
羟自由基氧化对高白鲑肌原纤维蛋白降解的影响
2019-08-30 06:12秦军委雷用东邱恒恒朱新荣
食品科学 2019年15期
秦军委,雷用东,邱恒恒,郭 欣,朱新荣,张 建,*
(1.石河子大学食品学院,新疆 石河子 832003;2.新疆农垦科学院,新疆 石河子 832000)
我国淡水资源丰富,淡水渔业养殖具有得天独厚的优势,目前除了养殖四大家鱼外,冷水鱼越来越受到人们的关注。高白鲑(Coregonus peled)属于鲑科(Salmonidae)、白鲑属(Coregonus),自然分布于高寒地区的湖泊及河流中,是典型的冷水鱼[1]。高白鲑适应性强,且肉味鲜美、营养丰富、经济价值较高,是国内外水产市场深受欢迎的冷水鱼品种[2-3]。但是由于鱼体中肌纤维较短、水分含量较高、内源酶活性高等特点,在加工和贮藏过程中容易发生肌肉软化和肌原纤维蛋白降解严重等问题,致使鱼肉品质下降。
蛋白质是鱼类肌肉组织的主要组成成分之一,其降解、聚合和变性都会严重影响鱼肉的品质特性,营养价值和加工性能[4]。肌原纤维蛋白是肌肉中最重要的一类功能性蛋白,主要包括肌球蛋白、肌动蛋白、原肌球蛋白和肌钙蛋白等,这类蛋白在热诱导作用下可以形成三维网络结构,赋予肉制品良好的感官特性和功能特性[5-6]。蛋白质氧化是导致宰后鱼肉品质下降的另一重要因素,研究表明,氧化降低了肌肉的颜色、风味和嫩度等食用品质特性,还会导致蛋白质功能性质(如凝胶性和乳化性等)的改变[7-8]。目前对鱼肉蛋白进行氧化使用最多的是羟自由基氧化体系,氧化对象多为肌原纤维蛋白。前人对鱼类肌原纤维蛋白的研究发现,氧化可导致羰基含量和表面疏水性增加,总巯基含量、活性巯基含量、乳化活性和乳化稳定性显著下降,氧化后的肌原纤维蛋白发生交联和聚集,生成大量高分子凝集体,肌球蛋白重链(myosin heavy chain,MHC)比肌动球蛋白更容易氧化交联[9]。另外也有研究发现蛋白质氧化后肌原纤维蛋白的水结合能力和白度下降,且氧化可使蛋白发生聚集[10]。姜晴晴[11]和李学鹏[12]等均发现,氧化后MHC发生交联聚集,产生了分子质量大于200 kDa的蛋白聚集体。目前研究大多集中于氧化对肌原纤维蛋白理化特性和功能特性的影响,而关于氧化对肌原纤维蛋白降解变化以及具体蛋白如何变化的研究较少。
由于高白鲑出水即死的特点,使其只能进行冷藏运输和加工,另外,氧化对鱼肉蛋白的影响越发突显,因此研究高白鲑氧化后在贮藏条件下肌肉蛋白的变化很有必要。本实验采用羟自由基氧化体系对高白鲑背肌肉氧化后进行贮藏,并对高白鲑肌原纤维蛋白中的MHC、肌动蛋白、肌间线蛋白和肌钙蛋白T的变化进行检测,探究氧化后贮藏对其产生的影响,以期为高白鲑在加工贮藏过程中控制蛋白氧化、提升鱼肉品质提供理论参考。
1 材料与方法
1.1 材料与试剂
新鲜高白鲑购买于新疆赛湖渔业科技开发有限公司,体长(32±2)cm、体质量(750±50)g,宰杀后去鳞、去头、去尾、去皮,取其背肌肉(约1 cm×1 cm×1.5 cm)于-80 ℃冰箱备用。
鼠单克隆抗肌动蛋白(3E9) 美国Abbkine公司;鼠单克隆抗肌间线蛋白(Clone DE-U-10)、鼠单克隆抗肌钙蛋白T(Clone JLT-12) 美国Sigma公司;过氧化物酶标记的羊抗鼠二抗 美国Abcam公司;聚偏氟乙烯(polyvinylidene difluoride,PVDF)膜 美国Millipore公司;二喹啉甲酸(bicinchoninic acid,BCA)蛋白检测试剂盒 美国Pierce公司;酶放大化学发光免疫分析(enhanced chemiluminescence,ECL)试剂盒 美国Thermo Scientific公司;FeCl3、H2O2、盐酸胍、抗坏血酸、三氯乙酸(trichloroacetic acid,TCA)、2,4-二硝基苯肼(2,4-dinitrophenylhydrazine,DNPH)、乙二胺四乙酸、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、丙烯酰胺、三羟甲基氨基甲烷(Tris)等试剂均为国产分析纯。
1.2 仪器与设备
PHS-3C pH计 上海仪电科学股份有限公司;XB 3200C电子天平、XB 220A分析天平 瑞士Precisa公司;Multifuge X1R高速冷冻离心机 美国Thermo Scientific公司;Cary 50紫外-可见分光光度计 美国Varian公司;电泳和印迹转运系统、凝胶成像系统美国Bio-Rad公司。
1.3 方法
1.3.1 氧化系统的制备
氧化系统由FeCl3、抗坏血酸、H2O2、磷酸盐缓冲液(pH 6.0)组成,包括4 组不同浓度的氧化体系:1 mmol/L H2O2+0.2 mmol/L FeCl3(组1)、5 mmol/L H2O2+0.4 mmol/L FeCl3(组2)、10 mmol/L H2O2+0.8 mmol/L FeCl3(组3)、20 mmol/L H2O2+1.0 mmol/L FeCl3(组4),每组分别在室温下氧化0、15、30、60、90 min,通过测定每组鱼肉样品的羰基含量来确定最适氧化浓度。最适氧化浓度确定后,将高白鲑背肌肉组织随机分成两组,一组作为空白组,另一组置于羟自由基氧化系统,之后将样品于4 ℃贮藏0、1、7、14 d并取样,进行后续实验。
1.3.2 肌原纤维蛋白的提取
参考Chin[13]和Jiang Xinjing[14]等的方法,取适量高白鲑背肌肉,加入10 倍体积0.3 g/100 mL NaCl溶液,均质1 min,4 ℃下8 000×g离心10 min,弃上清液,重复操作一次,将沉淀溶于10 倍体积0.6 mol/L NaCl-20 mmol/L Tris-HCl缓冲液中(pH 7.0),均质5 s,于4 ℃提取1 h以充分溶解蛋白,取蛋白溶液8 mL,加入16 mL去离子水稀释上清液使蛋白沉淀,10 000×g、4 ℃离心5 min,弃上清液,重复两次,沉淀用0.6 mol/L NaCl溶解,作为肌原纤维蛋白。采用BCA蛋白试剂盒测定并调整至同一质量浓度。
1.3.3 羰基含量的测定
参照Oliver等[15]的方法并稍作修改,取1 mL 4 mg/mL的蛋白溶液于离心管中,加入1 mL 10 mmol/L DNPH,室温下反应1 h(每10 min漩涡振荡一次)后加入1 mL 20%(体积分数)TCA,4 ℃下8 000×g离心5 min,弃上清液,用1 mL乙酸乙酯:乙醇(1∶1,V/V)清洗沉淀3 次,加3 mL 6 mol/L盐酸胍溶液,于37 ℃条件下水浴15 min使沉淀溶解,反应液8 000×g离心3 min除去不溶物质,所得上清液在370 nm波长处测吸光度。
1.3.4 SDS-PAGE样品制备及分析
调整蛋白质量浓度后,按1∶1(V/V)的比例加入样品处理液(100 mmol/L Tris-HCl、质量分数4% SDS、体积分数20%甘油、体积分数10% β-巯基乙醇、质量分数0.05%溴酚蓝,pH 8.0),100 ℃水浴5 min,冷却后放入-80 ℃冰箱备用。
SDS-聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,PAGE)参照Laemmli[16]的方法并略作修改,采用质量分数12%的分离胶和5%的浓缩胶检测肌原纤维蛋白变化,上样量10 μL,浓缩胶恒压80 V,分离胶恒压160 V,SDS-PAGE完成后进行染色、脱色、成像。
1.3.5 Western-blotting分析
参考Delbarre-Ladrat等[17]的方法并稍作修改,使用质量分数10%分离胶和5%浓缩胶检测肌动蛋白、肌间线蛋白和肌钙蛋白T的变化,上样量均为10 μL,凝胶电泳时电压与SDS-PAGE一致。
凝胶电泳后,将凝胶中相应的蛋白质在转移缓冲液(48 mmol/L Tris、192 mmol/L甘氨酸、0.037% SDS、20%甲醇)中转移到PVDF膜上。转膜后,将PVDF膜移到含有5%脱脂奶粉的TBST溶液(150 mmol/L NaCl、10 mmol/L Tris-HCl、0.05% Tween-20,pH 7.5)中,于室温下封闭2 h。然后将PVDF膜分别与稀释2 000 倍体积的鼠单克隆抗肌动蛋白、稀释2 000 倍体积的鼠单克隆抗肌间线蛋白和稀释400 倍体积的鼠单克隆抗肌钙蛋白T在4 ℃条件下孵育2 h后,先用TBST在脱色摇床上漂洗3 次,每次10 min,再用TBS缓冲液(150 mmol/L NaCl、10 mmol/L Tris-HCl,pH 7.5)洗10 min。再与稀释5 000 倍体积的过氧化物酶标记的羊抗鼠二抗室温反应1 h,继续用TBST漂洗3 次,最后用ECL法显色。
1.3.6 目标蛋白的半定量分析
通过凝胶成像仪对凝胶或PVDF膜进行成像,然后通过Quantity One软件对条带光密度值进行半定量分析。MHC、肌动蛋白、肌间线蛋白和肌钙蛋白T降解情况分别通过220、42、55 kDa和35 kDa处条带灰度来表示,其降解率通过相应降解条带的灰度除以完整条带的灰度表示。
1.4 数据统计分析
本实验除SDS-PAGE和凝胶电泳外,均采用3 次平行实验。采用SPSS Statistics 19.0软件进行数据分析,采用Duncan's多重比较检验法进行显著性分析(P<0.05),采用Origin 8.5软件作图。
2 结果与分析
2.1 羟自由基氧化对高白鲑肌原纤维蛋白羰基含量的影响
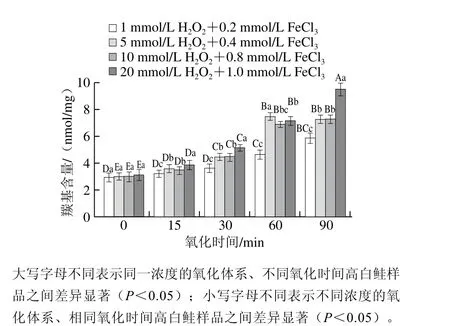
图1 不同浓度羟自由基氧化体系对高白鲑肌原纤维蛋白羰基含量的影响Fig. 1 Effect of different oxidation systems on the carbonyl content of myofibrillar proteins from Coregonus peled
为了明确合适的氧化浓度,本实验先测定了不同浓度羟自由基氧化体系中高白鲑背肌肉羰基的含量,结果如图1所示。随着氧化时间的延长,4 组不同浓度的氧化体系中的羰基含量显著升高(P<0.05),其中组1和组4在0 min时的羰基含量分别为2.93 nmol/mg和3.10 nmol/mg,在90 min时分别增长至5.90 nmol/mg和9.59 nmol/mg,分别增加了101.37%和209.35%,增长趋势显著(P<0.05)。另外,在相同氧化时间内高白鲑背肌肉羰基含量也随着氧化浓度的升高而增加,在孵育15 min之前,不同氧化浓度组的羰基含量增长差异不明显;但在30 min之后4 种组合的羰基含量变化差异显著(P<0.05)。值得注意的是,在孵育90 min时组1与组2差异显著(P<0.05),组2与组3差异不显著(P>0.05),组3与组4差异显著(P<0.05),这可能是因为蛋白质的羰基主要通过羟自由基攻击蛋白质的侧链或肽键、肽骨架的断裂、还原糖反应以及结合非蛋白羰基化合物而产生[18],也可经过分子重排直接形成;另外通过电子转移、加成和脱氢等方式使脂肪族氨基酸侧链基团转化成为烷氧自由基,并直接分解为羰基化合物。在长时间、高浓度氧化剂作用下会生成大量羰基基团,加速与亲核物质(如NH2-)的反应,最终生成席夫碱,促进蛋白质的聚合和交联,使得氧化后期羰基含量的增加速度逐渐减缓[19]。但氧化浓度过高或氧化时间过长,聚合蛋白质可能进一步发生降解和解聚,肽键断裂,羰基含量会继续增加,这可能是90 min时组4羰基含量升高的原因。
蛋白质中羰基的产生是蛋白发生氧化的一个显著性标志,可用羰基含量衡量肉制品中蛋白氧化的程度,羰基含量越高,表明蛋白质氧化程度越高[20]。综上,本实验最终采用组2氧化浓度(5 mmol/L H2O2+0.4 mmol/L FeCl3)孵育60 min的条件进行后续实验。采用SDS-PAGE分析各蛋白的降解变化情况,Western blotting分析肌动蛋白、肌间线蛋白和肌钙蛋白T的降解变化。
2.2 肌原纤维蛋白SDS-PAGE分析结果
羟自由基氧化对高白鲑肌原纤维蛋白影响的SDS-PAGE图谱如图2A所示,SDS-PAGE能够直观地反映蛋白质发生氧化后及不同氧化条件下,蛋白亚基间发生的聚集、断裂或降解等情况。经过氧化且在贮藏过程中,MHC、肌动蛋白、原肌球蛋白和肌球蛋白轻链都发生了不同程度的降解,并且氧化组降解程度比未氧化组高。在贮藏第14天时,氧化组的MHC降解率为46.39%,未氧化组为34.72%(图2B),两者差异显著(P<0.05)。特别的是,氧化组与未氧化组在贮藏1 d时MHC和肌动蛋白的条带灰度比0 d深,原因可能是在0 d时肌原纤维蛋白中的肌动球蛋白没有发生解离或者解离程度不高,分子质量过大无法进入泳道,贮藏1 d后,肌动球蛋白逐步解离为肌球蛋白和肌动蛋白,因此条带灰度深。另外可以发现,氧化组在贮藏过程中MHC和肌动蛋白的条带灰度比未氧化组低,这说明羟自由基氧化促进贮藏前期蛋白质分子发生交联或聚集,促进贮藏后期蛋白质的降解、肽键断裂及小分子化合物的降解,这可能是因为前期羟自由基影响了蛋白质结构中的氢键、二硫键、疏水作用和静电作用等,使得蛋白质的空间结构改变,活性巯基含量减少,二聚酪氨酸含量增加,促进了蛋白质的交联聚集[21-22];而后期羟自由基进一步渗透到组织内部,激活了钙激活酶类、组织蛋白酶类等内源酶,加速了蛋白的降解。

图2 羟自由基氧化体系对高白鲑肌原纤维蛋白SDS-PAGE图谱(A)和MC降解率(B)的影响Fig. 2 SDS-PAGE patterns of Coregonus peled myofibrillar proteins subjected to hydroxyl radical oxidation (A) and degradation efficiency of MHC (B)
2.3 羟自由基氧化对肌动蛋白的影响

图3 羟自由基氧化对肌动蛋白降解Western blotting图谱(A)和肌动蛋白降解率(B)的影响Fig. 3 Western blotting analysis of degradation of actin subjected to hydroxyl radical oxidation (A) and degradation efficiency of actin (B)
肌动蛋白可以组装成多种结构并广泛参与真核细胞生理功能的调节,是骨骼肌中第二丰富的蛋白质,也是肌节中细丝的主要组成成分[23]。肌动蛋白和肌球蛋白的降解能破坏肌动球蛋白的形成并导致结构完整性损伤[23]。由图3A可以看出,羟自由基氧化对肌动蛋白降解的影响不明显,氧化组与未氧化组的条带变化不明显,从图3B也可以看出,两组中肌动蛋白的降解率无明显差异(P>0.05)。只有在14 d时两组的降解率偏高,但这可能是肌动蛋白正常的降解损耗所致,贮藏前期鱼肉组织内部的内源酶开始对肌动蛋白及其他骨架蛋白进行分解,随着贮藏时间的延长,组织内部开始出现间隙,加速了蛋白质和脂质的氧化,保水性开始降低,鱼肉组织结构进一步松软解体,肌动蛋白降解加速,因此贮藏后期肌动蛋白的降解率偏高。结果表明,在高白鲑背肌肉组织中肌动蛋白的自然降解占主导地位,羟自由基氧化对其影响作用不明显。
2.4 羟自由基氧化对肌间线蛋白的影响

图4 羟自由基氧化对肌间线蛋白降解Western blotting图谱(A)和肌间线蛋白降解率(B)的影响Fig. 4 Western blotting analysis of degradation of desmin subjected to hydroxyl radical oxidation (A) and degradation efficiency of desmin (B)
肌间线蛋白是细胞骨架蛋白之一,分子质量约为53 kDa,连接于Z线与其他细胞骨架元件之间,保持肌细胞的有序性和完整性[25-26]。由图4A可以看出,氧化组的肌间线蛋白条带降解变化不明显,反而未氧化组降解明显,尤其是在贮藏后期。由图4B可以看出,未氧化组肌间线蛋白降解率在贮藏期间均比氧化组高,且一直在增加,氧化组则增长缓慢,甚至前7 d没有明显增加,这表明羟自由基氧化对鱼肉组织在贮藏期间肌间线蛋白的降解起到抑制作用,这一结果与薛梅等[27]研究牛肉肌原纤维蛋白降解的结果一致,其发现高氧化浓度下能显著抑制μ-钙激活酶对肌间线蛋白的降解。李铮等[28]的研究结果表明,肌原纤维蛋白磷酸化后肌间线蛋白的降解也受到了抑制,而去磷酸化加速肌间线蛋白的降解。这些实验结果均表明,肌间线蛋白是一类极其敏感的结构蛋白,受到外界物质影响后,其酶切位点发生改变,抑制了其降解,这也可能是细胞自我保护的一种机制。
2.5 羟自由基氧化对肌钙蛋白T的影响

图5 羟自由基氧化对肌钙蛋白T降解Western blotting图谱(A)和肌钙蛋白T降解率(B)的影响Fig. 5 Western blotting analysis of degradation of troponin-T subjected to hydroxyl radical oxidation (A) and degradation efficiency of troponin-T (B)
肌钙蛋白T分子质量约为35 kDa,是肌钙蛋白复合物的原肌球蛋白结合成分起连接作用,和肌钙蛋白I及肌钙蛋白C共同参与骨骼肌收缩与舒张的调节[29]。由图5可以看出,随着贮藏时间的延长,氧化组与未氧化组中的肌钙蛋白T条带灰度均变浅,降解率增加。但在贮藏前期,两组肌钙蛋白T的降解率差异并不显著(P>0.05),而在后期氧化组中肌钙蛋白T的降解率要显著低于未氧化组(P<0.05),这说明羟自由基氧化抑制了鱼肉组织中的肌钙蛋白T的降解,特别是在贮藏后期。前人研究表明,肌钙蛋白T是μ-钙蛋白酶的降解底物,且其在宰后易发生降解,从而改善肉的嫩度[30]。这与本研究结果有一定的差别,分析其原因可能是研究对象不同,前期研究对肌原纤维蛋白体外进行氧化,本实验则是对鱼肉组织进行氧化然后贮藏,体内复杂的内环境和各种抑制酶的存在导致肌钙蛋白T前期降解缓慢,氧化组中肌钙蛋白T降解率低于未氧化组也说明氧化对其产生了影响,降低了其前期的降解速率。
3 结 论
采用羟自由基氧化体系对高白鲑背肌肉进行氧化后,其羰基含量随着氧化时间的延长和氧化浓度的增加而升高。氧化后进行贮藏,氧化组MHC的降解率比未氧化组高,表明氧化显著促进了MHC在贮藏期间的降解;两组中肌动蛋白的差异变化不显著,自然降解占主导地位;氧化组中肌间线蛋白和肌钙蛋白T的降解率低于未氧化组,氧化抑制了肌间线蛋白和肌钙蛋白T降解,尤其是贮藏后期,对肌间线蛋白降解的抑制效果尤为显著。高白鲑鱼肉蛋白的降解变化势必影响其结构发生改变,从而影响其功能性质,所以高白鲑在贮存及加工过程中应该控制其氧化,减少鱼肉品质的劣变。
猜你喜欢
今日农业(2022年3期)2022-11-16
食品工业科技(2022年17期)2022-08-27
中国典型病例大全(2022年11期)2022-05-13
食品科学(2020年11期)2020-07-13
中国畜牧杂志(2020年1期)2020-01-16
河南水产(2019年4期)2019-12-13
水产科学(2017年5期)2017-12-18
浙江农业科学(2016年11期)2016-05-04
中国医科大学学报(2015年10期)2015-03-01
食品工业科技(2014年23期)2014-03-11
