
睾丸酮丛毛单胞菌QYY降解喹啉的转录组分析
2021-01-13 07:19赵雪莹霍洪亮彭举威霍明昕
东北师大学报(自然科学版) 2020年4期
许 馨,赵雪莹,霍洪亮,于 杨,彭举威,霍明昕
(1.东北师范大学环境学院,吉林 长春 130117;2.吉林建筑大学市政与环境工程学院,松辽流域水环境教育部重点实验室,吉林 长春 130118;3.东北师范大学环境学院,吉林省城市污水处理与水质保障科技创新中心,吉林 长春 130117)
喹啉(Quinoline)是一种含氮杂环芳香性有机化合物,因其含有苯环结构使喹啉较为稳定,属于难降解有机化合物.据报道,喹啉及其部分衍生物具有毒性、致癌性和诱变性[1-2].喹啉是冶金、化学助剂、染料、杀虫剂、医药等领域的重要化工原料[3-4],同时石油石化企业和煤炭工业生产中也会产生大量含有喹啉的废水[5].由于喹啉的广泛使用和不适当的排放,导致其在水体、土壤中大量积累,危害环境和生物健康[6-7].
利用生物法去除环境中的喹啉及其衍生物是应用最为广泛的方式,由此喹啉降解菌的分离和鉴定也成为研究的热点之一.目前已被分离出具有喹啉降解特性的菌株有假单胞菌(Pseudomonassp.)、皮氏伯克霍尔德氏菌(Burkholderiapickettii)、红球菌(Rhodococcussp.)、丛毛单胞菌(Comamonassp.)等[8-13].
睾丸酮丛毛单胞菌(Comamonastestosteroni)是一种严格需氧的革兰氏阴性菌,不能利用糖类物质生长,但能够利用氨基酸、有机酸以及睾丸酮等类固醇化合物作为营养物质生长繁殖[14-16].此外,睾丸酮丛毛单胞菌还能降解芳香族化合物[17].崔明超等[5,13,18]利用分离筛选出的睾丸酮丛毛单胞菌Q10降解喹啉及其衍生物,发现这种丛毛单胞菌对喹啉有较高的降解性能;Liu等[19]利用睾丸酮丛毛单胞菌bdq06处理染料废水,通过生物强化完全去除了染料废水中高浓度的喹啉、苯酚以及其他酚类物质.
目前,利用睾丸酮丛毛单胞菌对喹啉进行生物降解的研究主要停留在降解特征和代谢途径方面,对喹啉降解相关基因表达的研究鲜有报道[20].为此,本文利用从污水处理厂活性污泥中分离出来的一株睾丸酮丛毛单胞菌(QYY)对喹啉进行了降解研究,采用转录组测序技术,从基因转录的水平上分析了喹啉生物降解时基因的差异表达情况,分析了参与降解过程的相关功能基因转录状态变化,以为进一步揭示喹啉降解基因的研究奠定基础,同时也为喹啉降解工程菌的构建提供科学指导和有力依据.
1 材料与方法
1.1 菌株
菌株睾丸酮丛毛单胞菌(QYY)由本实验室从某污水处理厂的活性污泥中分离纯化所得,现保存于中国普通微生物菌种保藏管理中心(CGMCC No.1.15223).
1.2 方法
1.2.1 喹啉降解
将QYY菌株接种到LB培养基(蛋白胨 10.0 g/L,酵母提取物5.0 g/L,NaCl 10.0 g/L)中,30℃、120 r/min条件下富集培养12 h后,10 000 r/min离心2 min,收集菌体.将菌体悬浮于磷酸盐缓冲溶液中,制成D(600)=0.5的菌悬液.取1 mL菌悬液接种到含有不同浓度(0,70,100,200,300,400 mg/L)喹啉的100 mL LB培养基中,30℃、120 r/min 条件下进行降解实验.在不同培养时间,测定喹啉剩余浓度和细菌生长情况.细菌生长的变化情况通过D(600)的数值确定,喹啉浓度变化用高效液相色谱进行测定.
1.2.2 基因组测序、组装和注释
菌株QYY基因组DNA提取后,根据Oxford Nanopore Technologies 公司提供的标准方法进行样品质量检测、文库构建、文库质量检测和文库测序等.基于Nanopore三代测序技术平台进行基因组测序,下机后的原始数据经进一步过滤接头、低质量及短片段(长度<2 000 bp)数据筛选后得到总数据集.利用Canu v1.5软件对过滤后片段进行组装后,通过Prodigal软件对组装后的基因组进行编码基因预测.
1.2.3 睾丸酮丛毛单胞菌QYY降解喹啉的转录差异分析
根据对喹啉降解实验数据的分析,选取喹啉质量浓度为100 mg/L培养6 h(Q-100组)、喹啉质量浓度为400 mg/L培养7 h(Q-400组)时的菌体,以及不含有喹啉的空白对照组培养7 h(C组)后的菌体进行转录组测序实验,每个样品3个生物学重复.收集样品进行处理,包括总RNA的提取及质量检测、cDNA文库构建、通过Illumina HiSeq测序平台对文库进行测序.
基因定量表达分析和注释:对Illmina HiSeq测序平台测得的原始数据进行过滤后,使用Bowtie2软件将过滤后的序列比对到参考基因组上.根据比对结果,采用FPKM(Fragments Per Kilo bases per Million fragments,每百万片段中来自某一基因每千碱基长度的片段数目)对表达量进行标准化,计算每个基因的表达量,一般认为FPKM值>1的基因是表达的.将比对好的基因分别与数据库进行比对注释.
差异表达基因筛选及功能富集分析:利用DESeq软件对基因表达进行差异分析,筛选出满足表达倍数差异log2|FC|>1(FC:差异变化倍数(fold change))且显著性P<0.05的基因为差异表达基因.以整个基因组为背景,将差异表达基因与GO,KEGG数据库进行序列比对(blast),得到差异表达基因的功能注释.
2 结果与分析
2.1 睾丸酮丛毛单胞菌QYY对喹啉的降解
菌株QYY对不同浓度喹啉的降解情况如图1a所示,初始质量浓度为70,100,200,300,400 mg/L的喹啉被完全降解分别需要6,8,9,11,13 h.在4 h时,菌株QYY对不同浓度喹啉的降解速率有明显变化,含有70,100,200 mg/L喹啉的培养基中菌株QYY降解速率显著较快,而质量浓度为300,400 mg/L喹啉的培养基中降解出现滞后现象.图1b表示的是不同喹啉浓度条件下菌株QYY的生长曲线,培养基中喹啉初始浓度越高,菌株QYY的生长越慢,这与图1a中菌株QYY降解喹啉的情况相符合.随着喹啉初始浓度的增加,菌株QYY的生长情况也变得迟缓,且均低于不含有喹啉的空白对照组.这是由于喹啉具有毒性,浓度越高、毒性越强,对菌株的生长抑制作用越加明显.
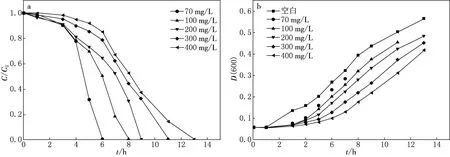
图1 睾丸酮丛毛单胞菌QYY对不同浓度喹啉的降解情况(a)和菌株在不同喹啉浓度下的生长速率(b)
2.2 睾丸酮丛毛单胞菌QYY全基因组概况
对菌株QYY进行全基因组测序,进一步了解菌株QYY基因的功能和信息.表1为睾丸酮丛毛单胞菌QYY的基因组基本信息.由表1可见,对序列进行拼接组装后得到大小约为5.67 Mb的环状染色体,GC含量为61.31%.预测到的编码基因个数为5 145个,占基因组总长度的86.93%;预测到的编码蛋白质为5 098个.非编码RNA共161个,其中rRNA有27个,tRNA有102个,其他的非编码RNA有32个.同时,在基因组中注释到8个有规律的间隔短回文重复序列.假基因、基因岛以及前噬菌体的数量分别为11,20,7个.

表1 睾丸酮丛毛单胞菌 QYY的基因组基本特征
2.3 睾丸酮丛毛单胞菌QYY降解喹啉的转录差异分析
2.3.1 转录组数量质控
基于菌株QYY全基因组结果,利用转录组测序技术研究菌株QYY降解喹啉时相关基因的表达情况:选取喹啉质量浓度为0(C)、100 mg/L(Q-100,降解6 h)和400 mg/L(Q-400,降解7 h)的菌株进行转录组测序,将测序得到的序列与参考基因组进行比对.数据质量的基本情况如表2所示.由表2可知,各样品过滤后读长均占原始读长的96%以上,Q20(碱基识别准确率在99%以上的碱基所占的百分比)、Q30(碱基识别准确率在99.9%以上的碱基所占的百分比)均能达到97%和92%,GC含量均能达到59%.这表明转录组的测序数据质量较高,可进行后续分析.
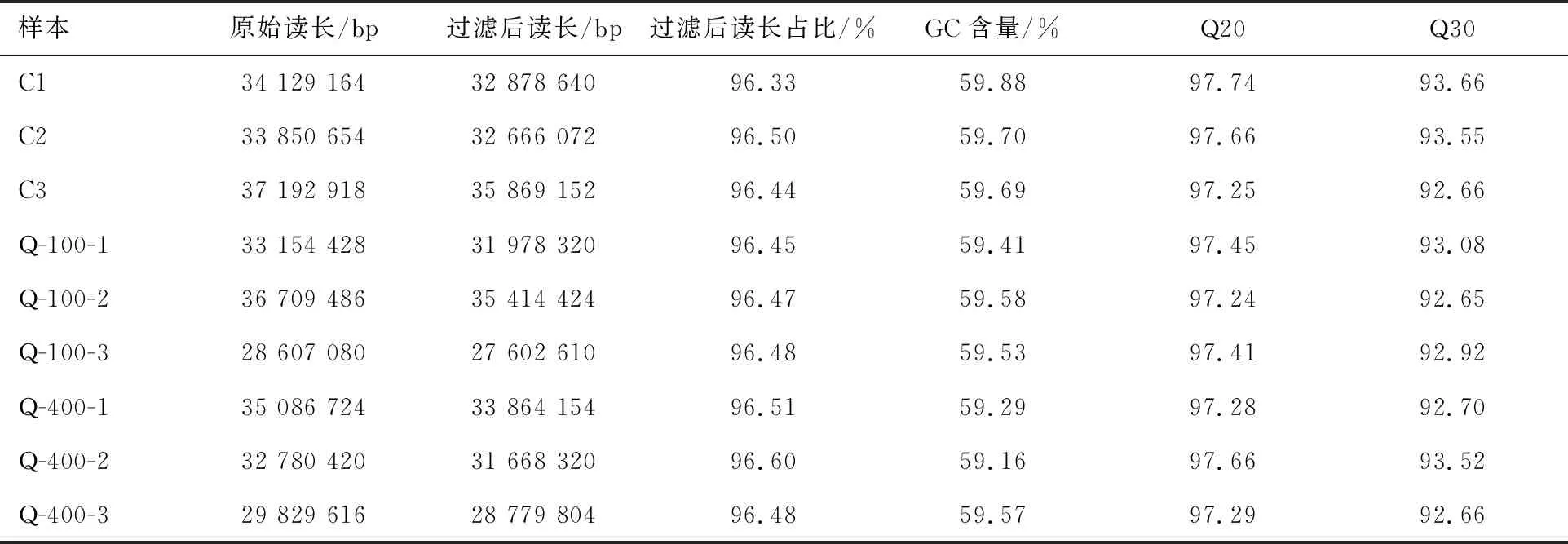
表2 数据质量统计
2.3.2 差异表达基因分析
将三组样本按照C/Q-100,C/Q-400的分组进行比对,得到1 116和1 600个差异表达的基因(log2|FC|>1,P<0.05).C/Q-100中(见图2a),上调和下调基因分别为606,510个;C/Q-400中(见图2b),上调基因为915个,下调基因为685个.以对照组C为参考基准时,Q-100和Q-400两组共有的相同差异基因的数量为951个,Q-100组中特有的差异基因数量为165个,Q-400组中特有的差异基因数量为649个.这是由于加入的喹啉浓度不同,导致菌株QYY在相同底物生长条件下差异基因有所区别.
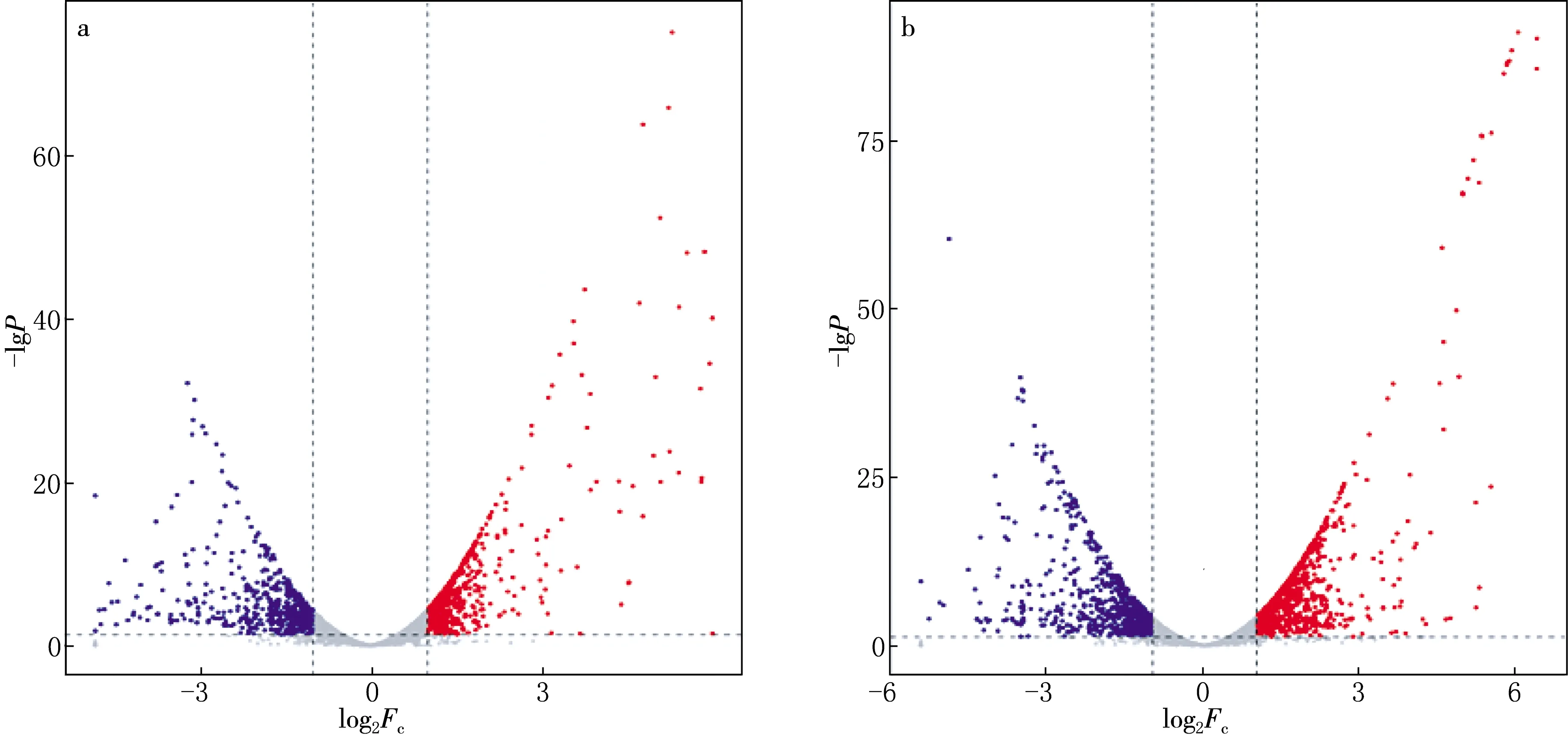
a:C/Q-100;b:C/Q-400
2.3.3 差异表达基因的KEGG富集分析
KEGG(Kyoto encyclopedia of genes and genomes)数据库整合了基因组、化学和系统功能信息,是一个综合数据库.C/Q-100和C/Q-400组中的1 116和1 600个差异表达基因分别匹配到104和110个KEGG途径中,其中匹配到代谢(metabolism)途径中的通路数量最多,分别有81和87个通路;在菌株QYY降解喹啉的生物过程中,ABC转运蛋白作为膜转运(membrane transport)中的重要部分,差异表达基因富集到其上的数目分别为61和79个.对C/Q-100和C/Q-400两组差异表达基因中关于代谢途径部分的上调表达的基因进行二级分类富集,结果见图3.编码与喹啉降解过程相关酶和ABC转运蛋白的部分差异表达基因信息见表3.其中碳水化合物代谢(carbohydrate metabolism)和氨基酸代谢(amino acid metabolism)过程均能产生能量,这些过程与能量代谢(energy metabolism)过程一同为菌株QYY降解喹啉、维持生长提供能量,转录组数据显示相关基因显著上调表达.在异生物素的生物降解与代谢(xenobiotics biodegradation and metabolism)的转录组数据中,发现与喹啉降解过程中相关的ligABC和bphB基因的差异表达.
(1) 与喹啉降解相关的差异基因.目前对喹啉降解途径和降解基因的研究不多,并且只针对假单胞菌进行了深入研究.假单胞菌中的qorMSL、oxoOR基因是降解喹啉的关键基因,但这些基因在睾丸酮丛毛单胞菌中并不存在[21-23].对转录组的结果进行分析发现,原儿茶酸代谢基因有明显上调,原儿茶酸是细菌降解多种芳香族化合物途径中最重要的中间体代谢产物,在多环芳烃的降解中发挥了重要作用.原儿茶酸通过ligAB基因编码的原儿茶酸4,5-双加氧酶(EC:1.13.11.8)转化为4-羧基-2-羟基粘康酸酯-6-半醛,再通过与ligC基因编码的2-羟基-4-羧基粘康酸酯半醛半缩醛脱氢酶(EC:1.1.1.312)、ligI基因编码的2-吡喃酮4,6-二羧酸内酯酶(EC:3.1.1.57)、ligJ基因编码的4-草酰氨基甲酸水合酶(EC:4.2.1.83)以及ligK基因编码的4-羟基-4-甲基-2-氧代戊二酸酯醛缩酶(EC:4.1.3.17)反应,最后产生丙酮酸等物质[24-27].本实验中,ligABC基因在Q-100组中较对照组基因表达量上调(见表3),但在Q-400组中该基因与对照组基因表达量相差较小,未达到差异表达的标准,但均有表达.其余参与原儿茶酸代谢酶的编码基因也均有表达.菌株QYY能够在降解喹啉的过程中产生原儿茶酸,但喹啉浓度从100 mg/L增大到400 mg/L时,会抑制菌株ligABC基因的表达.多环芳烃在双加氧酶的作用下生成二氢二醇化合物,编码二氢二醇脱氢酶(EC:1.3.1.56)的bphB基因可将二氢二醇化合物转化为2,3-二羟基联苯,进一步代谢[28].在Q-100和Q-400的转录组结果中均发现bphB基因较对照组有明显上调(见表3),表明菌株QYY的bphB基因参与喹啉降解的部分过程.

图3 Q-100和Q-400组中关于代谢途径部分表达上调的差异基因KEGG二级分类富集

表3 菌株QYY转录组中部分与喹啉降解过程有关的差异表达基因
(2) 与能量代谢相关的差异基因.为了抵抗环境的不利影响,细菌需要更多的能量维持生长,并同时抵抗环境压力[29-30].在Q-100和Q-400组中均发现属于能量代谢的氧化磷酸化过程中NADH脱氢酶(EC:7.1.1.2)、琥珀酸脱氢酶(EC:1.3.5.1)、细胞色素c氧化酶(EC:1.9.3.1)以及F型ATP酶(EC:7.1.2.2)的基因表达显著上调(见表3),这些基因的上调表达可为菌株QYY降解喹啉以及对抗喹啉毒性提供更多的能量.同时,富集到碳水化合物代谢过程中的糖酵解途径和TCA循环的关键酶基因也有明显上调.氨基酸的分解代谢可以促进能量产生,例如氨基酸代谢中丙氨酸、天冬氨酸和谷氨酸代谢可产生用于ATP合成的NADH和FADH2[31].转录组结果显示,菌株QYY中富集到丙氨酸、天冬氨酸和谷氨酸代谢等氨基酸代谢途径中的差异基因表达显著上调,这些基因的上调表达有助于维持菌株QYY的自身保护机制并对喹啉的降解代谢活动提供充分能量.
(3) 与ABC转运系统相关的差异基因.ABC转运蛋白是已知的最大和最古老的蛋白质家族之一,可以转运无机盐离子、糖、氨基酸和金属离子[32].在Q-100和Q-400组中,硫酸盐/硫代硫酸盐转运蛋白基因(cysP、cysU、cysA)显著上调(见表3).由于硫是细胞辅助因子的一种成分,包括生物素、硫胺素、辅酶A、硫辛酸、谷胱甘肽和铁硫簇,因此硫的增加能够促进细胞的生命活动[33].这与注释到的硫代谢过程的关键基因上调相符合.甜菜碱和脯氨酸是一种细胞调节渗透剂,能够在细胞受到环境胁迫时改变其在细胞内的含量,调节渗透平衡,从而保护细胞的结构[34-36].在Q-100和Q-400组中均检测到甜菜碱/脯氨酸转运蛋白的基因(proX、proW、proV)的表达上调(见表3),证明喹啉的加入使菌株在吸收和降解代谢过程中细胞渗透压发生改变,从而引起转运蛋白的增加.同时根据结果发现,两组实验条件下菌株QYY中脯氨酸代谢基因下调,这可能会造成脯氨酸在细胞内的积累,以便更好地调节细胞渗透压.支链氨基酸转运蛋白可以将包括亮氨酸在内的支链氨基酸转运到细胞中,在转录组结果中发现支链氨基酸转运蛋白的相关基因(livK、livH、livG、livM、livF)表达明显下调(见表3),不利于亮氨酸的摄取.这与结果中注释到的亮氨酸代谢基因表达下调相关.
3 结论
本研究利用全基因组和转录组测序技术,对睾丸酮丛毛单胞菌QYY降解不同浓度喹啉时的差异表达基因进行了分析,转录组结果显示:原儿茶酸作为多环芳烃降解途径中的重要中间产物,编码其降解的基因ligABC在Q-100组中表达上调明显,但在Q-400组中表达上调不明显;参与喹啉降解代谢过程中的bphB基因均上调表达.氧化磷酸化、糖酵解以及TCA循环中关键酶基因的上调保证了菌株QYY生长的能量供应.硫酸盐/硫代硫酸盐转运蛋白基因以及甜菜碱/脯氨酸转运蛋白的基因表达显著上调,以维持细胞生命活动和细胞结构.当喹啉质量浓度从100 mg/L增加到400 mg/L时,部分与喹啉降解相关的基因表达未出现差异的情况,具体原因还需要进行后续的研究.本研究从转录水平上分析了睾丸酮丛毛单胞菌QYY降解喹啉时与转运和代谢途径相关的基因的表达情况,为后续深入研究喹啉的生物代谢途径及调控提供了有力依据和方向.
猜你喜欢
中国防痨杂志(2022年7期)2022-11-25
军事文摘(2022年16期)2022-08-24
当代水产(2022年3期)2022-04-26
纺织检测与标准(2021年1期)2021-12-05
绵阳师范学院学报(2021年11期)2021-11-24
今日农业(2021年11期)2021-08-13
感染、炎症、修复(2021年1期)2021-07-28
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
科技与创新(2020年16期)2020-08-18
