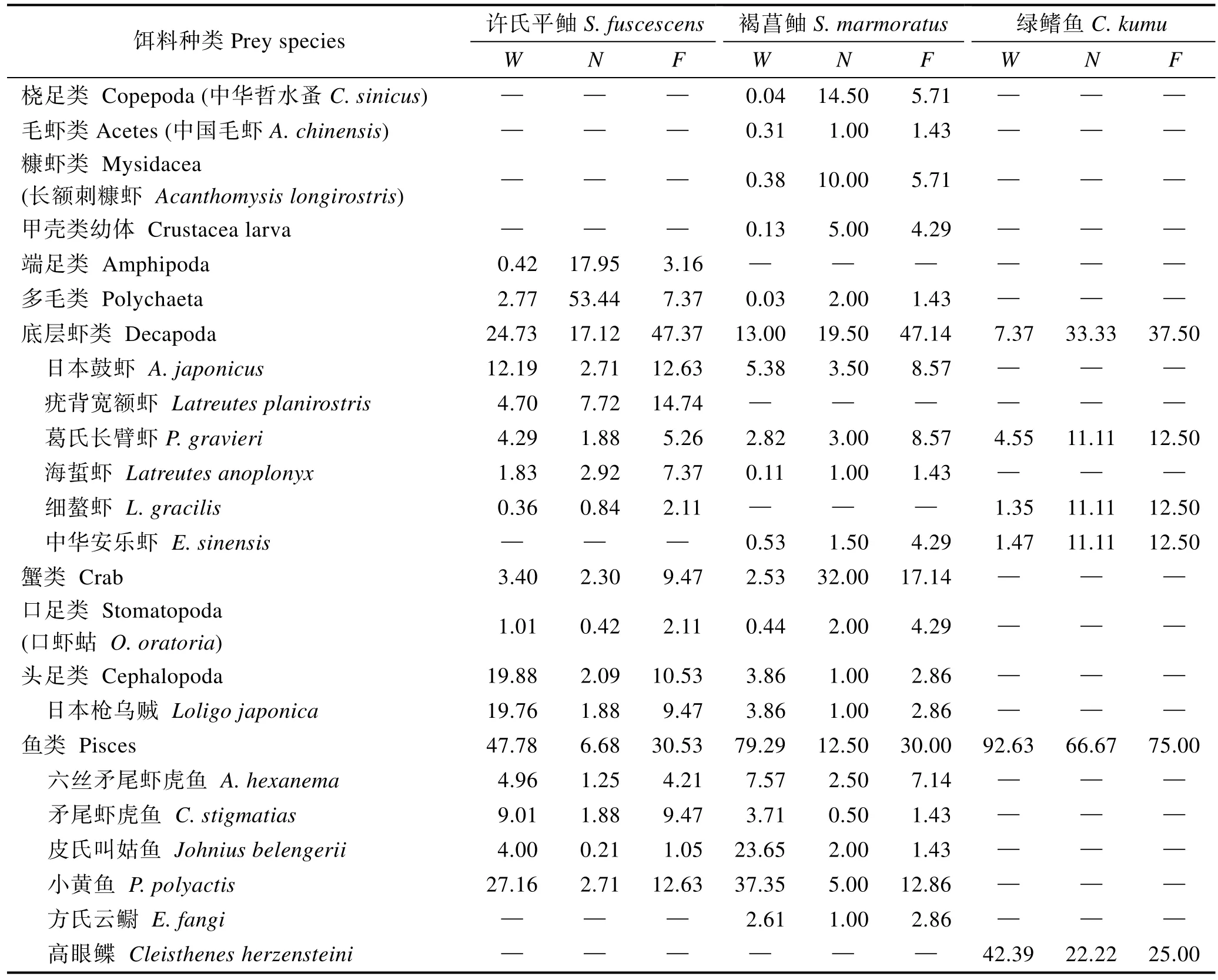
表6 许氏平鲉、褐菖鲉和绿鳍鱼的食物组成Tab.6 Diet composition of S. fuscescens, S. marmoratus and C. kumu (%)

表7 渤海各鱼种的营养级和饵料生境宽度Tab.7 Trophic level and dietary breadth of each fish in the Bohai Sea
3 讨论
食物联系是海洋生态系统结构与功能的基本表达形式,能量通过食物链-食物网转化为各营养层次生物生产力,形成生态系统生物资源产量,并对生态系统的服务和产出及其动态产生影响(唐启升, 1999)。海洋生态系统对物理、化学过程的响应常常表现为食物网的变化,而且海洋食物网又直接与生态系统的多样性、脆弱性和生产力密切相关。当前,在人类活动(富营养化、资源的过度开发利用等)与气候变化(全球变暖、海洋酸化等)相互叠加产生的多重压力下,近海生态系统发生了显著的变化,并严重影响了近海生态系统的食物产出功能。因此,对海洋生态系统的食物联系进行长期、系统的监测和研究是非常必要和有意义的。近20年来,随着我国海洋生态系统动力学研究的启动,先后开展了国家自然科学基金重大项目“渤海生态系统动力学与生物资源持续利用”以及国家重点基础研究发展计划(973计划) “东、黄海生态系统动力学与生物资源可持续利用”、“我国近海生态系统食物产出的关键过程及其可持续机理” 和“多重压力下近海生态系统可持续产出与适应性管理的科学基础”等项目,均将海洋食物网研究作为开展海洋可持续生态系统整合研究的重要切入口,在全程食物网研究中进一步突出以食物网关键种构成的食物产出主线,如“硅藻类群-中华哲水蚤-鳀-蓝点马鲛”,并在各营养层次的层面上展开研究(唐启升等, 2005)。
个体水平上鱼类摄食生态的研究是食物网研究的基础,评价一种饵料生物重要性的常用指标,如饵料的重量百分比、个数百分比、出现频率、出现频率百分比组成、相对重要性指标和相对重要性百分比都有一定的局限性(窦硕增, 1996; 薛莹等, 2003),不同的研究者选用了不同指标开展分析研究。张其永等(1983)研究二长棘鲷的摄食习性时,认为由于其饵料生物个体大小差异悬殊,饵料的个数百分比不适合;个数百分比适用于摄食的饵料生物个体差异不大的鱼类食性研究。郭斌等(2010)在分别以饵料的重量百分比、个数百分比、出现频率百分比组成和相对重要性百分比为指标进行聚类分析来研究小黄鱼幼鱼摄食习性随体长的变化时,发现使用个数百分比和出现频率百分比组成进行聚类分析能够更加准确地描述小黄鱼摄食习性的转变特点;而使用重量百分比和相对重要性百分比则会掩盖了以数量多、质量小为特点的饵料生物对于小黄鱼幼鱼食物组成的重要性。由于各鱼种摄食的饵料生物个体差异较大,因此本研究综合饵料生物的重量百分比、个数百分比和出现频率来评价各鱼种的摄食习性,在计算各鱼种的营养级和Shannon-Wiener多样性指数时选用了饵料的出现频率百分比组成这一指标。
营养级分析结果表明,本研究中渤海的27种鱼类有12种低营养级鱼类、12种中营养级鱼类和3种高营养级鱼类;总体来看,营养级越高,饵料生境宽度变小。3种高营养级鱼类均为鱼食性鱼类,饵料生境宽度值很低,属于狭食性鱼类。矛尾虾虎鱼和小黄鱼是当前渤海生态系统食物网中最重要的 2种饵料种类,同时也是摄食饵料生境宽度最大的2种鱼,均摄食浮游动物、底栖生物和游泳动物3大类饵料,食物网中重要环节的广食性有利于食物网各营养层次的物质、能量流动。Zhang等(2007)的研究表明,种类组成的变化,个体的小型化,以及摄食食物种类的变化是导致海洋生态系统营养级波动的主要原因,针对渤海而言,采用不同的研究方法均发现渤海渔获物的平均营养级呈下降趋势(Zhang et al, 2007; 许思思等, 2014; 林群等2016),从各鱼种营养级分析,渤海1992~1993年中、高营养级鱼类的营养级较1982~1983年有所下降(邓景耀等, 1997),而本研究表明,当前渤海各鱼种营养级较1992~1993年变化不大。可见,种类组成的变化是渤海当前生态系统营养级波动的主要原因。
从各鱼种饵料组成分析表明,渤海的27种鱼类有杂食性鱼类、浮游动物食性鱼类、底栖动物食性鱼类、混合动物食性鱼类和鱼食性鱼类;主要饵料种类有浮游动物的太平洋磷虾、中国毛虾、小拟哲水蚤、中华哲水蚤;底栖动物的多毛类、瓣鳃类和腹足类;底层虾类的口虾蛄、日本鼓虾、葛氏长臂虾、脊腹褐虾和细螯虾;鱼类的六丝矛尾虾虎鱼、矛尾虾虎鱼、小黄鱼、皮氏叫姑鱼和黄鲫。1982~1983年渤海鱼类食物网中的几个主要环节是浮游动物、鼓虾、矛尾虾虎鱼、六丝矛尾虾虎鱼、鳀、短尾类和软体动物,其中鼓虾是鳎类、鳐类和石首鱼类摄食竞争对象,但因其资源量大,所以对这些鱼类的摄食限制作用并不大(邓景耀等, 1986)。1992~1993年渤海食物网与1982~1983年相比,小黄鱼、白姑鱼摄食鳀的比例增加;长蛇鲻摄食的鳀取代了青鳞鱼;花鲈摄食的石首鱼科鱼类取代了青鳞鱼;蓝点马鲛摄食的毛虾和口虾蛄取代了鳀和小型虾虎鱼。蓝点马鲛、小黄鱼、油和长蛇鲻之间的饵料重叠系数较高与共同捕食鳀有关。其中数量占优势的鳀处于渤海鱼类食物网的重要环节,它是多种大型经济鱼类如蓝点马鲛、长蛇鲻、油、鲬,以及小黄鱼和黄姑鱼为代表的石首科鱼类及枪乌贼的重要饵料。与1982~1983年相比,主要中高级肉食性鱼类摄食鳀的比例增加,这主要是由于鳀资源量增加所致。渤海鱼类饵料重叠的主要对象已由1982~1983年的鼓虾类和虾虎鱼类发展为鳀(邓景耀等,1997)。而本研究中渤海鱼类摄食的头足类饵料减少,如小黄鱼、大泷六线鱼、细纹狮子鱼等;27种鱼中仅小黄鱼、黄、蓝点马鲛和鲬摄食少量的鳀,小黄鱼、白姑鱼和长蛇鲻摄食的虾虎鱼类又取代了鳀;蓝点马鲛除了摄食较多的毛虾和口虾蛄以外,还摄食较多的小黄鱼。
20世纪 80年代渤海鱼类群落占主要地位的有黄鲫、黑鳃梅童、鳀、孔鳐、小黄鱼、花鲈、长绵鳚、蓝点马鲛、焦氏舌鳎和黄盖鲽(邓景耀等, 1988)。邓景耀等(1986)将渤海鱼类分为5个捕食类群:(1)以黄盖鲽为代表,主食腔肠动物、多毛类和棘皮动物;(2)以鳎类为代表,主食鼓虾和短尾类;(3)以鳐类、石首鱼类为代表,主食鼓虾和虾虎鱼;(4)以鳀为代表,主食浮游动物;(5)以牙鲆为代表,主食鱼类。此时渤海食物网的主要食物链为“植物、有机碎屑→鼓虾→鱼类”,“小型底栖动物→虾虎鱼→大型经济鱼类”和“浮游动物→鳀→大型肉食性鱼类”。20世纪 90年代鳀资源密度迅速增加,成为渤海鱼类资源最丰富的种类(金显仕等, 1998),渤海食物网中主要食物链转变为“浮游动物→鳀→大型肉食性鱼类”(邓景耀等,1997)。渤海的鳀资源密度在 1992~1993年达到最高峰后迅速下降,2010年春季调查未发现鳀,夏季也仅0.13 kg/h (金显仕, 2014),当前渤海食物网“浮游动物→鳀→大型肉食性鱼类”的食物链基本被破坏。同时20世纪80年代时渤海鱼类5个捕食类群的代表种资源量衰退,当前渤海食物网主要食物链转变为“植物、有机碎屑→鼓虾→鱼类”和“底栖动物→虾虎鱼、小黄鱼→大型经济鱼类”。浮游食物链的削弱不仅不利于渤海生态系统的有效固碳,还不利于食物网的物质、能量流动,从而影响生态系统的生产力(张波等, 2015)。因此,当前渤海全程食物网以及突出以食物网关键种构成的食物产出主线的研究也应做出相应的改变。同时,进一步的研究需要应用如海洋食物网拓扑学方法(朱江峰等, 2016)等新方法,定量描述食物网各成员间关系,以及各成员对整个食物网的作用。
Cheng JS, Zhu JS. Study on the feeding habit and trophic level of main economic invertebrates in the Huanghai Sea. Acta Oceanologica Sinica, 1997, 19(6): 102–108 [程济生, 朱金声. 黄海主要经济无脊椎动物摄食特征及其营养层次的研究. 海洋学报, 1997, 19(6): 102–108]
Deng JY, Meng TX, Ren SM. Food web of fishes in Bohai Sea.Acta Ecologica Sinica, 1986, 6(4): 70–78 [邓景耀, 孟田湘,任胜民. 渤海鱼类食物关系的初步研究. 生态学报, 1986,6(4): 70–78]
Deng JY, Meng TX, Ren SM, et al. Species composition,abundance and distribution of fishes in the Bohai Sea.Marine Fisheries Research, 1988(9): 11–90 [邓景耀, 孟田湘, 任胜民, 等. 渤海鱼类种类组成及数量分布. 海洋水产研究, 1988(9): 11–90]
Deng JY, Jiang WM, Yang JM, et al. Species interaction and food web of major predatory species in the Bohai Sea.Journal of Fishery Sciences of China, 1997(4): 1–7 [邓景耀,姜卫民, 杨纪明, 等. 渤海主要生物种间关系及食物网研究. 中国水产科学, 1997(4): 1–7]
Dou SZ. Feeding ecology of fish—acritical review on theories,methods and their application. Oceanologia et Limnologia Sinica, 1996, 27(5): 556–561 [窦硕增. 鱼类摄食生态研究的理论及方法. 海洋与湖沼, 1996, 27(5): 556–561]
Guo B, Zhang B, Jin XS. Diet composition and ontogenetic variation in feeding habits of juvenile small yellow croaker Pseudosciaena polyactis Bleeker in the Yellow Sea. Journal of Fishery Sciences of China, 2010, 17(2): 289–297 [郭斌,张波, 金显仕. 黄海海州湾小黄鱼幼鱼的食性及其随体长的变化. 中国水产科学, 2010, 17(2): 289–297]
Jin XS, Tang QS. The structure, distribution and variation of the fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 1998(3): 18–24 [金显仕, 唐启升. 渤海渔业资源结构、数量分布及其变化. 中国水产科学,1998(3): 18–24]
Jin XS. The dynamics of major fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 2001, 7(4):22–26 [金显仕. 渤海主要渔业生物资源变动的研究. 中国水产科学, 2001, 7(4): 22–26]
Jin XS. The foundation and prospect of stock enhancement of fishery resources in the Bohai Sea and Yellow Sea. Beijing:Science Press, 2014 [金显仕. 黄渤海渔业资源增殖基础与前景. 北京: 科学出版社, 2014]
Li J. The food chain structure of Japanese Spanish mackerel in the Bohai Sea. Studia Marina Sinica, 1990, 31: 93–107 [李军. 渤海蓝点马鲛食物链结构的研究. 海洋科学集刊,1990, 31: 93–107]
Lin Q, Wang J, Yan W, et al. Effects of fishing and environmental change on the ecosystem of the Bohai Sea.Journal of Fishery Sciences of China, 2016, 23(3): 619–629[林群, 王俊, 袁伟, 等. 捕捞和环境变化对渤海生态系统的影响. 中国水产科学, 2016, 23(3): 619–629]
Li ZY, Wu Q, Shan XJ, et al. Interannual variations in fish community structure in the Bohai Sea. Journal of Fishery Sciences of China, 2017, 24(2): 403–413 [李忠义, 吴强,单秀娟, 等. 渤海鱼类群落结构的年际变化. 中国水产科学, 2017, 24(2): 403–413]
Shan XJ, Jin XS, Li ZY, et al. Fish community structure and stock dynamics of main releasing fish species in the Bohai Sea. Progress in Fishery Sciences, 2012, 33(6): 1–9 [单秀娟,金显仕, 李忠义, 等. 渤海鱼类群落结构及其主要增殖放流鱼类的资源量变化. 渔业科学进展, 2012, 33(6): 1–9]
Tang QS. Strategies of research on marine food web and trophodynamics between high trophic levels. Marine Fisheries Research, 1999, 20(2): 1–6 [唐启升. 海洋食物网与高营养层次营养动力学研究策略. 海洋水产研究, 1999,20(2): 1–6]
Tang Q, Jin X, Wang J, et al. Decadal-scale variation of ecosystem productivity and control mechanisms in the Bohai Sea. Fisheries Oceanography, 2003, 12(4/5): 223–233
Tang QS, Su JL, Zhang J. Key processes and sustainable mechanisms of ecosystem food production in the coastal ocean of China. Advances in Earth Science, 2005, 20(12):1280–1287 [唐启升, 苏纪兰, 张经. 我国近海生态系统食物产出的关键过程及其可持续机理. 地球科学进展, 2005,20(12): 1280–1287]
Xue Y, Jin XS. Review of the study on feeding habits of fishes and food webs. Marine Fisheries Research, 2003, 24(2):76–87 [薛莹, 金显仕. 鱼类食性和食物网研究评述. 海洋水产研究, 2003, 24(2): 76–87]
Xu SS, Song JM, Li XG, et al. Variation characteristics of catch structure in the Bohai Sea and its influencing factors.Journal of Natural Resources, 2014, 29(3): 500–506 [许思思,宋金明, 李学刚, 等. 渤海渔获物资源结构的变化特征及其影响因素分析. 自然资源学报, 2014, 29(3): 500–506]
Zhang QY, Zhang YZ. Study on food habits of Parargyrops edita in Minnan-Taiwan shoal. Acta Oceanologica Sinica, 1983,5(3): 349–362 [张其永, 张雅芝. 闽南-台湾浅滩二长棘鲷食性研究. 海洋学报, 1983, 5(3): 349–362]
Zhang B, Tang QS. Study on trophic level of important resources species at high trophic levels in the Bohai Sea, Yellow Sea and East China Sea. Advances in Marine Science, 2004,22(4): 393–404 [张波, 唐启升. 渤、黄、东海高营养层次重要生物资源种类的营养级研究. 海洋科学进展, 2004,22(4): 393–404]
Zhang B, Tang Q, Jin X. Decadal-scale variations of trophic levels at high trophic levels in the Yellow Sea and Bohai Sea ecosystem. Journal of Marine System, 2007, 67(3–4):304–311
Zhang B, Li ZY, Jin XS. Functional groups of fish assemblages and their major species in the Bohai Sea. Journal of Fisheries of China, 2012, 36(1): 64–72 [张波, 李忠义, 金显仕. 渤海鱼类群落功能群及其主要种类. 水产学报,2012, 36(1): 64–72]
Zhang B, Wu Q, Jin XS. Interannual variation in the food web of commercially harvested species in Laizhou Bay from 1959 to 2011. Journal of Fishery Sciences of China, 2015, 22(2):1–10 [张波, 吴强, 金显仕. 1959–2011年间莱州湾渔业资源群落食物网结构的变化. 中国水产科学, 2015, 22(2):278–287]
Zhu JF, Dai XJ, Wang XF, et al. A review of methodology in marie food-web topology. Progress in Fishery Sciences,2016, 37(2): 153–159 [朱江峰, 戴小杰, 王学昉, 等. 海洋食物网拓扑学方法研究进展. 渔业科学进展, 2016, 37(2):153–159]
猜你喜欢
当代水产(2022年8期)2022-09-20
学苑创造·A版(2021年10期)2021-10-30
小猕猴学习画刊(2021年9期)2021-10-11
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
农民致富之友(2020年27期)2020-09-12
小读者(2020年4期)2020-06-16
当代水产(2019年9期)2019-10-08
江西农业学报(2019年8期)2019-09-04
作文与考试·小学高年级版(2017年9期)2017-05-16
