
植物转录因子TIFY家族蛋白结构和功能的研究进展
2020-12-21 09:19杨锐佳张中保吴忠义
生物技术通报 2020年12期
杨锐佳 张中保 吴忠义
(1. 北京市农林科学院北京农业生物技术研究中心,北京 100097;2. 首都师范大学生命科学学院,北京 100048)
转录因子(Transcription factors)是RNA聚合酶在真核生物特异启动子上起始转录所需要的作用因子,由DNA结合区(DNA-binding domain,DBD)、转录调控区(Tanscription regulation domain,RD)、寡聚化位点(Oligomerization site,OS)和核定位信号区(Nuclear localization signal,NLS)组成。TIFY蛋白是植物中一类特异性转录因子,曾被称为花序分生组织中表达的锌指蛋白(Zinc-finger protein expressed in inflorescence meriste,ZIM)家 族,属于植物特异的GATA转录因子家族,后因其存在一个高度保守的功能结构域TIFY域,即TIF[F/Y]XG氨基酸序列,所以被命名为TIFY基因家族[1]。TIFY只发现于陆生植物中,在拟南芥中首次鉴定出了TIFY基因ZIM(AT4G24470)[2],随后TIFY基因陆续在水稻、杨树和小麦等中被鉴定出来[3]。根据保守结构域的组成将TIFY家族蛋白分为4个亚家族:TIFY、JAZ、ZML和PPD,且可分成两个亚群,亚群I包含C2C2-GATA结构域,亚群II不含该结构域。本文通过回顾近年研究成果,从TIFY蛋白的结构特征、分布及在调控植物的生长发育,响应各种逆境胁迫和不同激素信号等方面的生物学功能进行综述,旨在对TIFY蛋白家族深入的研究与利用奠定理论基础。
1 TIFY蛋白的结构特征
根据保守结构域的组成可将TIFY家族蛋白分为4个亚家族:TIFY、JAZ、PPD和ZML(图1)。其中TIFY蛋白包含高度保守的TIFY结构域,此结构域含28个氨基酸和一个核心基序TIF[F/Y]XG,预测分析表明TIFY结构域二级结构为α-α-β[4]。JAZ蛋白为不含DNA结合结构域的阻遏蛋白,因此必须依赖于蛋白间的相互作用来行使功能[1],除包含TIFY结构域外,在其C端还存在一个保守Jas结构域,由12-29个残基组成,可与MYC2蛋白发生互作,能参与到JA信号传导途径中,还可以与MYB[5]、EIN3、EIL1等转录因子互作,并且其Jas结构域中存在细胞核定位信号,使JAZs具有核定位特性[6]。组成PPD蛋白的3个结构域中除了保守的TIFY结构域,还存在一个N端PPD结构域以及PY基序缺失的Jas结构域,PPD蛋白不能结合DNA,但是可以依赖TIFY结构域形成同源或者异源二聚体。ZML蛋白也是除TIFY结构域,还包含有一个CCT基序、GATA锌指结构域,其中CCT结构域最早是在转录因子TOC1和CONSTANS(CO)蛋白中发现的,这两种蛋白被认为参与了植物的光周期信号传导,而CCT结构域参与了介导蛋白质与蛋白质间的相互作用[4]。

图1 TIFY蛋白家族的分类及结构特征[7]
2 TIFY蛋白在植物中的分布
TIFY家族蛋白在植物中的分布十分广泛。目前,在拟南芥中发现有18个TIFY家族基因[1]。水稻中有20个TIFY家族基因,其中9个基因对于茉莉酸和损伤处理会有应答,而几乎所有水稻TIFY基因都可以响应一种或多种的非生物胁迫,如干旱、盐碱、低温[8]。玉米中有30个TIFY家族基因,包括1个TIFY、3个ZML和26个JAZ基因,部分基因可响应生物或非生物胁迫[9]。二穗短柄草中的TIFY基因对于干旱、盐碱、低温和高温等一种或多种非生物胁迫也都有应答[10]。此外,在一些其他植物如小麦(Triticum aestivum)[3]、菜豆(Phaseolus vulgaris)[11]、丹 参(Salvia miltiorrhizaBunge)[12]、番茄(Solanum lycopersicum)[13]、香蕉(Musa acuminate)[14]、野大豆(Glycine soja)[15]、木豆(Ca-janus cajan)[16]、柳 枝 稷(Panicum virgatumL.)[17]、毛竹(Phyllostachys edulis)[18]、苹果(Malus×domestica)[19]、西瓜(Citrullus lanatus)[20]、沙梨(Pyrus pyrifolia)[21]、葡 萄(Vitis vinifera)[22]、牵 牛 花(Ph-arbitis nil)[23]、雷蒙德氏棉(Gossypium raimondii)[24]等植物中均能找到TIFY家族转录因子。从上述植物中分别挑选出一些代表性的基因进行进化关系分析,共选取不同物种的142个TIFY家族基因,其中拟南芥18种、玉米30种、水稻20种、小麦40种、二穗短柄草4种、葡萄19种、番茄5种、苹果4种及梨3种,得以展示TIFY家族在植物中的广泛分布及亲缘关系(图2)。可以发现在TIFY的4个不同亚家族中,JAZ家族属于分布最广泛及数目最多的一个家族。
3 TIFY蛋白的生物学功能
3.1 TIFY在植物生长发育中的功能
TIFY家族在植物中分布广泛,其4个亚家族在调控植物的茎、叶、花的发育过程中起到重要作用。
3.1.1 TIFY调控茎叶的发育 过量表达OsTIFY11b的水稻植株,通过增加植物茎和叶鞘中碳水化合物的积累,不仅在抽穗期叶片较长,叶鞘和茎中的淀粉和蔗糖含量高于未转化植株,其籽粒的长度、宽度和厚度也均超过未转化植株,籽粒重增加了9%-21%[25]。在拟南芥中,AtTIFY4a(PPD1)和AtTIFY4b(PPD2)基因调控叶片的发育,表现在调节叶片的大小、限制叶片边缘的弯曲程度[26]。另外有研究发现拟南芥中PPD2能与NINJA共同调节叶片的整齐度[27]。拟南芥中另外一个基因AtTIFY1(ZIM)可促进叶柄和下胚轴的延伸[28]。还有一些植物中,如雷蒙德氏棉GrTIFY基因参与纤维的发育[24]。沙梨中发现其PpTIFY基因在叶片中的表达量最高,而在果实成熟过程中表达量明显下降,并且用茉莉酸和光照处理还能诱导PpTIFY,与沙梨果皮中花青素的积累有关[21]。

图2 植物中部分TIFY家族基因聚类分析
3.1.2 TIFY调控花的发育 拟南芥ZIM(AT4G 24470)是最早被鉴定的TIFY家族成员,研究表明其与花序的发育和开花有关[2]。牵牛花的PnFL-2蛋白含有TIFY家族典型的TIFY与CCT结构域,研究发现与野生型相比,组成型过表达PnFL-2的转基因植株在长日照条件下的开花时间稍早,表明PnFL-2可能在花诱导发挥作用[23]。另外过表达SlJAZ2的转基因番茄株高和节间长度降低,毛状体减少,侧芽萌发早,花期提前,控制开花时间的基因明显上调,加速植物生长向生殖生长的转变[29]。
3.2 TIFY在植物逆境胁迫应答中的调控
TIFY家族响应生物胁迫的相关报道较少,主要集中在TIFY家族的非生物胁迫响应。
3.2.1 TIFY响应干旱胁迫 在拟南芥中JAZ7基因可通过调节植物的光合作用、氧化还原、氨基酸、植物激素和防御代谢物来产生耐旱性[30]。Li等[19]根据苹果基因组序列的预测基因共鉴定了30个TIFY基因,包括18个JAZ、2个PPD、2个ZML和8个TIFY基因,发现其中一些特定基因对非生物胁迫或激素处理有响应,如MdJAZ1、9、11、13、15和MdPPD1在干旱和高盐处理时均上调;外源脱落酸(Abscisic acid,ABA)和茉莉酸(Jasmonic acid,JA)处理时也均能诱导苹果JAZ基因显著上调表达,相比之下,大部分JAZ、PPD和ZML基因对水杨酸(Salicylic acid,SA)和乙烯(Ethylene,ET)没有响应。同样在葡萄中VvTIFY也受JA与ABA诱导,而对SA与ET不敏感,而且其中一些特定基因,如VvJAZ4、5、9、11和VvZML1表现出对冷胁迫有响应同时也都受到干旱或盐碱的调控,这意味着这两种胁迫途径在植物中可能存在相互作用。只有一个葡萄的TIFY基因(VvJAZ11)表现出对热胁迫的下调响应[22]。Zhang等[9]通过芯片数据发现,ZmTIFY基因参与玉米不同阶段发育,具有组织特异性表达模式,同时响应干旱胁迫。RNA-seq和qPCR分别鉴定了11个、13个响应脱水和冷胁迫的毛竹家族基因PeTIFY[18]。有实验发现,在水稻中过量表达OsTIFY11a基因时,可以显著性的提高水稻对于盐胁迫和脱水胁迫的耐受性[8]。OsJAZ1可作为OsbHLH148相关的茉莉酸信号通路的转录调控因子,在茉莉酸盐信号通路中发生相互作用,导致水稻耐旱[31]。
3.2.2 TIFY响应渗透胁迫 番茄的转录组数据表明,多数SlJAZ基因对盐、渗透胁迫或JA、ABA处理均有响应,如SlJAZ7和SlJAZ11可同时被JA和ABA上调,SlJAZ1、SlJAZ3和SlJAZ36分别被JA上调和ABA下调。盐对多数SlJAZ基因的表达具有正向和负向调节作用,如SlJAZ3、SlJAZ7和SlJAZ10在番茄叶片中被盐诱导上调,而抑制SlJAZ1的转录,并在根和叶中轻微地抑制SlJAZ6的转录。渗透胁迫主要抑制SlJAZ基因的表达,在番茄根中8个SlJAZ基因中的6个被渗透胁迫下调。此外,叶片中SlJAZ1、SlJAZ3、SlJAZ10的转录也受到抑制,SlJAZ7是唯一被渗透胁迫上调的基因[13]。
3.2.3 TIFY响应盐胁迫 TIFY转录因子涉及物种较多,而报道文章主要集中于TIFY表达调控分析,有关TIFY基因家族具体生物学功能的实验研究报道较少,其中有关拟南芥和大豆中TIFY响应盐胁迫的相关实验支撑较多。Zhu等[15]发现重碳酸盐、盐胁迫和植物激素JA均可诱导野生大豆GsTIFY10的表达,在拟南芥中过表达GsTIFY10可增强野生大豆在种子萌发、幼苗和成年早期幼苗发育阶段时植物对碳酸氢盐的抗性;并且在JA存在下,过表达GsTIFY10的拟南芥植物根系比野生型长,部分JA响应和JA生物合成相关基因在GsTIFY10过表达拟南芥株系中被抑制。朱丹等[32]发现在拟南芥中基因GsTIFY11b通过调控盐胁迫信号通路中关键基因(NHX1、KIN1和DREB)的转录来增加拟南芥对盐胁迫的敏感性,即降低了拟南芥对盐胁迫的耐性,表明该基因参与并干扰了植物在应对盐胁迫逆境过程中的防御机制。阎文飞等[33]采用荧光定量PCR技术检测GsTIFY6B在野生大豆不同部位以及盐碱胁迫和激素处理下的转录水平,发现GsTIFY6B可能通过参与ABA和MeJA信号通路响应盐碱胁迫。另有研究发现,隶属于TIFY家族的JAZ亚家族成员GsJAZ2可提高大豆碱胁迫耐受性,过表达GsJAZ2的转基因大豆在碱性条件下生长良好,野生型植株出现萎黄病、生长受阻甚至死亡。另外,转基因植株的叶绿素浓度高于野生型,丙二醛(MDA)含量低于野生型。大豆中GsJAZ2过表达导致JA不敏感,包括JA介导的根系生长抑制。在碱性胁迫下检测并改变了部分JA应答基因的转录本,结果表明GsJAZ2过表达可能参与了JA信号通路中抗渗透物质的积累,从而防御碱性胁迫[34]。还有研究发现在小麦中过表达TdTIFY11a的转基因株系,在高盐条件下的发芽率和生长速率均高于野生型[3]。在茉莉酸(JA)和盐、干旱等非生物胁迫处理下的qRT-PCR结果分析显示,西瓜部分TIFY基因均有应答,其中ClJAZ1和ClJAZ7的诱导表达上调最为显著[20]。
3.2.4 参与其他胁迫响应 Sirhindi等[16]发现,木豆部分CcTIFY家族基因能响应铜胁迫。玉米的ZmJAZ基因呈现出对某些生物胁迫真菌(如丝黑穗、茎腐病和炭疽病)具有一定程度的胁迫响应[9]。
3.3 TIFY参与植物多种激素信号转导网络调控
TIFY家族中的JAZ蛋白作为茉莉酸途径的抑制因子,不仅是调节茉莉酸激素应答的关键枢纽,而且可以通过和其他信号通路中调控因子的相互作用,在复杂激素调节网络中发挥重要作用[12]。JAZ蛋白可以和多种转录因子及共阻遏蛋白结合,便将茉莉酸信号通路和其他多种信号通路联系起来,有研究表明JA介导的JAZ蛋白转录水平的变化与生长素(Auxin,IAA)、ET、赤霉素(Gibberellins,GAs)及油菜素内脂(Brassinosteroid,BR)等激素信号的调节相关,从而在植物的生长发育、激素应答、胁迫响应等方面发挥作用[35]。当植物处于正常生长条件时,其体内的JA-Ile含量较低,MYC2作为JA应答基因的起始转录因子,会被JAZ蛋白结合同时吸引NINJA和TPL这些共阻遏物来共同抑制JA应答基因的转录,导致植物无法进行JA应答反应。但是当植物在生长发育过程中遇到不利环境等胁迫时,其体内的JA-Ile含量就会增多,JAZ蛋白也会被COI1结合通过泛素化蛋白酶体的途径被降解,这样MYC2等转录因子便可以顺利启动JA应答基因的转录(图3)。在苹果中的研究发现,MdCIbHLH1在体外和体内与MdJAZ1/4和MdMYC2相互作用。MdJAZ1/4作为抑制因子,在茉莉酸信号通路中结合MdMYC2。愈伤组织中MdMYC2的过表达增加了MdCIbHLH1、MdCBF1、MdCBF2和MdCBF3的表达水平,导致耐冻性增加。此外,MdJAZ1或MdJAZ4在转基因红肉苹果愈伤组织中的过表达减弱了MdMYC2对耐寒性的促进作用[36]。从茶树基因组数据库中鉴定了12个JAZ基因,RT-PCR分析显示,其中7个CsJAZ基因在根中优先表达。此外,冷、热、PEG、MeJA和GA刺激对CsJAZ的表达也有差异[37]。有研究发现,玉米ZmJAZ14基因在拟南芥中的过表达增强了植株对JA和ABA处理的耐受性以及PEG胁迫,而在GA刺激下促进了植株的生长,ZmJAZ14是JA、ABA和GA信号通路之间的交叉通讯枢纽[38]。黄丝瓜藓PnJAZ1介导JA与ABA在盐胁迫下的信号交谈[39]。棉花中GhJAZ3与GhSLR1的相互作用介导了JA和GA对表皮细胞分化与伸长的调控[40]。水稻OsJAZ9与SLR1之间的蛋白互作参与介导应激响应的JA信号与促生长GA信号之间的拮抗作用[41]。茉莉酸甲酯(Methyl jasmonate,MeJA)处理草莓果实接种灰霉病菌后防御系统启动及MYC2、JAZ1基因表达上调[42]。
3.4 TIFY蛋白的其他功能
TIFY蛋白家族还有一些其他的功能。如JAZ蛋白通过与小麦和拟南芥中的ABI5相互作用调控种子萌发[43]。罗冬兰等[14]研究发现香蕉基因MaTIFY能够随着果实的成熟而表达量增强,并可能受到乙烯诱导参与香蕉果实成熟的调控。紫云英AsJAZ1基因与豆血红蛋白AsB2510的相互作用定位于细胞膜和细胞质,参与根瘤发育及氮素固定[44]。茉莉酸甲酯(MeJA)通过诱导丹参毛状根中丹参酚酸和丹参酮的生物合成途径基因的表达,促进了丹参毛状根中丹参酚酸和丹参酮的积累,如SmJAZ8负调控丹参酚酸B的生物合成,而SmJAZ3和SmJAZ9负调控丹参酮的生物合成[45]。
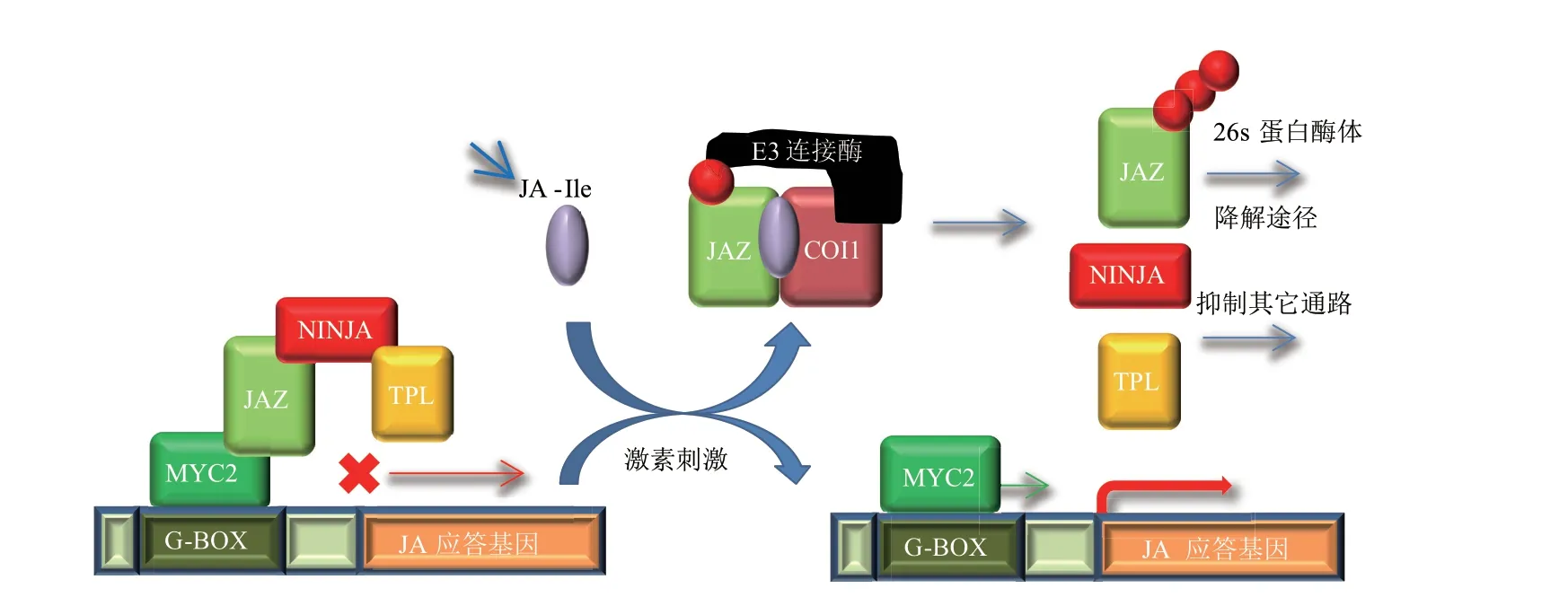
图3 JAZ蛋白介导的JA信号通路模型[35]
4 总结与展望
TIFY蛋白作为植物特有的转录因子,在植物的生长发育、响应逆境胁迫和信号转导等中发挥重要作用,其家族成员不多,目前研究最为广泛的是JAZ亚类群蛋白,因为可与MYC2互作,参与茉莉酸信号转导过程来响应生物或非生物胁迫。目前,虽然从各高等植物中分离鉴定出的TIFY蛋白很多,但已知具体功能的蛋白所占比例却还是很低。未来的研究应着力于(1)机理研究:当前只有JAZ亚家族参与JA信号通路的作用机制比较清楚,而其他亚家族如PPD、ZML可参与的通路或作用机理并不完全清楚,还需更深的认识与了解;(2)结构研究:TIFY蛋白均含有保守的TIFY结构域,各亚族包含的基本结构域也较为清楚,但除了JAZ结构域的功能研究丰富外,其他各个结构域的功能也要深入研究;(3)响应胁迫研究:近年来,TIFY蛋白在植物抗逆胁迫中的研究成为热点,多集中在植物在应对一些非生物胁迫如干旱、盐碱、渗透等逆境,而对于生物胁迫相关研究较少。而且已经发现的多为JAZ亚家族基因在逆境胁迫中的重要作用,所以挖掘其他亚家族抗逆基因将成为今后工作重点,可为作物育种提供理论依据。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
生物信息学(2020年1期)2020-05-16
文苑(2020年12期)2020-04-13
意林·全彩Color(2019年11期)2019-12-30
意林彩版(2019年11期)2019-11-22
上海农业学报(2017年3期)2017-04-10
天津科技大学学报(2016年1期)2016-02-28
戏剧之家(2016年1期)2016-02-25
红领巾·探索(2015年9期)2015-09-10
现代检验医学杂志(2015年2期)2015-02-06
