
滇楸种质生长性状遗传变异及表型性状遗传多样性分析
2018-11-02 03:20麻文俊杨桂娟王秋霞王军辉
植物研究 2018年6期
肖 遥 麻文俊 易 飞 杨桂娟 王秋霞 王 平 王军辉*
(1.中国林业科学研究院林业研究所,林木遗传育种国家重点实验室,国家林业局林木培育重点实验室,国家林木种质资源平台,北京 100091; 2.河南省南阳市林业科学研究院,南阳 473000)
滇楸(Catalpafargesiif.duclouxii)属紫葳科(Bignoniaceae)梓树属(Catalpa)落叶乔木。其主要分布在四川、云南及贵州等地区。滇楸树干通直,材质优良,其木材是制作高档家具及木制品的珍贵用材。此外,滇楸树形优美,枝叶繁茂,花色艳丽,常作为园林观赏树种和绿化树种[1~3]。然而由于过度砍伐利用的原因,其天然资源已相当匮乏[4],人工选育及创制新种质是改善和利用其种质资源现状的有效途径之一[5~6]。
林木的遗传变异是改良的基础,充分了解其变异规律及遗传多样性大小能为合理制备育种策略提供依据,有效地减少育种工作的盲目性,大大提高改良的效果[7~8]。在分类学上滇楸是灰楸(C.fargesii)的变种,与灰楸同属楸树组。不同于楸树(C.bungei)和灰楸,滇楸自然结实性较好,长期的有性繁殖过程中累积了较多的遗传变异[9],这为滇楸的遗传改良奠定了良好基础。然而滇楸遗传育种工作长期处于停滞状态,截止目前相关研究主要局限于苗期生长的遗传变异[4,10~11]。遗传多样研究仅见滇楸花部性状的多样性,研究结果表明其花部性状在群体间和群体内均存在丰富的变异,各性状平均变异系数为25.94%,平均表型分化系数为27.95%[2]。就研究现状看,滇楸遗传变异较大,多样性丰富,具有较大的利用潜力。但相关研究程度较低且新种质创制缺乏科学性。鉴于此,本研究选取了20个滇楸无性系,分析了其生长性状遗传变异规律、叶部性状和皮孔性状遗传多样性,旨在加速滇楸优良种质选育进程,科学选择杂交育种亲本,同时也为其种质资源的保护和利用提供科学依据。
1 材料与方法
1.1 试验材料与试验地概况
滇楸种质材料采集自贵州瓮安、织金、贵定、紫云、毕节、关岭、晴隆、盘县、普安、兴仁、贞丰、安龙和兴义13个地区,涵盖了黔中、黔北亚热带湿润温和气候区、黔西暖温带干湿交替温凉气候区等多个气候区。共收集的151个滇楸优良单株种子,对151个家系进行评比试验,筛选出超级苗。再将超级苗无性系化,进行优良无性系筛选。对选出的优良无性系在进行三级筛选。一级筛选采用间比法无重复试验设计;二级为随机完全区组设计[12]。2008年进行无性系的扩繁嫁接,采用随机区组设计,7株小区,共4个区组。于2009年春季采用2年生苗在河南省南阳市潦河镇对二级筛选后的无性系进行移栽造林,开展三级大田试验,试验采用完全随机区组设计,2株小区,5个区组。试验材料详见表1。
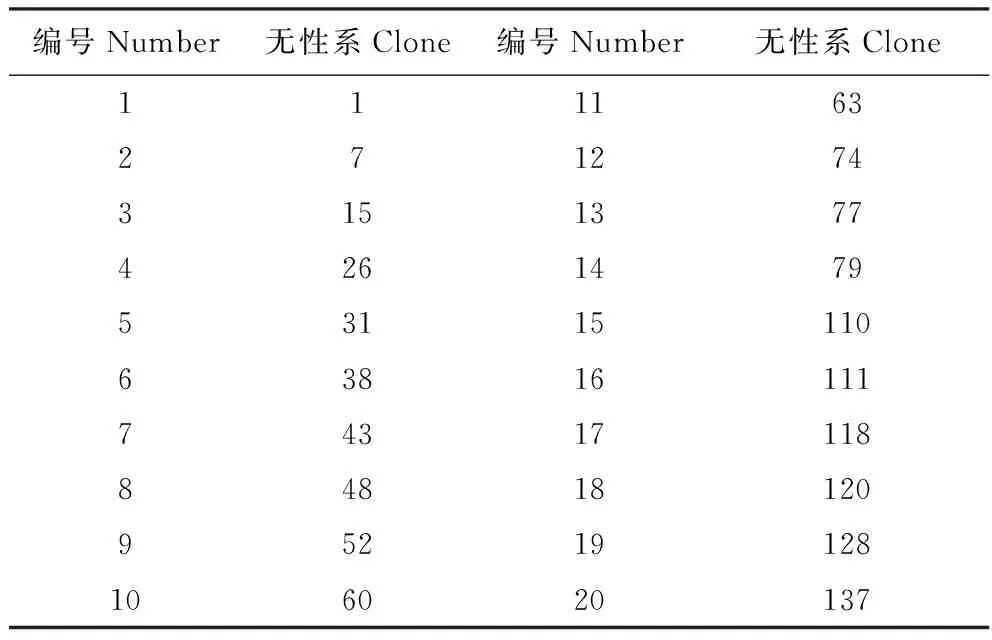
表1 试验材料
试验地位于河南省南阳市潦河镇潦东村(32°.93′N,112°.41′E),属季风大陆湿润半湿润气候,四季分明。海拔145 m,土壤类型为黄棕壤土,自然肥力较高。年平均气温14.4~15.7℃,七月平均气温26.9~28.0℃,一月平均气温0.5~2.4℃。年降雨量703.6~1173.4 mm。年日照时数1897.9~2120.9 h,年无霜期220~245 d。
1.2 方法
2008~2013年,每年年末对滇楸无性系试验林进行每木调查,测定其树高和胸径。2013年末选取3个保存较好的区组,每个区组选择1株生长良好的植株,分别测定其叶长、叶宽、叶柄长、叶痕长、叶痕宽和叶痕面积,每个性状测定3次重复,叶片部位均选择与向南枝条的中部生长良好的功能叶。皮孔性状测定:选择树干上、中、下3个面积为2 cm2×3 cm2大小的区域,计算每个区域皮孔数量,测定每个皮孔的长度、宽度并计算其面积。叶痕面积和皮孔面积计算方法参考文献[13]。
1.3 数据处理
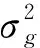
滇楸生长性状线性模型为:
Yijk=μ+Bi+Cj+BCij+eijk
(1)
式中:Yijk第i区组内第j无性系的性状观察值;μ为群体平均效应;Bi为第i区组效应;Cj为第j无性系效应;BCij为第i区组和第j无性系的互作效应;eijk为机误。
叶部和皮孔性状线性模型为:
Yijk=μ+Bi+Cj+eijk
(2)
式中:Yijk第i区组内第j无性系的性状观察值;μ为群体平均效应;Bi为第i区组效应;Cj为第j无性系效应;eijk为机误。
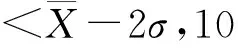
表型变异系数公式[16]:
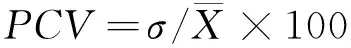
(3)
式中:σ为标准差;X为性状均值。
遗传变异系数公式[16]:
(4)
表型相关公式[17]:
(5)

遗传相关公式[17]:
(6)
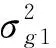
2 结果与分析
2.1 滇楸种质生长性状遗传变异
2.1.1 生长性状的方差分析
生长性状的方差分析表明(表2),滇楸无性系间仅2和6 a树高不存在显著差异,1、3、4和5 a树高在无性系均具有显著差异,其中1和5 a树高的无性系差异达5%的极显著水平。滇楸各年胸径在无性系间均具有极显著差异。两种性状的方差分析结果差异可能原因在于树高和胸径受环境影响的程度不同,树高更易受到环境影响。而无性系生长性状差异的结果意味着滇楸无性系遗传变异明显,具有较好的改良基础。另一方面,区组效应对滇楸无性系各年的高生长及胸径生长影响显著,仅1a胸径没有显著的区组效应。这意味着在选育滇楸优良无性系不能忽略环境效应的影响。区组与无性系的交互效应到了5年生和6年生时不再存在,表明滇楸无性系遗传稳定性加强了,选择出的优良无性系在不同的环境中生长都更具优势。
2.1.2 生长性状遗传参数的年份变化
重复力的大小能反映遗传因素对林木生长影响的程度,由图1可知滇楸无性系树高和胸径重复力年度变化差异较大。树高重复力随树龄变化波动较大,1 a树高重复力较高,而2 a树高重复力接近0。这可能与当年移栽造林,苗木处于扎根阶段影响了无性系的高生长,进而影响树高重复力。6 a树高重复力下降较大,可能与当年低温条件有一定关系。不同的是,胸径重复力从3~6 a一直保持相对稳定的状态,平均达0.69。这表明环境的改变对滇楸高生长影响显著,而相反胸径受遗传因素控制的程度较强且稳定性较高。
滇楸无性系各树龄树高和胸径的遗传变异系数较小,表型变异系数相对较大(图2)。对比各树龄树高和胸径的遗传变异系数可见,树高的遗传变异系数年度变化幅度较大,而胸径的遗传变异系数相对稳定,这与树高和胸径重复力年度变化规律一致。2 a树高遗传变异系数接近于0也说明了造林初期苗木处于扎根期影响其树高生长和遗传参数估计。此外,胸径的遗传变异系数大于树高的遗传变异系数,在一定程度上表明了胸径受遗传因素控制的程度大于树高。
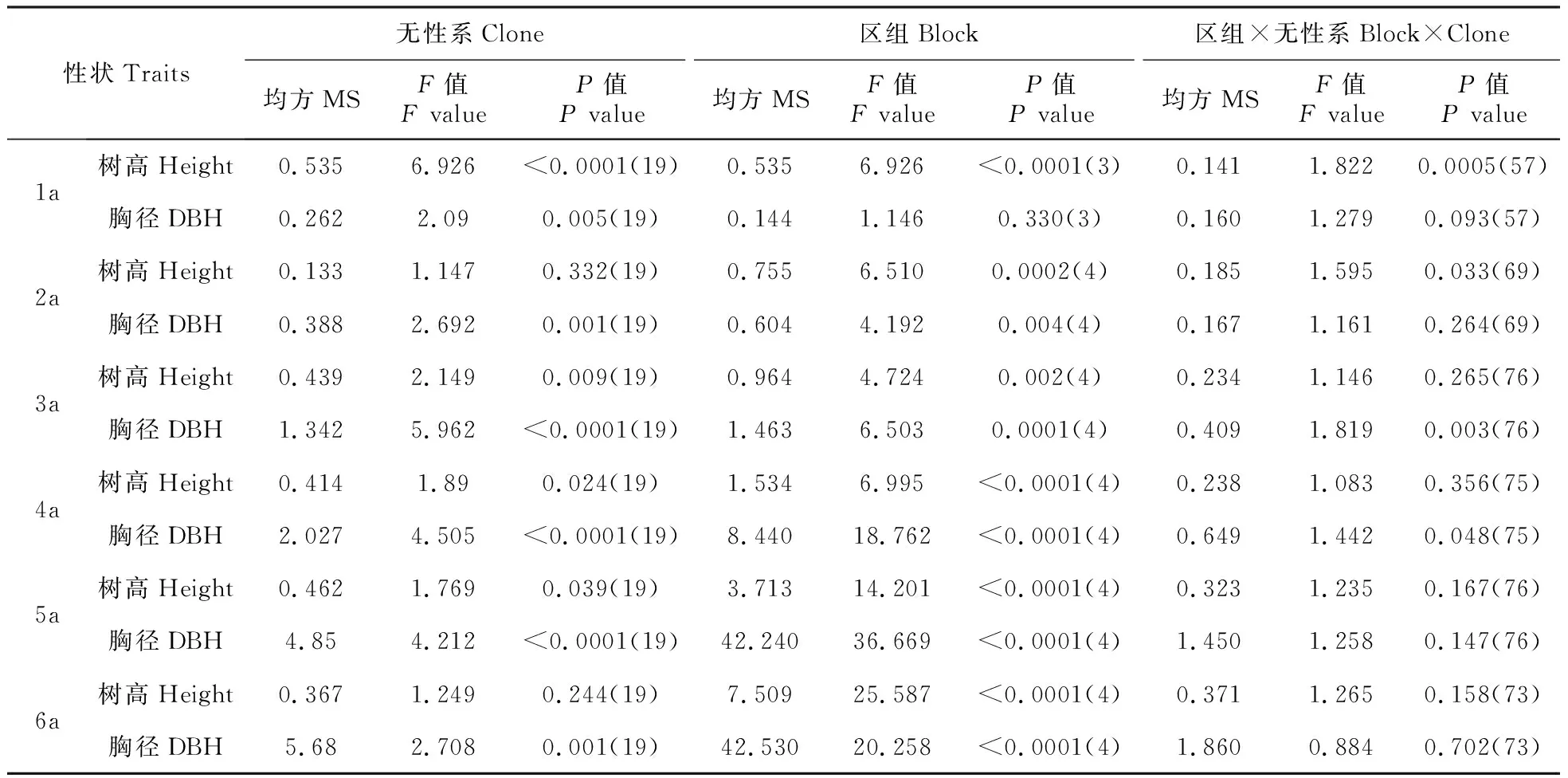
表2 滇楸无性系生长性状方差分析
注:括号里的数字为自由度。
Note:The figures in brackets were degree of freedom.
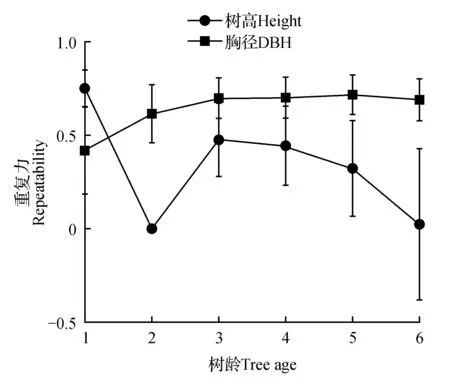
图1 不同树龄滇楸无性系树高和胸径重复力差异Fig.1 Repeatability of height and DBH of C.fargesii f. duclouxii in different ages
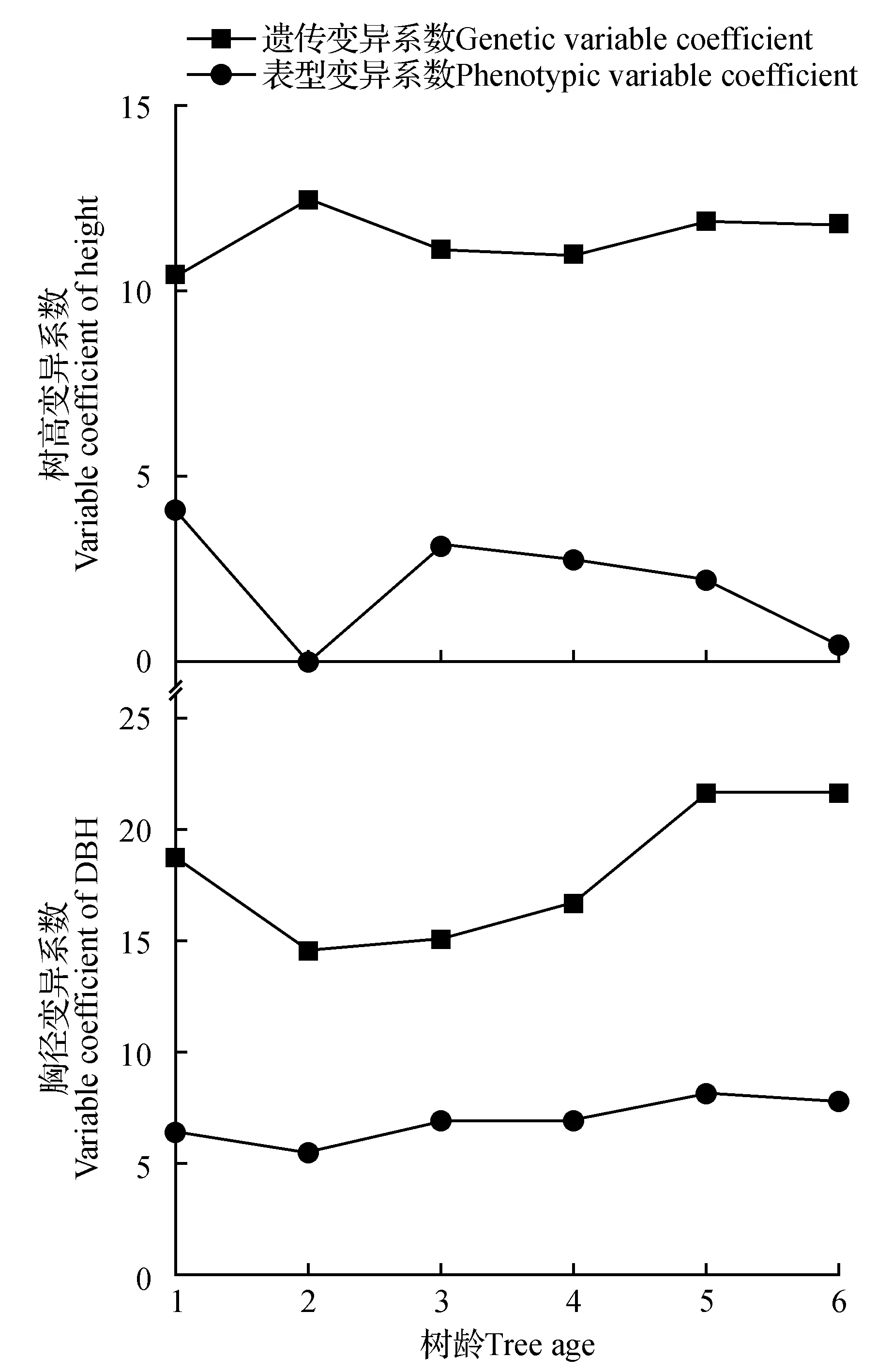
图2 不同树龄滇楸无性系树高和胸径变异系数差异Fig.2 variable coefficient of height and DBH of C.fargesii f. duclouxii in different ages
2.2 滇楸种质叶部性状遗传变异
2.2.1 表型性状方差分析
表3滇楸种质表型性状方差分析结果表明,无性系间叶部性状存在一定差异,叶宽和叶长宽比在无性系间分别达到了5%水平和1%水平的显著差异。皮孔相关性状变异较大,皮孔长、皮孔宽、皮孔面积和皮孔密度在无性系间均存在现在差异,其中皮孔密度的差异达到5%的极显著差异(P=0.004)。说明滇楸种质表型变异显著,从表型性状研究和评价滇楸种质遗传变异有可靠依据。
2.2.2滇楸种质叶部性状和皮孔性状遗传多样性分析
滇楸种质遗传多样性分析结果发现(表4),其叶柄长度表型变异达20.95%,是叶部性状中变异程度最大的性状,极大值为极小值的3.6倍。其余叶部性状表型变异系数相对较低,均未超过20%。不同的是,滇楸种质皮孔性状变异幅度较大,表型变异系数除皮孔宽为17.37%,其余性状均超过25%,且皮孔面积变异系数高达49.69%。叶长、叶长宽比虽变异系数较小,但遗传多样性指数多样性指数较高分别为2.016和2.012。说明滇楸种质叶形变异处于遗传分化的过度时期。滇楸种质叶长宽比具有最大的重复力0.849,叶宽重复力较为适中为0.539。表明叶形的宽度和长宽比主要受遗传因素影响,长度主要受环境因素影响。皮孔各性状的重复力在0.5左右,表明滇楸皮孔性状遗传控制程度较为适中。
2.3 生长性状和叶部性状的相关分析
生长性状和表型性状相关分析结果显示(表5),参试的滇楸种质各表型性状和生长性状均未达到显著相关。可能的原因是,滇楸种质叶部性状、皮孔性状与生长性状的相关性并不是线性相关。而Pearson相关主要分析的是性状与性状间的
线性相关大小。从相关系数大小分析,叶痕长和宽与树高呈中偏弱的负相关,皮孔密度与胸径也呈中偏弱的负相关。而皮孔长、宽及面积与树高呈较弱的正相关。
表3滇楸无性系表型性状方差分析
Table3VarianceanalysisofphenotypictraitsofC.fargesiif.duclouxii
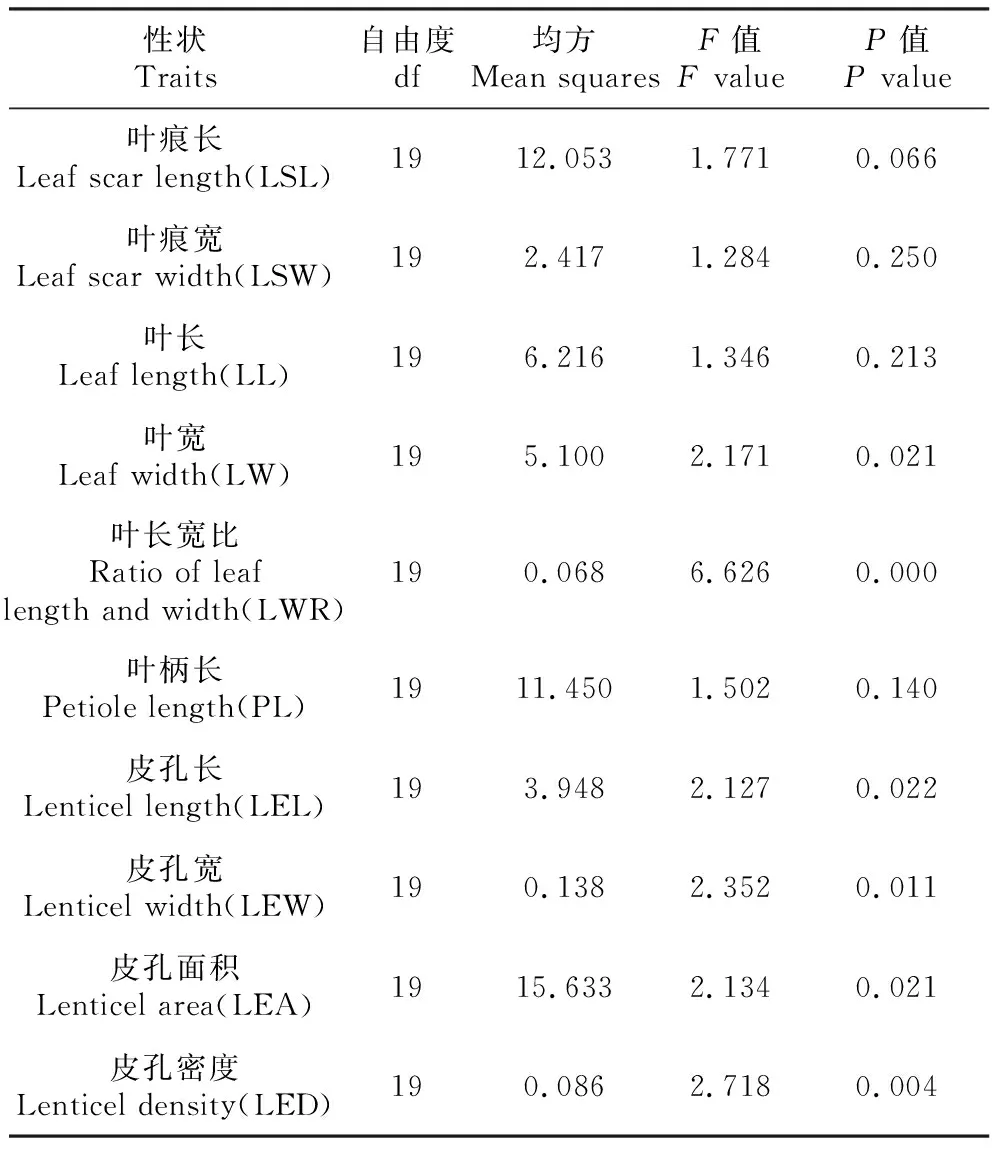
性状Traits自由度df均方Mean squaresF值F valueP值P value叶痕长Leaf scar length(LSL)1912.0531.7710.066叶痕宽Leaf scar width(LSW)192.4171.2840.250叶长Leaf length(LL)196.2161.3460.213叶宽Leaf width(LW)195.1002.1710.021叶长宽比Ratio of leaf length and width(LWR)190.0686.6260.000叶柄长Petiole length(PL)1911.4501.5020.140皮孔长Lenticel length(LEL)193.9482.1270.022皮孔宽Lenticel width(LEW)190.1382.3520.011皮孔面积Lenticel area(LEA)1915.6332.1340.021皮孔密度Lenticel density(LED)190.0862.7180.004
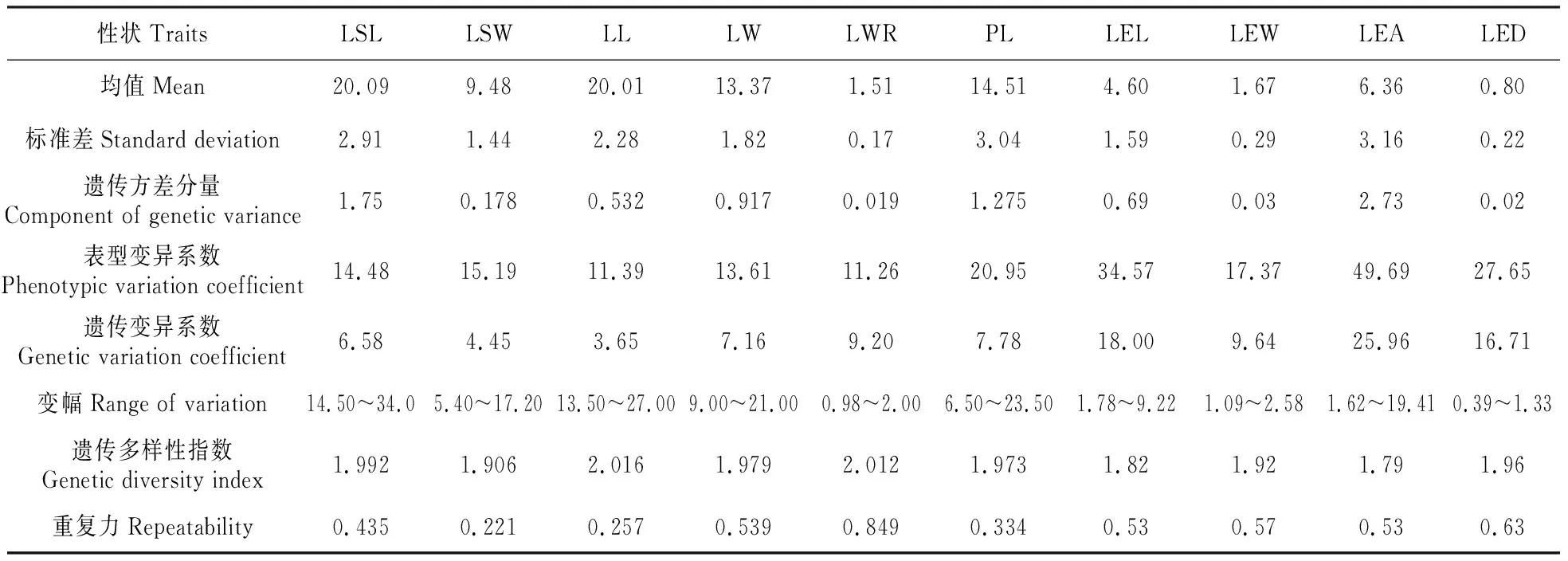
表4 滇楸叶部性状遗传变异与遗传多样性
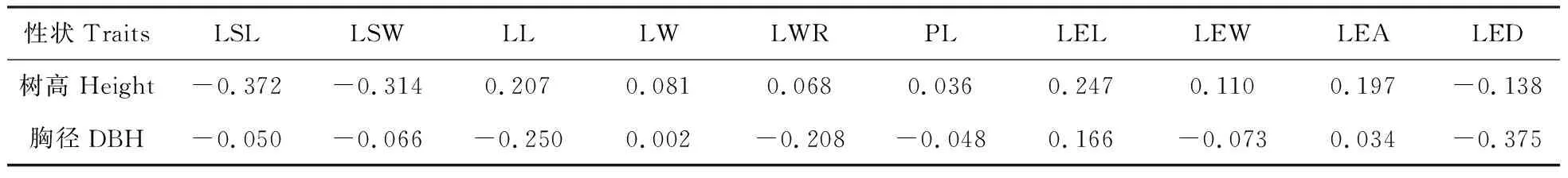
表5 滇楸树高、胸径与叶部性状的Pearson相关性
2.4 滇楸种质生长性状和表型性状主成分分析和聚类分析
2.4.1 主成分分析
为了更加清晰的明确各性状在滇楸表型多样性构成中的作用大小,采用主成分分析的方法使多个表型性状降维成几个主要的成分因子综合的反应原有的多种性状(表6)。主成分分析的前4个成分累积贡献率达81.02%,表明这4个主成分能反应原有的12个性状的绝大信息。根据载荷值大小可知,第一主成分与皮孔相关的载荷值绝对值较大,将改主成分定义为皮孔形态因子,其贡献率最高,占32.72%。第二主成分载荷较高的性状有,叶痕长、叶痕宽、叶长、叶宽,可称改主成分为叶形因子,其贡献率达21.68%,第三类成分与树高和胸径较为相关,称为生长因子,占比为16.07%。第四类主成分载荷较高的性状分散于生长、叶部和皮孔性状,是对之前的主成分信息的补充,因此定义为综合因子。
表6滇楸种质性状主成分分析
Table6PrincipalcomponentanalysisoftraitsofC.fargesiif.duclouxii

分量来源Component主成分名称Name of component皮孔形态因子Lenticel type factor叶形因子Leaf type factor生长因子Growth factor综合因子Comprehensive factor树高Height-0.127-0.2820.377-0.326胸径DBH-0.067-0.0190.522-0.495皮孔长Lenticel length-0.444-0.0030.1400.161皮孔宽Lenticel width-0.4180.1300.0490.305皮孔面积Lenticel area-0.4670.0590.0890.259皮孔密度Lenticel density0.350-0.134-0.318-0.039叶痕长Leaf scar length0.0310.528-0.084-0.002叶痕宽Leaf scar width0.0800.5180.0080.084叶长Leaf length-0.080-0.487-0.2400.200叶宽Leaf width0.229-0.2840.2890.524叶长宽比Length-width ratio-0.285-0.083-0.473-0.345叶柄长Petiole length0.3470.1000.2880.143特征值Eigen values1.9811.6131.3891.125贡献率Proportion of variance(%)32.7221.6816.0710.55累积贡献率Cumulative proportion(%)32.7254.4070.4781.02
2.4.2 聚类分析
以欧氏距离作为衡量单位,聚类分析将滇楸种质20个无性系分为了4大类(图3)。第Ⅰ类群包含4个无性系,其皮孔长、皮孔宽和皮孔面积最大,皮孔密度最小,叶长宽比较高。第Ⅱ类包含137、74和120共3个无性系,该类群树高和胸径显著小于其他3类,但皮孔密度最大,叶长和叶宽也最大,叶形偏长条型。第Ⅲ类树高和胸径均最大,属于生长最快的类群。第Ⅳ类群叶柄最长,皮孔大小最小,叶长宽比最小,叶形属于偏宽型的。从聚类分析可见,叶部性状及皮孔性状较大的类群生长不一定优越,相对的叶部性状或皮孔性状偏小的生长不一定较差。这表明叶部性状与皮孔性状的相关并不是呈简单的线性关系。

图3 滇楸无性系聚类分析Fig.3 Clustering analysis of C.fargesii f. duclouxii clones

性状Traits类群GroupsⅠⅡⅢⅣ树高Height(m)6.256.076.356.10胸径DBH(cm)8.367.548.878.28皮孔长Lenticel length(mm)6.253.664.823.78皮孔宽Lenticel width(mm)2.001.611.631.53皮孔面积Lenticel area(mm2)10.274.806.344.69皮孔密度Lenticel density(个·cm-2)0.640.980.770.86叶痕长Leaf scar length(mm)20.2320.0619.4520.75叶痕宽Leaf scar width(mm)9.509.249.299.82叶长Leaf length(cm)20.0021.0920.6418.74叶宽Leaf width(cm)12.5812.6013.9613.59叶长宽比Ratio of leaf length and width1.601.681.491.39叶柄长Petiole length(cm)12.3213.2014.9816.07
3 讨论
在长期的生长演变过程中,林木会积累大量的遗传变异来逐渐适应变化的环境。掌握林木的遗传变异规律是利用林木种质资源的有力依据。变异导致物种多样性产生的前提,所以遗传多样性与遗传变异密切相关。研究遗传变异和遗传多样性对选育优良种质[18~19],杂交亲本的合理选择及开展高世代育种[20~22],明确林木生长的环境影响[23]等均有重要的意义。本研究结果发现滇楸无性系生长性状的遗传变异较大,20个无性系间树高和胸径差异显著,具有较好的遗传改良基础,为选择优良种质提供了可行性。通过其生长性状遗传参数的年份变化发现,滇楸无性系树高的重复力和遗传变异系数树龄动态曲线波动较大,而胸径的重复力和遗传变异系数变化则相对稳定。这与日本落叶松(Larixkaempfeni)家系生长性状遗传参数的树龄变化结果类似[24],这意味着滇楸种质胸径的遗传稳定性较强。此外,胸径的重复力平均高于树高的,说明胸径受遗传控制程度更高[8,25~26]。因此,评价滇楸生长情况,选育速生高产品种时以胸径作为衡量指标更适佳。
滇楸种质叶部性状有一定程度的变异,无性系间差异主要存在于叶宽和叶长宽比上。群体的叶长和叶长宽比的遗传多样性较大,均大于2。叶长宽比作为滇楸主要的叶形变异其表型变异系数较低为11.26%,重复力高达0.849。说明该性状比较稳定,且主要受遗传因素控制。这与楸树[7]、核桃[27]和白桦[28]叶部性状遗传变异较大,且受遗传控制较强的结果类似。滇楸皮孔长、皮孔面积和皮孔密度遗传变异系数较大,超过15%,重复力接近0.5受中度的遗传控制。贠慧玲等[13]研究发现楸树皮孔性状遗传变异系数超过20%,重复力高达0.7。从皮孔性状变异程度上看与本研究类似,但重复力大小差异较大。这可能是材料生长的环境极其物种生物学特性差异导致的。综合结果看,滇楸种质表型性状具有丰富的遗传变异,遗传多样性较高,充分挖掘其表型变异能为优良种质选育及其杂交亲本选育等工作的开展提供有利依据。
相关分析结果表型性状与生长性状相关程度较弱,但仍表现出一些规律。皮孔的密度与生长性状呈负相关,皮孔大小与生长呈正相关。说明皮孔越大树木与外界环境进行的物质交换效率越大,有利于其生长。类似的结果也出现在楸树[13]和小黑杨[29]的研究中。聚类分析的结果将滇楸种质分为了4类,类群Ⅰ为大皮孔、低皮孔密度型;类群Ⅱ为生长较慢,长叶型;类群Ⅲ速生,宽叶型;类群Ⅳ为长叶柄型。根据不同育种目的其利用的途径也不同,如若要培育速生种质可选择Ⅰ类和Ⅲ类作杂交亲本。若要培育大叶观赏滇楸种质可选择Ⅱ类和Ⅲ类进行杂交育种。
猜你喜欢
农业工程学报(2022年6期)2022-06-27
中国烟草科学(2022年2期)2022-05-27
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
海南医学(2020年1期)2020-01-18
电子制作(2019年24期)2019-02-23
湖北农业科学(2018年18期)2018-12-11
农业研究与应用(2014年1期)2014-08-15
食品工业科技(2014年5期)2014-03-11
