
高原蚕区家蚕茧层率遗传分析
2020-06-29 12:09任晓晓罗朝斌孙运朋杨万军吴康云
农学学报 2020年6期
任晓晓,罗朝斌,孙运朋,杨万军,吴康云
(贵州省农业科学院蚕业研究所,贵阳 550006)
0 引言
家蚕的全茧量、茧层量、茧层率等茧质性状是蚕业生产中的重要经济性状,决定了该行业的产量和经济效益[1]。蚕桑产业作为中国的传统产业之一,在中国发展已有几千年的历史。中国早在20 世纪70 年代就成为了世界上最大的生丝生产和出口国,但出丝率低仍然是当今蚕桑产业提质增效的阻碍之一[2-3]。茧层率是决定蚕茧出丝率的基本要素,因此选育高茧层率的家蚕实用品种,一直是家蚕育种工作者的重要目标[4]。研究并了解茧层率的遗传规律,是提高家蚕茧层率水平、加快育种步伐的必要前提[5]。
前人已围绕茧层率性状做过部分研究,并阐明了一些基本的规律。钟伯雄等[6]通过性连锁模型分析茧层率的遗传规律,结果表明茧层率性状受到加性效应、显性效应和性别效应的控制,其中加性效应作用最大。司马杨虎等[7]研究发现同一品种家蚕茧层率性别效应极显著,对该性状QTL(quantitative trait loci)定位分析有较大影响。李斌等[8]对家蚕重要经济性状进行了多重区间作图分析,结果检测出3 个对茧层率贡献较大的QTLs,其加性效应为负效应。艾均文等[9]对16个夏秋用家蚕品种茧层率性状的遗传力进行了分析,结果显示茧层率遗传力较高,可在早期世代进行定向选择,而Malik等[10]的研究结果显示茧层率的遗传力和遗传进度较低,对性状直接选择以改良品种的效果较差。张烈等[11]研究表明茧层率遗传稳定性较高,且雄性较雌性更稳定。
目前,关于家蚕茧层率多是基于经典数量遗传学理论开展的遗传效应、遗传力参数等的研究。而数量性状的遗传机制比较复杂,且其性状表现容易受到环境的影响,试验材料、饲养环境等因素可能导致研究结果的差异[12]。为验证并进一步探讨茧质性状的遗传规律,本研究在贵州高原蚕区选用不同于前人研究的家蚕实用品种,并配置子一代和回交一代群体,对茧层率等性状的遗传规律进行分析,以期为茧层率数量性状位点定位分析及高茧层率品种选育提供参考。
1 材料与方法
1.1 供试材料
亲本为生产上推广使用的4个家蚕实用品种,分别为属于中系的‘菁松’、‘芙蓉’和属于日系的‘皓月’、‘湘晖’,其中‘菁松’和‘皓月’属于多丝量品种,‘芙蓉’和‘湘晖’属于中丝量品种。4个亲本均经过多代同蛾区交配,遗传背景接近于纯合,由贵州省农业科学院蚕业研究所提供。杂交种为‘菁松’、‘皓月’分别与‘芙蓉’、‘湘晖’进行正(中丝量×高丝量)、反(高丝量×中丝量)杂交获得的F1代。回交群体(B1、B2)为‘芙蓉’×(‘芙蓉’ב皓月’)、‘皓月’×(‘芙蓉’ב皓月’)。
1.2 养蚕时间及地点
2019 年春、夏、秋蚕饲养在贵州省蚕业研究所遵义蚕桑基地进行。
1.3 试验方法
1.3.1 家蚕饲养及主要经济性状调查2019年春季,饲养家蚕亲本‘菁松’、‘皓月’、‘芙蓉’和‘湘晖’,出蛾后杂交获得F1代和亲本同蛾区交配蚕种。同年夏季,将亲本和F1同室同条件催青,每个品种或组合饲养一区,每区收蚁0.35 g。同环境良桑饲养,待上簇6 天后采茧,第7 天削茧进行茧质调查,每小区随机选择雌、雄各40粒蚕茧,采集个体的全茧量、茧层量、蛹体重等表型值[13],出蛾后,以茧层率性状和血缘关系差异较大的‘皓月’与‘芙蓉’为亲本配置回交一代。2019年秋季,催青结束后将2个回交群体各收蚁0.35 g,之后同环境同条件下饲养,上簇结茧第7 天每小区随机选择雄性120 粒蚕茧,采集个体的全茧量、茧层量、蛹体重等表型值。计算个体茧层率和8 个杂交组合的杂种优势,运用多世代联合分析方法分析家蚕的茧层率等重要经济性状的遗传规律。
1.3.2 统计分析 运用Excel 对样本的各性状值进行描述统计;亲本间的经济性状差异分析通过Excel 中的T-test 进行;运用SPSS 软件对亲本和回交群体的茧层率频数分布进行分析;运用公式(1)计算8 个杂交组合的杂种优势;通过Excel 中的CORREL 公式对茧层率与全茧量、茧层量之间的相关性进行分析。
2 结果与分析
2.1 亲本及杂交组合茧层率等性状统计
由表1 可以看出,在供试的4 个亲本中,‘菁松’、‘皓月’的茧层率均大于‘芙蓉’、‘湘晖’,其中‘皓月’雄性的茧层率最高,达到了25.80%,其次为‘菁松’雄性,茧层率为25.04%,‘湘晖’的茧层率最低。对亲本的茧层率进行差异显著性分析,结果表明,无论雌雄,‘菁松’、‘皓月’的茧层率与‘芙蓉’、‘湘晖’相比,差异均达到了极显著水平(图1)。
家蚕的茧质性状具有明显的性别效应,其中同一家蚕品种雌性的全茧量、茧层量、蛹体重均大于雄性,而雄性的茧层率大于雌性。F1代的全茧量、茧层量、茧层率和蛹体重等表型值均介于两亲本之间,4 种茧质性状的高值对低值为显性。反交组合‘菁松’ב芙蓉’、‘菁松’ב湘晖’、‘皓月’ב芙蓉’、‘皓月’ב湘晖’的茧层率均高于其对应的正交组合‘芙蓉’ב菁松’、‘湘晖’ב菁松’、‘芙蓉’ב皓月’、‘湘晖’ב皓月’,茧层率表现偏向于母本(表1)。
2.2 亲本和回交群体茧层率性状频数分布
为避免性别效应对茧层率数据分析的影响,对亲本‘皓月’、‘芙蓉’和回交群体‘芙蓉’×(‘芙蓉’ב皓月’)、‘皓月’×(‘芙蓉’ב皓月’)雄性个体的茧层率频数分布进行分析,结果见表2。由表2可以看出,亲本‘皓月’、‘芙蓉’的茧层率呈连续性变化,属于典型的数量性状。对两亲本、回交群体的茧层率分布频数进行正态性检验,结果显示‘皓月’(P=0.184>0.05)、‘芙蓉’(P=0.458>0.05)的茧层率符合正态分布;回交群体‘皓月’×(‘芙蓉’ב皓月’)(P=0.002<0.05)的频数呈偏态分布且只有单峰,‘芙蓉’×(‘芙蓉’ב皓月’)(P=0.001<0.05)的频数分布呈现双峰。
2.3 茧层率与全茧量、茧层量的相关性分析
对4 个亲本的茧层率与全茧量、茧层量的相关性进行分析,结果如表3所示。由表3可见,亲本的茧层率性状与茧层量呈正相关,且均达到了极显著水平;除‘湘晖’的茧层率与全茧量呈显著正相关外,其余亲本茧层率与全茧量呈负相关。
2.4 子一代杂种优势率估算
对8个杂交组合的全茧量、茧层量、茧层率和蛹体重等经济性状的杂种优势率进行计算,结果如表4 所示。由表4 可以看出,不同杂交组合同一性状间的杂种优势率差异较大,其中杂交组合‘皓月’ב芙蓉’的茧层量、全茧量、蛹体重的优势率最大,分别为23.78%、23.14%、22.69%。全茧量、茧层量、蛹体重的杂种优势较大,优势率在5.10%~23.78%之间;除‘芙蓉’ב皓月’、‘湘晖’ב皓月’的茧层率杂种优势率为负值外,其余均为正值,优势率在-3.82%~5.63%之间,杂种优势较小或几乎没有杂种优势。上述4种重要的经济性状,中系与日系杂交的杂种优势率较大,其次为中系×中系,日系×日系的杂种优势率最小。
3 结论与讨论
本研究对贵州高原蚕区4个家蚕品种和8 个杂交组合的茧层率、全茧量、茧层量和蛹体重进行了统计分析,试验结果显示上述茧质性状的表现与性别相关,这与谭远德等[7]、司马杨虎等[14]的研究结果一致,表明家蚕性染色体上可能存在控制全茧量、茧层量、茧层率和蛹体重等数量性状的位点。2013 年,侯成香等[15]利用SSR(simple sequence repeat) 标 记 和 SNP(single nucleotide polymorphism)标记对家蚕性染色体上与茧、丝相关的QTL 进行了定位研究,结果在性染色体上扫描到了与全茧量、茧层量有关的QTL 位点,其中茧层量相关QTL 的LOD值在10 以上,但并未检测到与茧层率相关的QTL。

表2 亲本及回交群体茧层率频数分布 %
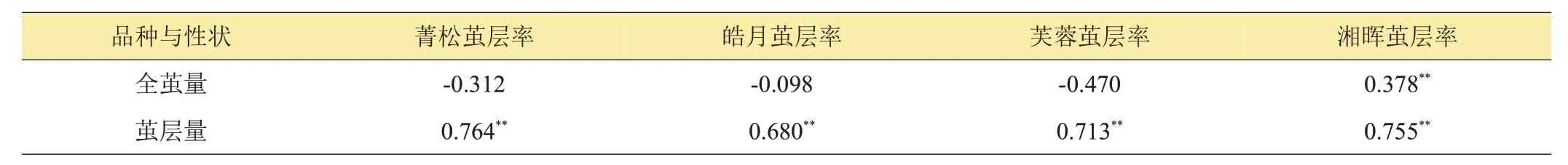
表3 不同家蚕品种茧层率与全茧量、茧层量的相关系数

表4 8个杂交组合杂种优势率 %
数量性状遗传分析表明,由于多基因和环境修饰作用,亲本和子一代等不分离世代的表型分布呈典型的正态分布[16-17]。2018年,李春林等[18]对多份家蚕种质资源的茧层率频数分布进行了分析,结果显示茧层率的频数分布表现为明显的多峰,不服从典型的正态分布,其认为这是由于样本中性状表现较差的地方品种影响了频数分布结果。但在本研究中,对‘皓月’、‘芙蓉’雄性个体的茧层率频数分布的正态性检验,结果显示两者茧层率呈典型的正态分布。2009年,司马杨虎等[19]对家蚕茧层率、茧层量等数量性状的频数分布分析结果表现为明显的双峰,但其对两性状作了性别效应调整后,频数分布接近于正态分布。因此分析认为,李春林在分析家蚕种质资源茧层率的频数分布时,并未将雌雄个体分开,性别效应或许也是非正态分布的影响因素之一。
最早研究者们会根据回交群体、子二代群体等分离世代的性状频数分布,直观地判断控制数量性状的遗传体系是单纯的多个微效基因还是存在主基因[20-21]。一般认为,若该性状由纯粹的多基因控制,其频数分布呈对称的单峰,服从典型的正态分布;若偏离正态分布或呈现多峰,则可能存在主基因[17,21]。本研究中,回交一代‘皓月’×(‘芙蓉’ב皓月’)的频数呈偏态分布的单峰,‘芙蓉’×(‘芙蓉’ב皓月’)呈现双峰,由此推断,控制茧层率性状的并非是纯粹的多个微效基因,可能存在主基因。
家蚕性状之间的相关关系是家蚕育种工作的重点内容之一,根据各性状之间的相关性,有望达到通过一种表型值以选择多个性状的目的[22-23]。本研究对4 个家蚕品种的茧层率与全茧量、茧层量的相关性进行了分析,结果显示茧层率与茧层量呈极显著正相关,这与钱荷英等[24]的研究结果相符。而本研究中,除‘湘晖’外,其余3个品种的茧层率与全茧量呈负相关,这与朱水芬等[24]、钱荷英等[25]的显著正相关结果不一致,分析认为这可能与家蚕品种、样本数量、家蚕饲养环境与条件等有关。例如本研究中的供试家蚕来源并饲养于贵州高原,与江苏、云南等其他蚕区的海拔、气候都有较大差异,这是否是影响试验结果的原因,还需进一步研究。要确定家蚕各经济性状之间的相互关系,还应加大样本品种数量、在高原和平原蚕区多次饲养研究。
家蚕是农业生产中最早利用杂种优势的物种之一,与双亲相比家蚕杂交种在适应性、抗病性、茧丝产量等方面都有较大优势[26-27]。本研究对8 个杂交组合的4 种茧质性状的杂种优势率进行了估算,结果显示中系与日系杂交的杂种优势大于中系×中系、日系×日系,这说明杂交亲本之间的血缘关系越远,杂种优势越大;茧层率的杂种优势率很小,因此不能依靠杂种优势获得高茧层率的性状表现,而只能通过培育高茧层率的家蚕纯种,但由于茧层率性状的遗传进度较低[10],所以控制茧层率性状的QTL 精细定位和分子标记辅助育种应是下一步的研究重点。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
四川蚕业(2022年2期)2022-11-19
海外文摘·文学版(2022年4期)2022-04-14
蔬菜(2021年7期)2021-11-27
四川蚕业(2021年2期)2021-03-09
四川蚕业(2021年1期)2021-02-12
作文评点报·低幼版(2018年17期)2018-07-12
校园英语·下旬(2017年12期)2017-12-09
作文大王·低年级(2017年2期)2017-02-16
江苏农业科学(2015年1期)2015-04-17
