
miR-300通过靶向BRD7对肝癌细胞增殖与侵袭的影响
2018-11-14 03:45邓爱民李建华
中国老年学杂志 2018年21期
邓爱民 李建华
(郑州澍青医学高等专科学校,河南 郑州 450064)
目前,针对肝癌最有效的治疗方法是手术治疗〔1〕,然而85%的患者在诊断时已发生远处转移,单纯的手术切除甚至肝脏移植都不能获得很好的治疗效果〔2〕。MicroRNA(miRNAs)为单链非编码小分子RNA,通过与靶基因互补结合抑制基因的蛋白翻译或者降解mRNA使基因的表达水平下调。研究发现,miRNAs通过调节细胞增殖、凋亡、侵袭和转移等过程参与肿瘤的发生发展〔3〕。同样,许多miRNAs在肝癌的发生过程中发挥重要作用。比如miR-135a通过作用于FOXO1促进肝癌细胞的迁移和侵袭过程〔4〕。而miR-300在骨肉瘤和胃癌组织中均有较高的表达水平,在骨肉瘤中miR-300通过靶向下调溴含蛋白质(BRD)7参与肿瘤的增殖和侵袭过程,但是关于miR-300在肝癌中发挥作用的研究较少。本文运用肝癌细胞系HepG2和Hep3b,探讨miR-300对肝癌细胞增殖和侵袭的影响,并通过过表达实验判断miR-300的靶基因BRD7是否参与该过程。
1 材料与方法
1.1细胞与质粒 人肝癌细胞系HepG2、Hep3b和人胚肾上皮细胞293T细胞由中国科学院上海细胞库提供,其中HepG2和Hep3b生长于RPMI1640培养基中〔含10%灭活胎牛血清(FBS)、青霉素(100 U/ml)、链霉素(100 μg/ml)、2 mmol/L L-谷氨酰胺〕。293T细胞生长于10% FBS DMEM培养基中。内参质粒为海肾荧光素酶的质粒pRL-TK,购自美国 Promega 公司。pGL3-control报告质粒由本实验室提供,同时本实验室将质粒(序列为BRD7 3′UTR)插入pGL3-control虫荧光素酶序列下游,测序后命名为pGL3-BRD7 3′UTR。miRNAs定量引物(广州锐博生物科技公司)。上海吉玛制药技术有限公司合成miR-300模拟物(mimic)和阴性对照(NC)。
1.2荧光素酶实验 将293T细胞(规格为2×104个/孔)接种于48孔板,同时加入0.2 ml DMEM培养基孵育。次日,根据LipofectamineTM 2000(Invitrogen,CA,USA)说明书在293T细胞内将miR-300 mimic和NC分别与pGL3-control或pGL3-BRD7 3′UTR及pRL-TK质粒共同转染。转染48 h后,收集细胞,应用GLOMAX多功能酶标仪分析荧光素酶。
1.3实时荧光定量PCR(qRT-PCR)检测miRNAs的表达 于实验前1 d在6孔板中铺入5×104HepG2和Hep3b,次日细胞贴壁,细胞汇合度在60%~70%,依据LipofectamineTM 2000说明书操作,将miR-300 mimic和NC分别转染入HepG2和Hep3b细胞,转染48 h后,Trizol试剂提取总RNA,按照SYBR®Premix Ex TaqTM(Tli RNaseH Plus)说明书建立反转录体系,qRT-PCR实验的操作由ABI7300定量PCR仪完成。所用引物序列见表1。反应温度: 95℃ 30 s 预变性,95℃ 5 s,60℃ 31 s 循环40次。Ct值按照Applied Biosystem公司 ABI PRISM 7700 Sequence Detection System User Bulletin #2推荐方法获得,并进行计算,比较目的基因在细胞中的表达水平。采用Real-Time PCR仪器(ABI公司,型号:7900)进行分析,结果根据2-ΔΔCt法进行计算。
1.4Western印迹检测 在293T细胞中完成miR-300 mimic和NC的转染过程,转染48 h后,收集细胞,完成Western印迹实验。将200 μg/孔加入酶标板中,在电压恒定为60 V的条件下,电泳0.5 h,当蛋白进入分离胶后,在电压恒定为110 V的条件下,电泳1.5 h,然后在电流恒定为80 mA的条件下,将蛋白转膜100 min至聚偏氟乙烯(PVDF)膜,然后用 5%脱脂牛奶37℃封闭 1 h,Tris盐酸缓冲液(TBS-T)洗 5 min,洗1 次;加入以抗体稀释液 1∶1 000配制的鼠抗人BRD7(Santa Cruz,USA)抗体4℃ 过夜,TBS-T洗 10 min共3次; 加入抗体稀释液 1∶4 000配制的辣根过氧化物酶(HRP)标记的山羊抗小鼠IgG,37℃作用1 h,使用TBS-T清洗3次,每次10 min,采用增强化学发光法(ECL)检测荧光度。使用抗体稀释液 1∶1 000配制的抗α-tubulin作为一抗,二抗为抗体稀释液 1∶4 000 配制的 HRP标记的山羊抗小鼠IgG。
1.5CCK-8增殖实验 实验前24 h,在HepG2和Hep3b细胞完成miR-300 mimic和NC的转染过程,第2天在96孔板中接种胰酶消化细胞(3 000个/孔),CO2培养箱温度设置为37℃,浓度为5%,培养5 d内每天每孔中加入10 μl CCK-8,培养1 h,空白孔作为对照,调零,波长设置为450 nm,采用Bioteck DR-3506全自动酶标仪测量OD值。
1.6Matrigel侵袭实验 RPMI1640 混合液〔2份基质胶(BD Biosciences,USA)和1份RPMI1640 培养基〕置于冰上。24 孔板中加入Transwell 小室(Merck Millipore,Darmstadt,德国),在每个小室中加入RPMI-1640 混合液,体积为60 μl,将500 μl 10%FBS RPMI1640培养基加入下室,CO2培养箱温度设置为37℃,浓度为5%,培养5 h。使用RPMI1640培养基配制成细胞悬液(5×105个/ml)消化转染有miR-300 mimic和NC的细胞。将200 μl 细胞悬液加入小室内,平行做3次。把24孔板放置在细胞培养箱中,培养6 h和12 h后进行小室染色固定,应用CARL ZEISS LSM 510显微镜计数,通过统计下室的细胞数量评估细胞侵袭状态。
1.7统计学方法 应用SPSS17.0软件行方差分析、t检验。
2 结 果
2.1过表达miR-300促进肝癌细胞的增殖 将miR-300 mimic和NC分别转染入HepG2和Hep3b细胞,qRT-PCR首先检测miR-300的表达水平,与转染NC比较,转染miR-300 mimic后,HepG2和Hep3b细胞中miR-300的表达水平均上升〔(4.30±0.61)vs(1.00±0.30),(5.60±0.30)vs(1.00±0.25)〕,差异具有统计学意义(t=8.521,20.403;P=0.001,0.000)。接着进行CCK-8增殖实验结果显示,在实验第3、4、5天,转染miR-300 mimic组HepG2细胞的增殖能力与NC组比较差异有统计学意义(P<0.05);同样,在实验第3、4、5天,转染miR-300 mimic组Hep3b细胞的增殖能力与NC组比较差异有统计学意义(P<0.05),提示过表达miR-300后细胞的增殖能力明显高于对照组。见表2。

表2 转染后不同时间点过表达miR-300促进肝癌细胞的增殖
与NC组比较:1)P<0.05,与第1天比较:2)P<0.05
2.2过表达miR-300促进肝癌细胞的侵袭 Matrigel侵袭实验结果发现,HepG2细胞在6 h或12 h侵袭到小室底部的细胞数,转染miR-300 mimic组多于NC组,差异有统计学意义(P<0.05)。同样,Hep3b细胞在6 h或12 h侵袭到小室底部的细胞数,转染miR-300 mimic组多于NC组(P<0.05),提示过表达miR-300明显促进HepG2和Hep3b细胞的侵袭能力。见表3。
表3 培养不同时间过表达miR-300促进肝癌细胞的侵袭
与6 h比较:1)P<0.05
2.3miR-300靶向作用于BRD7 荧光素酶双报告实验结果显示,miR-300能显著抑制pGL3-BRD7 3′UTR虫荧光素酶报告质粒的活性,差异具有统计学意义(P<0.05),见表4。进一步通过将miR-300 mimic与NC分别转染入293T细胞,利用Western印迹实验检测BRD7的表达水平,结果显示,与转染NC相比,转染miR-300 mimic后的293T细胞中BRD7的蛋白表达水平明显下调,见图1,提示miR-300通过直接靶向BRD7并抑制其表达。
2.4过表达BRD7抑制肝癌细胞的增殖过程 为了说明BRD7是否参与miR-300促进肝癌细胞的增殖过程,将BRD7质粒及对照分别转染入过表达NC和miR-300 mimic的HepG2和Hep3b细胞中进行CCK-8增殖实验。在HepG2组中,第3、4、5天,miR-300 mimic+对照质粒组细胞增殖能力与NC+对照质粒组比较,差异均有统计学意义(P<0.05);而第3、4、5天,NC+对照质粒组细胞增殖能力均高于NC+BRD7组,差异均有统计学意义(P<0.05);且第3、4、5天,miR-300 mimic+对照质粒组细胞增殖能力与miR-300 mimic+BRD7组比较,差异均有统计学意义(P<0.05)。而Hep3b中CCK-8的结果与HepG2的结果一致。以上结果说明过表达BRD7后能明显抑制肝癌细胞的增殖能力。见表5。
表4 miR-300靶向作用于BRD7比较
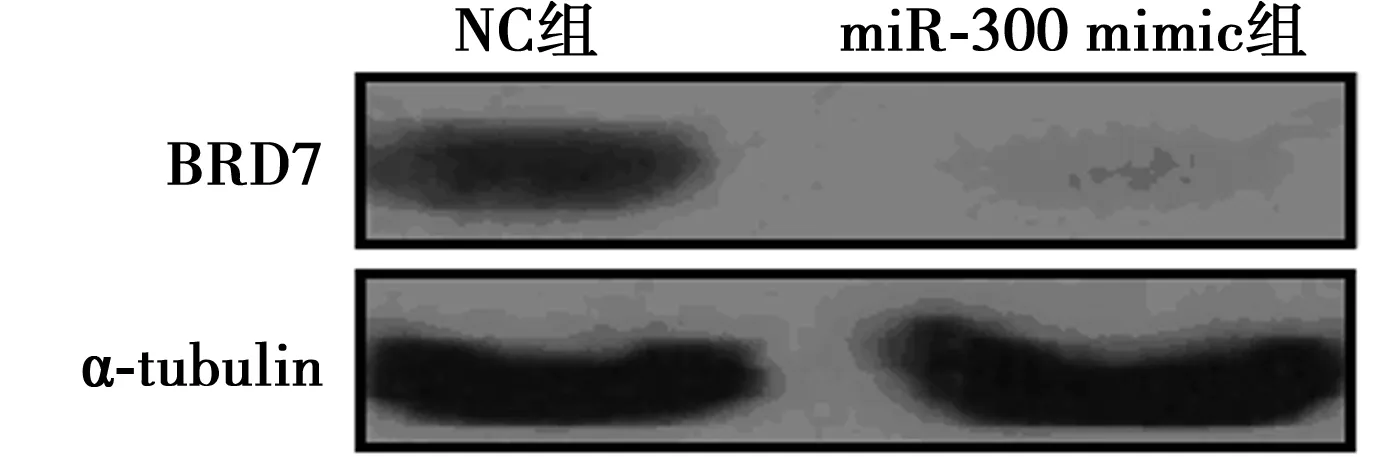
图1 Western印迹检测BRD7蛋白表达水平
表5 转染后不同时间点过表达BRD7抑制肝癌细胞的增殖过程
整体分析为两因素重复测量方差分析;组间两两比较为LSD-t检验;1)2)3)分别为与NC+对照质粒组,NC+BRDT组,miR-300 mimic+对照质粒组比较:P<0.05;时间两两比较为差值t检验;与第1天比较:4)P<0.05
2.5过表达BRD7抑制肝癌细胞的侵袭过程 为了说明BRD7是否参与miR-300促进肝癌细胞的侵袭过程,在转染入NC和miR-300 mimic的细胞HepG2中分别过表达BRD7质粒及对照质粒,进行Matrigel 侵袭实验,结果显示,无论6 h还是 12 h,过表达 BRD7 后HepG2细胞的迁移能力均明显减弱,差异具有统计学意义(P<0.05);同样,无论6 h还是12 h,过表达 BRD7后Hep3b细胞的侵袭能力均明显减弱,差异具有统计学意义(P<0.05),提示miR-300通过靶向BRD7促进肝癌细胞的侵袭能力。见表6。
表6 培养不同时间过表达BRD7抑制肝癌细胞的侵袭过程
整体分析为两因素重复测量方差分析;组间两两比较为LSD-t检验;1)2)3)分别为与NC+对照质粒组,NC+BRD7组,miR-300 mimic+对照质粒组比较:P<0.05;时间两两比较为差值t检验;与12 h比较:4)P<0.05
3 讨 论
从肝炎病毒感染到肝癌的发生包括一系列过程,如病毒导致持续的肝损伤发展为肝硬化、肝衰竭最后发展为肝癌〔5〕。证据表明,miRNAs在肿瘤发生的早期至晚期过程中都发挥重要作用〔6〕。许多特征性的miRNAs表达谱在肝脏疾病中发生变化。而对这些miRNAs作用的深入研究将为miRNAs作为肿瘤诊断、分期、预后等潜在的非侵入性的生物标记提供理论基础。本研究发现,过表达miR-300可以促进肝癌细胞的增殖和侵袭过程,且验证了BRD7为miR-300的靶基因,当过表达BRD7时肝癌细胞的增殖和侵袭过程受到抑制。以上结果得出结论:miR-300通过靶向BRD7促进肝癌细胞的增殖和侵袭过程。
研究表明,miR-300同时具有促癌及抑癌作用〔7,8〕。miR-300在神经胶质瘤中高表达,过表达miR-300可以促进胶质瘤干细胞的自我更新、增殖〔9〕。而在头颈鳞状细胞癌中和乳腺癌中miR-300表达水平较低〔10〕,过表达miR-300可以阻止乳腺癌细胞MDA-MB-231中转化生长因子(TGF)-β诱导的上皮间质转化。但在本实验发现miR-300具有促进肝癌细胞增殖与侵袭的作用,验证了BRD7为miR-300的靶基因,BRD7属于BRD家族,在心脏、肺、结肠和乳腺等多种组织中普遍表达。免疫荧光实验发现BRD7定位于细胞核中,可与乙酰化组蛋白H3结合,从而调节染色质重塑。BRD7在结肠癌和鼻咽癌中表达水平较低,过表达BRD7通过细胞周期阻滞抑制鼻咽癌细胞的增殖〔11〕;另外的研究表明,BRD7抑制前列腺癌细胞的增殖;而敲低BRD7可以促进p85α在细胞质聚集,从而稳定p110的表达水平,促进PI3K信号通路的激活〔12〕。本研究发现,miR-300可以靶向下调BRD7的蛋白水平,而过表达BRD7可以抑制肝癌细胞的增殖和侵袭过程,该发现与BRD7为抑瘤基因的结果一致,但miR-300是否通过抑制BRD7的表达间接激活PI3K信号通路发挥该作用仍需进一步研究。
猜你喜欢
成都医学院学报(2022年4期)2022-08-19
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
天津医科大学学报(2021年4期)2021-08-21
江西农业学报(2021年4期)2021-04-20
中日友好医院学报(2021年1期)2021-04-14
广东蚕业(2021年1期)2021-03-18
三农资讯半月报(2020年11期)2020-06-21
山东医药(2020年9期)2020-05-20
中华建设(2019年7期)2019-08-27
