
氮肥对风箱果幼苗形态和生理特性的影响
2018-11-02 03:20殷东生魏晓慧
植物研究 2018年6期
殷东生 魏晓慧
(1.黑龙江省林业科学研究所,哈尔滨 150081; 2.东北林业大学马克思主义学院,哈尔滨 150040)
施肥是调节土壤养分条件,促进植物生长,提高产量与质量的重要栽培技术措施[1~2]。氮是矿质营养中的核心元素,是限制植物生长发育的关键营养元素[3~4],而且在各种矿质元素含量相同且均未达到充分供应的情况下,植株首先表现出缺氮的症状,增施其他营养元素远没有施氮效果好[5]。近年来,施氮肥影响植物生长和生理变化的研究不断增加[6],一方面,一定范围内施氮肥可明显促进植物的光合作用,有利于植物的生长和生物量的积累,提高苗木质量和竞争力[7];另一方面,过量施氮肥降低了植物的光合作用,抑制了植物的生长和生物量的积累[8],而且施氮肥过量导致土壤酸化,硝酸盐和钙淋失,也会引起土壤污染[9]。因此,研究植物对氮素养分的需求具有重要意义。
风箱果(Physocarpusamurensis)是珍贵的乡土花灌木绿化树种,将该树种引入园林绿化中,对丰富园林景观具有重要意义,开发应用前景非常广阔[10]。目前针对风箱果开展了大量研究[10~12],但是该树种对土壤肥力的适应性研究较少。研究植物对土壤肥力的适应是限制栽培成功的重要生态因素,只有掌握植物对土壤肥力的特殊要求,才能做到成功栽培。本研究以1年生风箱果幼苗为研究对象,研究不同施氮肥水平对幼苗的生长、生物量积累、光合生理参数和碳水化合物积累等的影响,分析幼苗对不同氮肥水平的适应性,为该树种苗木培育和栽植过程中氮肥的合理利用提供理论指导。
1 材料与方法
1.1 试验材料和试验设计
试验材料为风箱果1年生播种苗,种子采自黑龙江省尚志市帽儿山天然种群。2015年4月20日,选取长势一致(苗高平均为14.1 cm,地径平均为2.61 mm)的苗木60株,采用盆栽的方法,盆直径20 cm,高度30 cm(花盆下部具孔),栽培基质为草炭土和沙子(2∶1),基质肥力较低,全氮含量为1.12 g·kg-1(全氮含量与风箱果分布区林下土壤接近),每盆装基质3.0 kg,盆下放置托盘,每盆栽植1株苗木。正常浇水培育,并进行适当管护,缓苗时间为1个月。施肥试验时间于2015年5月20日~9月20日进行。本试验所用肥料为硝酸铵(含氮35%),设置4种施氮肥水平:不施氮肥(CK)、1 g/盆(N1)、3 g/盆(N2)和5 g/盆(N3)。每个处理15株苗木,每15 d施氮肥一次,共施肥7次,将硝酸铵溶于适量水中施入土壤中,若流出盆下托盘中重新倒回土壤中。于试验结束后进行各形态和生理指标的测定。
1.2 采样与测定方法
1.2.1 生长指标和生物量指标的测定
生长量指标测定苗高、地径、分枝数和冠幅,生物量指标测定根生物量、茎生物量、叶生物量和根冠比,具体参照殷东生等[11]的方法进行测定。
1.2.2 净光合速率的测定
净光合速率测定采用美国生产的CI-340手持式光合作用测定系统进行,测定时选取生长较好、完全伸展的叶片为测定对象,测定时间为上午9:00~11:00,测定时用缓冲瓶控制CO2浓度360~380 μmol·mol-1,叶室温度28℃,相对湿度约50%~65%,气体流速为500 μmol·s-1,光合有效辐射为饱和光强1 100 μmol·m-2·s-1[12]。每个处理选3株重复测定,每株选取植株中部的2~3个叶片,每片叶的光合生理参数作3次记录,取平均值作为测定结果。
1.2.3 叶绿素含量的测定
净光合速率测定完以后,选取与测定净光合速率相同部位的叶片,每个施氮肥处理选20片健康的叶片,装入内有冰袋的保鲜袋,迅速带回实验室,叶绿素含量测定参照王文杰等[13]的方法。
1.2.4 叶氮和碳水化合物含量的测定
每个施氮肥处理分别随机选取5株苗木,分别对根、茎和叶等组织取样,带回实验室后,清洗干净后于105℃恒温杀青20 min,在75℃条件下烘干至恒重,用不锈钢粉碎机粉碎过80目筛后备样。本研究测定的碳水化合物为非结构性碳水化合物(NSC),即可溶性糖(葡萄糖、蔗糖、果糖等)和淀粉的总和。可溶性糖和淀粉的含量采用于丽敏等[14]的方法测定。叶氮含量采用元素分析仪(Vario MAX CN Elemental Analyzer,Elementar,Germany)进行测定。
1.3 数据处理
所有数据均通过Excel进行整理,采用SPSS 18.0统计软件对不同施氮肥处理的生长指标、生物量指标、光合生理参数、叶氮含量、非结构性碳(NSC)等进行单因素方差分析和多重比较。
2 结果与分析
2.1 施氮肥对风箱果幼苗生长的影响
从表1可以看出,施氮肥对风箱果幼苗的苗高增量影响不显著(P>0.05),对地径,分枝数和冠幅增量影响显著(P<0.05)。N1处理的地径增量与CK差异不显著(P>0.05),而N2和N3处理的地径增量显著高于CK(P<0.05);施氮肥处理的分枝数增量显著高于CK(P<0.05),但施氮肥处理间(N1、N2、N3)的分枝数增量差异不显著(P>0.05);N2和N3处理的冠幅增量显著高于CK(P<0.05),而N3处理的冠幅增量与CK差异不显著(P>0.05)。
表1施氮肥对风箱果幼苗生长的影响
Table1EffectofnitrogenfertilizationongrowthofP.amurensisseedlings
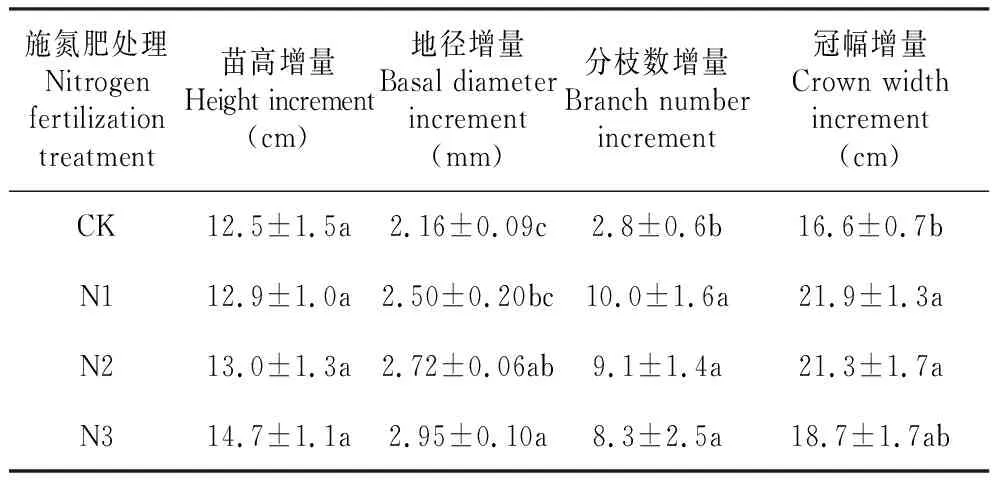
施氮肥处理Nitrogen fertilization treatment苗高增量Height increment(cm)地径增量Basal diameter increment(mm)分枝数增量Branch number increment冠幅增量Crown width increment(cm)CK12.5±1.5a2.16±0.09c2.8±0.6b16.6±0.7bN112.9±1.0a2.50±0.20bc10.0±1.6a21.9±1.3aN213.0±1.3a2.72±0.06ab9.1±1.4a21.3±1.7aN314.7±1.1a2.95±0.10a8.3±2.5a18.7±1.7ab
注:同列不同字母表示差异显著(P<0.05),下同。
Note:Different letters in the same column represented significant difference at 0.05 level,the same as below.
2.2 施氮肥对风箱果幼苗生物量积累和分配的影响
从表2可以看出,施氮肥对根生物量和叶生物量比影响不显著(P>0.05),显著增大了茎生物量、叶生物量、总生物量和茎生物量比,显著减小了根冠比、根生物量比(P<0.05),各生物量指标在施氮肥处理间(N1、N2、N3)差异不显著(P>0.05),但叶生物量比随施氮量的增加呈逐渐增大的趋势,其它生物量指标呈逐渐减小的趋势。
2.3 施氮肥对风箱果幼苗净光合速率、叶氮和叶绿素含量的影响
从表3可以看出,施氮肥显著提高了风箱果幼苗的净光合速率和叶氮含量(P<0.05),施氮肥处理间(N1、N2、N3)差异不显著(P>0.05);N1和N2处理的叶绿素a、叶绿素b和总叶绿素含量与CK差异不显著(P>0.05),N3处理的叶绿素a、叶绿素b和总叶绿素含量显著低于CK(P<0.05)。
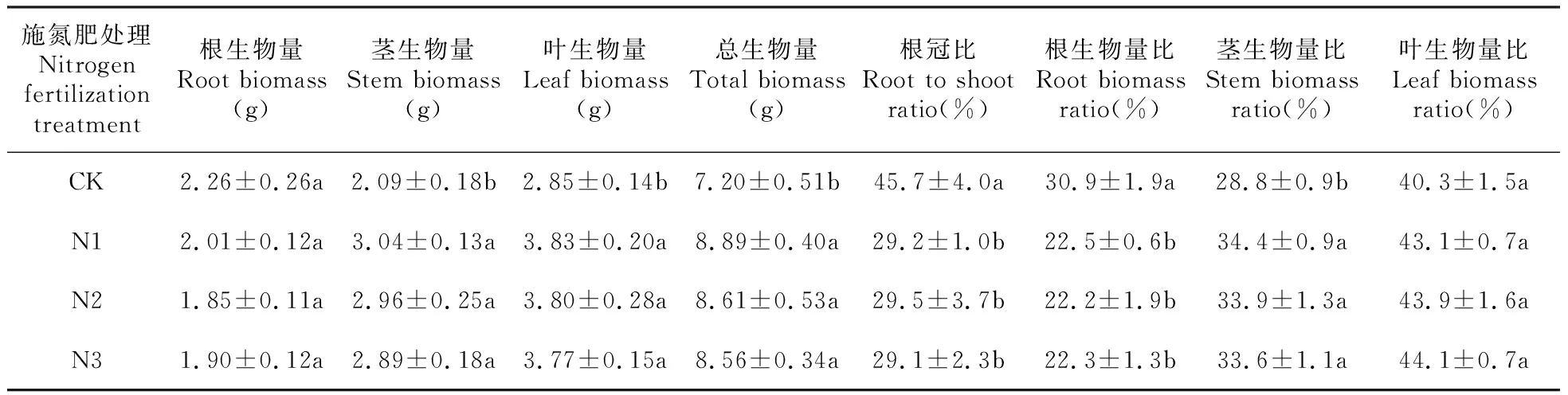
表2 施氮肥对风箱果幼苗生物量分配的影响
表3施氮肥对风箱果幼苗净光合速率、叶氮含量和叶绿素含量的影响
Table3Effectofnitrogenfertilizationonnetphotosyntheticrate,leafnitrogencontentandchlorophyllcontentofP.amurensisseedlings
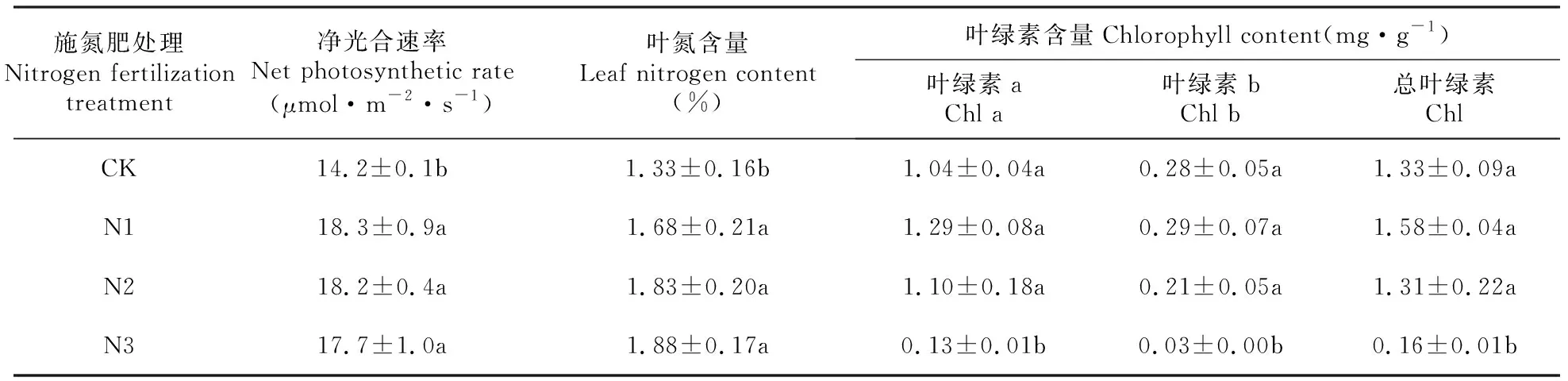
施氮肥处理Nitrogen fertilization treatment净光合速率Net photosynthetic rate(μmol·m-2·s-1)叶氮含量Leaf nitrogen content(%)叶绿素含量Chlorophyll content(mg·g-1)叶绿素aChl a叶绿素bChl b总叶绿素ChlCK14.2±0.1b1.33±0.16b1.04±0.04a0.28±0.05a1.33±0.09aN118.3±0.9a1.68±0.21a1.29±0.08a0.29±0.07a1.58±0.04aN218.2±0.4a1.83±0.20a1.10±0.18a0.21±0.05a1.31±0.22aN317.7±1.0a1.88±0.17a0.13±0.01b0.03±0.00b0.16±0.01b
2.4 施氮肥对风箱果幼苗NSC积累和分配的影响
从表4可以看出,施氮肥对风箱果幼苗根和茎中的可溶性糖和NSC含量影响显著(P<0.05),对淀粉含量影响不显著(P>0.05),对叶中的可溶性糖、淀粉和NSC含量影响不显著(P>0.05)。根中N1处理的可溶性糖和NSC含量与CK差异不显著(P>0.05),N2和N3处理显著低于CK(P<0.05);茎中N1处理的可溶性糖和NSC含量显著高于CK(P<0.05),N2和N3处理与CK差异不显著(P>0.05)。
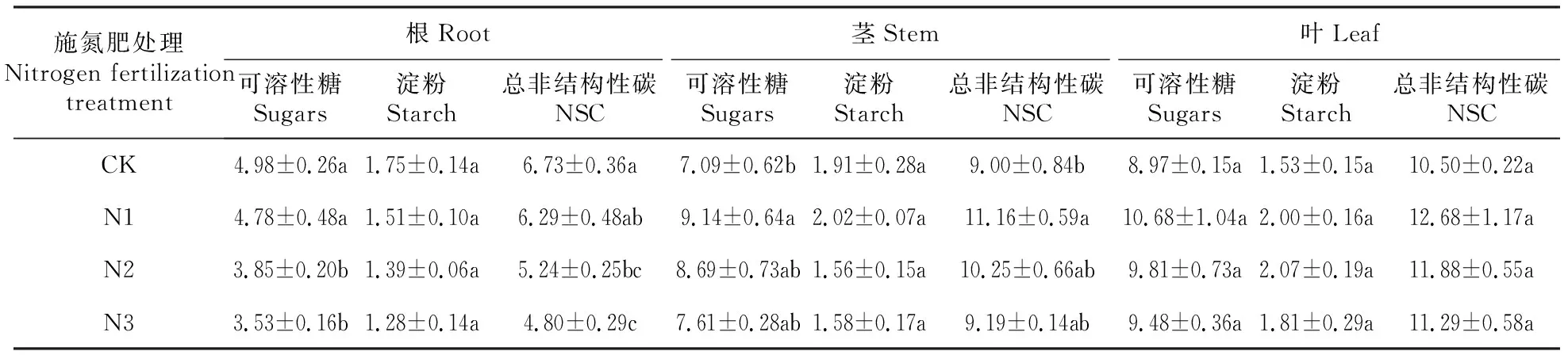
表4 施氮肥对风箱果幼苗根、茎、叶中NSC含量的影响
3 讨论
形态特征是植物适应环境变化最直接的外在表现,植物对其的调节能力很强,其中苗高、地径、冠幅等调整最为明显[15~16]。大量研究证实植物的苗高、地径、分枝数等生长指标都随养分供应水平的提高而显著增大[17~18]。本研究结果表明,施用氮肥显著促进了风箱果幼苗的地径生长,增加了分枝数量、增大了冠幅(表1),证实了上述结论。生物量在根系和茎、叶之间的分配格局及其变化是植物对养分缺乏在形态变化方面的适应和反应的重要表征[19]。本研究中,施用氮肥显著增大了茎和叶生物量的分配,显著减少了根冠比和根生物量比(表2)。分析认为,氮供给不足时,导致较多的生物量向根系分配,促进根系的生长,提高根系吸收养分的能力,导致根生物量、根生物量比和根冠比增大[20];而氮供给充足时,根系更容易获得土壤中的养分,因此降低了根生物量分配,将更多的生物量分配给地上部分用于竞争其他资源(例如光照),导致茎生物量、叶生物量、茎生物量比和叶生物量比的增大[7,21],其他研究也证实了这一结论[17~18]。
氮素对植物生长发育的影响与叶片光合作用存在直接关系,净光合速率、叶氮和光合色素含量比形态指标更能反映幼苗在不同氮供给水平下的生理状况[6,20],通常,这些指标在一定的供氮范围内随供氮量的增加而显著增加[18,22]。本研究发现,施用氮肥显著提高了风箱果幼苗的净光合速率和叶氮含量,明显增加了叶绿素含量(表3)。分析认为由于施用氮肥增加了叶氮含量,改变了与光合密切相关的酶的含量[23~25],增加了光合色素含量,增强了叶片吸光能力和叶肉细胞的光合活性[17,26],最终提高了净光合速率,从而促进植株的生长和生物量的积累。通常,随着施氮量的增加能够提高叶绿素含量[17],但是本研究中N3处理的叶绿素含量显著低于CK(表3),可能是施氮肥过量引起植物体内的矿物营养失衡导致的[18],特别是钾元素失衡会引起叶绿素降解[27],具体原因有待进一步研究。
NSC是树木生长代谢过程中重要的能量供应物质[14,28~29]。本研究发现施用氮肥显著减少了风箱果幼苗根中的可溶性糖和NSC含量,对根中的淀粉影响不显著(表4),说明根中NSC的减少主要是由于可溶性糖含量的减少引起的。施用氮肥能够明显增加茎和叶中的可溶性糖、淀粉和NSC的积累(表4)。大量研究已经证实,当土壤氮素充足时,碳水化合物向根部的分配减少,向茎叶分配比例增加[30]。分析认为土壤氮素充足时,根系输送给茎和叶的细胞分裂素增加,茎和叶的细胞分裂加快,导致碳水化合物由韧皮部向茎和叶输送的量增加,从而减少了韧皮部附近的碳水化合物含量,引起叶片膨压的降低;而根系细胞分裂继续进行,细胞膨压没有发生变化,从而使源(叶片)和汇(根部)之间产生膨压梯度,促使碳水化合物相对较多地向茎、叶分配[31]。本研究发现,N1处理显著提高了茎中可溶性糖和NSC的积累,而N2和N3处理茎中可溶性糖和NSC与CK差异不显著(表4),说明过量施氮肥并不利于碳水化合物的积累,与其他植物的研究结果一致[32~33]。
本研究中的大部分生长指标、生物量指标、光合生理参数和NSC等在施肥处理间(N1、N2、N3)的差异均不显著(表1~4),说明风箱果幼苗对土壤养分的变化并不敏感,进一步说明风箱果较耐贫瘠,对土壤肥力条件要求不高,生产实践中对栽植的土壤条件适应性广。
猜你喜欢
戏剧之家(2022年25期)2022-10-31
中国交通信息化(2022年7期)2022-10-27
戏剧之家(2022年17期)2022-06-16
今日农业(2020年19期)2020-12-14
农业与技术(2020年12期)2020-07-04
安徽农学通报(2020年7期)2020-05-26
中国野生植物资源(2019年2期)2019-06-11
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
北京观察(2014年10期)2014-12-13
