
西部采煤沉陷地微生物复垦植被种群自我演变规律
2019-02-26 00:50毕银丽申慧慧
煤炭学报 2019年1期
毕银丽,申慧慧
(中国矿业大学(北京) 煤炭资源与安全开采国家重点实验室,北京 100083)
煤炭开采造成地表沉陷,植被原有生存环境受到扰动,矿区生态更为脆弱,植被单一,矿区生态重建迫在眉睫[1]。西部矿区干旱缺水,土壤沙化贫瘠,植被定植成为复垦成功的首要条件,植被多样性为矿区生态可持续发展奠定基础[2]。近年来我国西部采煤沉陷区采用菌根真菌等微生物技术进行大规模人工造林等重建工程,取得了较好的生态效应。菌根真菌是自然界土壤中广泛存在的一类真菌,菌根真菌是与植物共生最为密切的土壤微生物之一,陆地生态系统80%以上的植物可以与菌根真菌形成菌根共生体[3]。菌根可以扩大植物根系吸收面积,促进植物对水分和养分的吸收,从而改善植物的营养状况、加速植被恢复进程[4-5]。URCELAY和DAZ认为,接种菌根真菌可以显著改变植物群落的物种多样性[6]。O’CONNOR等对澳大利亚南部半干旱草本植物群落的研究发现,菌根真菌制约着植物群落内物种组成比例,并且显著降低了植物群落的物种多样性[7]。GRIME通过人工植被技术,增加地表覆盖度,改善土壤肥力,使植被具有自行繁殖和再生的能力[8],VAN DER HEIJDEN[9]、张涛等[10]认为接种菌根真菌促进了植物多样性。史立君[11]和胡婵娟等[12]认为微生物作为土壤重要组分,不仅可以改良退化土壤理化性质,还可以促进植物生长,提高环境安全性,对生物多样性和稳定生态系统均具有一定作用。目前大多研究集中在微生物对自然植物群落物种组成和多样性影响方面[13],而对我国西部煤矿区,气候干旱、土壤贫瘠、采煤沉陷频繁扰动,微生物复垦后不同生态群落中植物种类、数量及多样性变化的研究较少,对该人工生态系统的长期定位监测与种群演变规律更缺乏系统研究。
笔者针对我国西部典型采煤沉陷区不同种植类型樟子松、沙棘和文冠果人工林接种菌根真菌4 a后,原位监测其自然恢复过程中植被种类、生活型组成及植被多样性指数,借助ArcGIS软件及地统计学方法对物种数量进行克里金插值,研究其数量生态学特征,揭示接种菌根真菌对植被群落物种多样性和生态数量的影响规律,比较不同人工林微生物复垦后群落植被组成结构及物种多样性的变化规律,建立当地的物种指标数据库,为进一步揭示人工干预生态修复的演变规律以及接菌对生态群落结构功能变化影响提供理论依据[14-15]。
1 材料与方法
1.1 研究区概况
试验基地在陕西省榆林市神木县大柳塔镇东山煤矿开采沉陷区,处于陕北黄土高原沟壑区向毛乌素沙漠的过渡地带。所处地理坐标为东经110.28°,北纬39.30°,平均海拔1 255 m,年均气温7.3 ℃,年均降水量368 mm左右,年均蒸发量1 319 mm,降水主要集中在7~9月,属于干旱半干旱地区。主要特征是光照充足,降水稀少,蒸发量大,地表干旱,大风频繁[14]。土壤质地以风积沙土为主。人工种植前,草本植物以一年生的狗尾草、绳虫实和雾冰藜为主,灌木主要是油蒿,稀疏分布,盖度30%。2012年人工种植樟子松(株高1 m,间距3 m×2 m)4 hm2,分别设接菌彩色豆马勃(Pisolithustinctorius)和不接菌对照各一半,文冠果和沙棘(3 a生,株高1 m,间距3 m×2 m)各4 hm2,分别设接菌AM(菌种为摩西管柄囊霉(Funneliformismosseae,Nicolson & Gerdemann,简称Fm))和不接菌对照各占一半。苗木4月份种植,第1月人工浇水2次,达到饱和状态,以后免人工养护自然生长。
1.2 样地设置及植被调查
分别以4 a林龄(4 a)的樟子松、沙棘和文冠果人工林20 000 m2的接菌区和对照区为试验区,每个样区设有10个2 m×2 m的灌木层样方,和10个1 m×1 m的草本层样方,随机布置样方。按照灌木和草本分层调查林下植被种类和数量。样区的植物调查在植被生长旺盛期即2016年8月开始,每个样地上定位设置各层标准样方进行植被调查。标准样方内记录内容包括,灌草各层出现物种的种类、株数、分布状况等信息。利用手持GPS读取样本点的经纬度、海拔,观察记录样方周围的其他环境特征[7]。该区域2012—2016年的气象资料如图1所示,植被生长与降雨和气温有很大关系,种植后4 a中(2012—2016年)该区域降水量增加,引起植被种类变化。

图1 试验区2012—2016年气象资料Fig.1 Meteorological data of 2012—2016 in test area
1.3 分析计算
(1)根据Raunkiaer频度定律,物种分离频度F=(群落中某种植物出现的样方数/总样方数)×100%[16]。
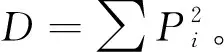
(3)接菌样区植物侵染率和菌丝密度测定。随机选择样区灌层和草本层植物根系,菌根侵染率测定采用Phillips和Hayman法染色,玻片镜检测定侵染根段数,菌根侵染率=菌根段数/被检根段数×100%。菌丝密度采用真空泵微孔滤膜抽滤-网格交叉法测定[18]。
1.4 克里金插值法
克里金法又称为空间局部估计及空间局部插值法,是建立在变异函数理论及结构分析基础上,在有限区域内对区域化变量的取值进行线性无偏最优估计的一种方法[19],包括普通克里金法、简单克里金法和通用克里金法等。一般步骤如下:
(2)变异函数分析及最优模型确定。作不同物种的半变异函数图,并分别通过对球状模型、指数模型、线性有基台值模型及高斯模型4种模型的验证。利用GS+软件,根据残差越小越好,决定系数越大越好的原则,确定最优拟合模型。
(3)克里金插值。根据确定的最优拟合模型,采用适合的克里金方法进行插值计算。
(4)交叉验证。交叉验证指的是每次从采样数据中取出一点,用其他点预测该点的值,将预测值与实测值比较后,再将该点放回原始采样数据,然后从所有样点中取出另外一个未被取出过的点,用剩余点来进行预测,如此往复,直到所有点都被取过为止。
1.5 数据处理
采用ArcGIS10.0软件、SPSS 19.0 软件、GS+软件及Excel软件进行相关数据的分析及绘图。
2 试验结果与分析
2.1 样地植物种分类特征
经过调查发现,样区林下植物包括6科16属,共18种(表1)。其中草本层有菊科、禾本科、萝藦科、藜科和豆科,共5科13种植物。灌木层有菊科、豆科和蔷薇科,共3科5种植物。其中,樟子松对照组林下包括11种植被,分属5科11属。樟子松接菌组林下包括12种植被,分属5科11属。沙棘对照组林下包括12种植被,分属5科11属。沙棘接菌组林下包括12种植被,分属4科10属。文冠果对照组林下包括10种植物,分属4科9属。文冠果接菌组林下包括12种植被,分属4科11属。
表1样地植物基本信息
Table1Basicinformationofplantsinsites

层次名物种拉丁名科属绳虫实Corispermum declinatum藜科虫实属羊草Leymus chinensis(Trin.) Tzvel.禾本科赖草属白草Pennisetum centrasiaticum禾本科狼尾草属地稍瓜Cynanchum thesiodes (Freyn).K.Schum萝藦科鹅绒藜属糙隐子草Cleistogenes squarrosa (Trin.) Keng禾本科隐子草属沙芦草Agropyron mongolicum Keng var.mongolicum (Gramineae)禾本科冰草属草本层阿尔泰狗娃花Heteropappus hispidus (Thunb.) Less菊科狗娃花属大针茅Stipa grandis禾本科针茅属苦苣菜Sonchus oleraceus L.菊科苦苣菜属猪毛蒿Artemisia scoparia Waldst.et Kit菊科蒿属白花草木樨Melilotus albus Medic.ex Desr豆科草木樨属牛心朴子Cynanchum hancockianum (Maxim.) Al.Iljinski萝藦科鹅绒藜属雾冰藜Bassia dasyphylla藜科雾冰藜属油蒿Artemisia ordosica菊科蒿属花棒Hedysarum scoparium豆科岩黄耆属灌木层达乌里胡枝子Lespedeza davurica豆科胡枝子属柠条Caragana intermedia豆科锦鸡儿属野樱桃Cornus chinensis Wanger.蔷薇科李属
2.2 人工接菌对菌根侵染率及菌丝密度的影响
由图2可知,3种人工林样地接菌组的根外菌丝密度和菌根侵染率均显著大于对照组。菌丝密度在6块样地之间均表现出显著性差异,其大小顺序是:沙M>文M>樟M>沙CK>文CK>樟CK;3种人工林的接菌处理侵染率明显高于对照处理,其顺序与菌丝密度一致。可见,人工接菌4年菌根真菌持续发挥作用,对照区域中存在着一定的土著菌根真菌。
2.3 接菌对不同人工林植被生活型影响
不同生活型反映其生存微生境的变化[13]。根据样地植物组成将生活型划分为一年生草本、多年生草本和灌木3部分。表2为各生活型的物种组成。一年生草本主要是绳虫实、雾冰藜和苦苣菜,多年生草本的有羊草等10种植物,灌木主要是油蒿等5种植物。
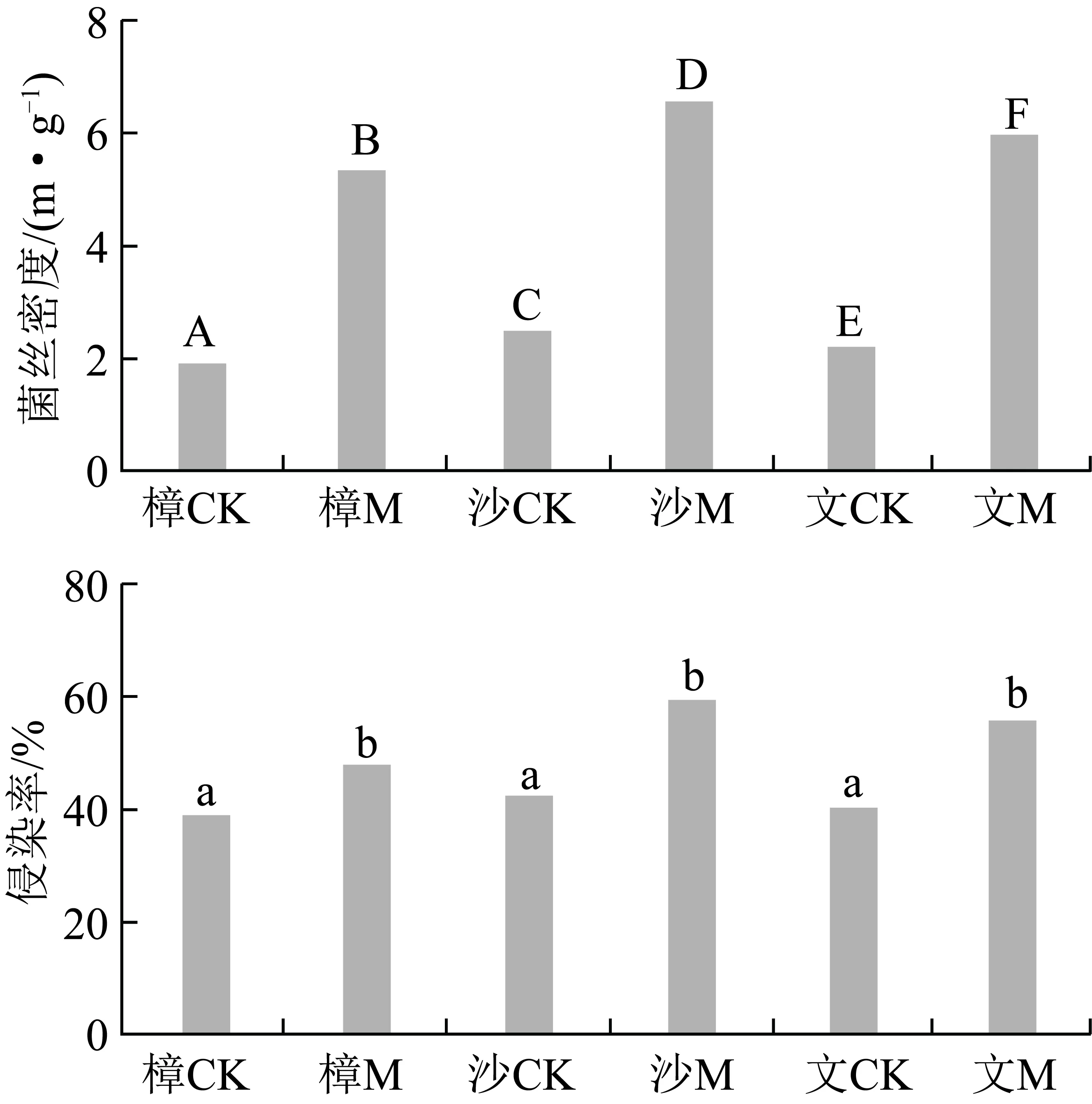
图2 不同样地的根外菌丝密度和侵染率Fig.2 External hyphae density and mycorrhizal infection rate in different sites

生活型植被一年生草本绳虫实苦苣菜雾冰藜多年生草本羊草白草地稍瓜糙隐子草沙芦草阿尔泰狗娃花大针茅猪毛蒿白花草木樨牛心朴子灌木油蒿花棒达乌里胡枝子柠条野樱桃
由图3可知,3种人工林的多年生草本种数最多,接菌组的数量均大于对照组的数量。接菌组样地内不存在一年生草本植物,而对照组仍存在1种或者2种一年生草本植物。灌木类变化不大。接菌组的菌根真菌的根外菌丝密度和根系侵染率相对于对照组较高,接菌组的多年生草本可以通过菌丝获得更多的水分和营养,利于多年生草本的入侵和定植,所以接菌组多年生草本种数大于对照组。随着时间的增加,多年生草本和灌木抢占一年生草本的生存空间和资源,多年生草本和灌木类植物根系菌根真菌的根外菌丝密度和根系侵染率相对一年生的草本植物较高,一年生草本在竞争中易被淘汰[14,20]。此外,藜科植物不能与菌根真菌形成菌根结构,属于非菌根营养型植物,菌根真菌抑制其生长[15-16],也是接菌组一年生草本退化原因之一。说明接菌组物种组成结构为多年生草本+灌木,而对照组则相对滞后,仍处于一年生草本+多年生草本+灌木的物种组成阶段。接菌促进了植物结构的演变进程。

图3 不同样地不同生活型的植物种类数量Fig.3 Plant species of different life forms in different sites
2.4 接菌对不同人工林物种频度分级
根据Raunkiaer频度定律,将频度划分为5个等级,凡频度在1%~20%的植物种归入A级,21%~40%者为B级,41%~60%者为C级,61%~80%者为D级,80%~100%者为E级。该定律得到5个频度级出现百分数的关系是:A>B>C>D 表3 不同样区频度级的百分数Table 3 Percentage of frequency bands in different sites% 因为樟子松接种的是外生菌根,而周围的草本和灌木大多感染的是内生菌根真菌,存在一定的竞争,因而表现出对照组均匀度高,而优势度接菌区高的特点。3种人工林灌木层植物油蒿的频度达到E级,分布均匀为优势种,花棒、达乌里胡枝子和柠条是常见种,野樱桃属于偶见种。 樟子松人工林对照组的优势种是羊草、地稍瓜、糙隐子草和油蒿,接菌组的优势种是羊草、地稍瓜、油蒿和花棒。沙棘人工林对照组的优势种是绳虫实、糙隐子草、沙芦草、阿尔泰狗娃花、雾冰藜和油蒿,接菌组的优势种是糙隐子草、阿尔泰狗娃花、油蒿和达乌里胡枝子。文冠果人工林对照组的优势种是羊草、地稍瓜、沙芦草、油蒿和花棒,而接菌组的优势种是白草、地稍瓜、糙隐子草和油蒿。对比3种人工林的对照组和接菌组的优势种可以发现,对照组优势种数量大于接菌组。 在接菌和对照两种处理下的3种人工林,群落植被微生物环境、种间竞争、种内斗争定会造成物种组成和群落结构差异,物种的多样性、丰富度、均匀度和优势度也随之变化。由图4可以看出,3种人工林草本层的Shannon-Wiener指数(H′)、Margalef指数(Ma)和Pielou指数(JSW)均大于灌木层,灌木层生态优势度(D)大于草本层,表明现阶段草本层占据优势地位。除了樟子松接菌组的灌木层JSW指数大于草本层。 图4 不同样地的生物多样性指数Fig.4 Biodiversity index of layers in different sites 分析樟子松的数据可知,草本层接菌组的H′指数、Ma指数和JSW指数均小于对照组,接菌组生态优势度大于对照组,灌木层则与草本层相反。说明草本层接菌组的物种多样性、丰富度和均匀度比对照组差,灌木层对照组的物种多样性、丰富度和均匀度比接菌组差。原因之一可能是,樟子松是外生菌根,与内生菌根真菌菌根的生存条件有竞争,从而对菌根真菌菌落结构及对其具有依赖性的林下植物产生抑制作用;原因之二可能是,接菌组樟子松周围1 m2内很少生长植物,一般分布的都是油蒿或者地稍瓜,在一定程度上了造成了草本层植物生态位的缺失;原因之三可能是,樟子松灌木层接菌组的植被比对照组分布均匀,且生态位缺失较少。这些原因可能造成樟子松草本层接菌组的物种多样性低于对照组,而灌木层接菌组的物种多样性大于对照组。说明接菌促进樟子松灌木层物种多样性的增大,而抑制了草本层的物种多样性增大。 沙棘和文冠果两种人工林结果一致。草本层接菌组的H′指数、Ma指数和JSW指数大于对照组,接菌组生态优势度小于对照组;灌木层接菌组的H′指数、Ma指数和JSW指数小于对照组,接菌组生态优势度大于对照组。说明接菌可以提高沙棘和文冠果人工林的草本植物群落稳定性和物种多样性及其均匀度,抑制灌木层物种多样性增加。 通过分析群落结构、生活型、频度及物种多样性,仍存在一定的偶然性和人为主观性,且仅能获得定性结果而不能得到定量指标。通过地统计学中克里金插值法估测植被数量,揭示不同人工林下,接菌干扰和自然生长两种状态下生命特征和数量生态学演替规律。 (1)采用GS+软件,确定最优拟合模型见表4,可知绳虫实等9种植物最优模型是高斯模型,沙芦草等3种植物是球状模型,阿尔泰狗娃花等2种植物是指数模型,苦苣菜等4种植物是线性有基台值模型。 (2)克里金插值。因为高斯模型所占比例最大,所以利用ArcGIS中的地统计学模块的高斯模型进行拟合,按照数量等级标准,估计不同等级的植物数量,结果见表5。由表5可知,樟子松接菌组相对于对照组,数量增加的是:羊草、地稍瓜、沙芦草、阿尔泰狗娃花、白花草木樨、牛心朴子、柠条和野樱桃;数量减少的是:白草、糙隐子草、苦苣菜、油蒿和花棒。沙棘接菌组相对于对照组,数量增加的是:羊草、白草、地稍瓜、阿尔泰狗娃花、猪毛蒿、牛心朴子、油蒿、达乌里胡枝子和柠条;数量减少的是:绳虫实、糙隐子草、沙芦草、大针茅、白花草木樨和雾冰藜。文冠果接菌组相对于对照组,数量增加的是:白草、地稍瓜、沙芦草、阿尔泰狗娃花、白花草木樨、牛心朴子、达乌里胡枝子和柠;数量减少的是:羊草、糙隐子草、苦苣菜、油蒿和花棒。 表4不同植物最优拟合模型及参数 物种最优模型C0C0+CR2C0/(C0+C)绳虫实高斯模型211.002 4170.6800.913羊草高斯模型0.10125.1000.0920.999白草高斯模型1.001 6290.5000.999地稍瓜高斯模型1.001 6760.2620.999糙隐子草高斯模型4.00320.9000.3580.988沙芦草球状模型1.001 28300.4860.999阿尔泰狗娃花指数模型3.9720.49000.806大针茅球状模型17.6074.4000.4360.763苦苣菜线性有基台值模型6.045 96.045 90.0840猪毛蒿高斯模型0.01015.7800.2460.999白花草木樨高斯模型0.0012.9150.1041.000牛心朴子高斯模型0.0106.5200.3470.998雾冰藜球状模型3.40074.2000.0470.954油蒿指数模型16.10047.9200.3310.664花棒线性有基台值模型89.25390.75900.017达乌里胡枝子高斯模型8582 509 00.4780.658柠条线性有基台值模型0.9550.955 00.3380野樱桃线性有基台值模型0.088 60.088 60.3970 注:C0,C0+C,R2和C0/(C0+C)分别表示块金常数、基台值、决定系数和块金系数。 表5不同样地物种的克里金插值 物种样地分类樟子松CK/株樟子松M/株沙棘CK/株沙棘M/株文冠果CK/株文冠果M/株绳虫实——6 078———羊草2 38421 463192 6512 157259白草1 930116—187—8 809地稍瓜2 16819 2109114 2796 70713 721糙隐子草 1 2135561 6091 4285 316967沙芦草6934117 7994 9091 3732 704狗娃花322273153 774—572大针茅——3 0262 050——苦苣菜105———26—猪毛蒿———7 281——白花草木樨—1 050464——77牛心朴子—403361 56741660雾冰藜——492———油蒿8514325295714288花棒1 112449——1 49941达乌里胡枝子——1 5533 138193 807柠条34117—262331野樱桃1463———— 注:“—”表示采样点未在发现该植物。 (3)交叉验证。笔者选择均方根和平均标准误差作为检验方程精度的评价指标,不同植物的检验方程及评价指标结果见表6。根据均方根和平均标准误差越小,精度越好的原则,可知阿尔泰狗娃花、苦苣菜、猪毛蒿、白花草木樨、牛心朴子、油蒿、柠条和野樱桃的预测值和实测值更加接近,精度较好,其中野樱桃的精度最高。其他植物的预测值与实测值偏差较大。在实际采样中,预测精度较好的植物,各采样点之间数量分布较为均匀,对克里金法插值的平滑效应影响不大,间接说明这些植物的空间分布较为均匀。其他不太理想的植物则存在样点之间数量差异大以及空间分布上的斑块化特征,使其预测值与实测值之间存在一定程度的偏差。这与实际情况相符。 表6不同植物的检验方程及评价指标 物种检验方程均方根平均标准误差绳虫实y=0.177 1x+5.472 619.522 819.077 7羊草y=0.070 5x+3.390 113.587 313.290 5白草y=0.056 3x+11.404 744.973 743.352 7地稍瓜y=-0.028 5x+28.474 243.455 643.063 2糙隐子草y=-0.049 8x+12.373 926.942 623.452 0沙芦草y=0.228 8x+6.256 826.933 622.313 4阿尔泰狗娃花y=0.080 9x+1.731 75.181 24.879 1大针茅y=0.206 4x+3.063 79.056 18.371 1苦苣菜y=-0.008 8x+0.132 92.165 81.766 9猪毛蒿y=0.062 3x+0.315 33.794 83.722 1白花草木樨y=0.034 1x+0.478 31.921 61.906 0牛心朴子y=0.025 6x+0.923 92.906 42.725 1雾冰藜y=0.006 5x+0.515 217.304 913.851 5油蒿y=0.244 0x+6.631 24.170 74.342 6花棒y=0.251 4x+1.809 18.943 39.389 2达乌里胡枝子y=0.071 1x+11.314 936.266 433.316 0柠条y=0.013 1 x+0.410 01.061 31.060 1野樱桃y=0.009 2 x+0.038 70.298 80.340 1 注:x和y分别表示实测值和预测值。 结合频度分析和克里金插值分析可知,樟子松对照组的群落结构是:羊草+白草+地稍瓜+糙隐子草+油蒿+花棒;接菌组的群落结构:羊草+地稍瓜+糙隐子草+白花草木樨+油蒿+花棒。对比樟子松的对照组,其接菌组的糙隐子草、苦苣菜、花棒的频度与数量均下降,而羊草、地稍瓜、沙芦草、阿尔泰狗娃花、白花草木樨、牛心朴子和柠条频度和数量均增加。沙棘对照组的群落结构是:绳虫实+糙隐子草+沙芦草+大针茅+雾冰藜+油蒿+达乌里胡枝子;接菌组的群落结构是:羊草+地稍瓜+糙隐子草+沙芦草+阿尔泰狗娃花+大针茅+猪毛蒿+牛心朴子+油蒿+达乌里胡枝子。对比沙棘的对照组,其接菌组的绳虫实、糙隐子草、沙芦草、大针茅、白花草木樨和雾冰藜的频度和数量均下降,而羊草、白草、地稍瓜、阿尔泰狗娃花、猪毛蒿、牛心朴子、达乌里胡枝子和柠条的频度和数量均增加。文冠果对照组的群落结构是:羊草+地稍瓜+糙隐子草+沙芦草+油蒿+花棒;接菌组的群落结构是:白草+地稍瓜+糙隐子草+沙芦草+牛心朴子+油蒿+达乌里胡枝子。对比文冠果的对照组,其接菌组的羊草、糙隐子草、苦苣菜和花棒的频度和数量均减少,而白草、地稍瓜、阿尔泰狗娃花、白花草木樨、牛心朴子、达乌里胡枝子和柠条的频度和数量都增加。另外需要指出,油蒿的频度几乎没有下降,但是其数量在樟子松和文冠果样地下降达到50%以上,原因之一可能是油蒿作为前期物种,本身就容易被其他物种更替[21];原因之二是油蒿为菊科蒿属,蒿属不能与菌根真菌形成良好共生关系[22]所致。 综上所述,笔者认为接菌可能促进地稍瓜、阿尔泰狗娃花、牛心朴子、达乌里胡枝子和柠条频度和数量的增加,可能使绳虫实、糙隐子草、苦苣菜和花棒的频度和数量下降。群落内出现暂时的生态位缺失和退化是正常的,应该尽量减少人工干扰,给物种适应环境的时间,使之自然发展。同时认为群落结构的演替,不仅与植物本身的生命特性有关系,还与植物的生活型、植物与菌根真菌的相互作用、种内种间的竞争、样地内兔子等动物的采食或者非采食活动、地形地势和气候等都有一定的关系,还有很多因子值得进一步分析和研究。 (1)3种人工林多年生草本占优势地位。接菌组进入多年生草本+灌木的阶段,而对照组则相对滞后,处于一年生草本+多年生草本+灌木的阶段。接菌提高樟子松人工林灌木层物种多样性,而降低草本层物种多样性。接菌提高沙棘和文冠果人工林的草本植物群落稳定性和物种多样性及其均匀度,抑制灌木层物种多样性增加。 (2)综合分析可知,植物的最优拟合模型以高斯模型居多。阿尔泰狗娃花、地肤、猪毛蒿、白花草木樨、牛心朴子、狗尾草、油蒿及柠条的预测值精度较好。接菌可能促进地稍瓜、阿尔泰狗娃花、牛心朴子、达乌里胡枝子和柠条频度和数量的增加,可能使绳虫实、糙隐子草、苦苣菜和花棒的频度和数量下降。 (3)本研究结果揭示了人工林在自然恢复状态和接种菌根真菌两种复垦方式下的物种组成、频度、物种多样性以及个体数量的变化及差异,对于生态系统内的其他生物与非生物因子还有待于进一步的深入监测、分析与探究。
2.5 接菌对不同人工林植被多样性影响

2.6 克里金插值在物种多样性研究中的应用
Table4Optimalfittingcurvesandparametersofdifferentspecies
Table5Kriginginterpolationofplantsindifferentsites
Table6Testequationsandevaluationindexesofdifferentspecies
3 结 论
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
西南农业学报(2022年8期)2022-10-28
亚热带资源与环境学报(2022年2期)2022-06-30
中国食用菌(2022年5期)2022-06-08
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
农民致富之友(2019年8期)2019-05-22
农民致富之友(2019年4期)2019-03-13
北极光(2018年12期)2018-03-07
现代农业研究(2017年8期)2017-09-22
