
长链非编码RNA MALAT1通过PI3K/Akt信号传导通路调控肝细胞癌生物学行为的研究
2021-04-23 02:52尹家俊
中国实验诊断学 2021年4期
吴 惧,李 贺,岳 坤,程 楠,徐 键,尹 敏,尹家俊,余 杰
(大连大学附属中山医院 肝胆胰腹腔镜外科病房,辽宁 大连116001)
肝细胞癌(HCC)是世界上最常见的恶性肿瘤之一,是与癌症相关死亡的第二大病因[1]。尽管目前在肝癌的诊断和治疗方面已取得一定的进展,但该疾病导致的高死亡率以及不良预后仍然是困扰着医学界的一大难题[2]。长链非编码RNA(lncRNA)是一种长度超过200个核苷酸(nt)的非编码RNA[3]。近年来,越来越多的证据表明,lncRNA参与了多种细胞过程的调控,包括细胞的生长和增殖,细胞凋亡和代谢[4-5]。转移相关的肺腺癌转录本1(MALAT1),是一个进化高度保守的lncRNA基因,由8 000 nt组成,位于染色体11q13[6]。最近研究发现MALAT1可调节多种癌症中的细胞迁移,增殖,侵袭和凋亡,例如乳腺癌[7],大肠癌[8]和胃癌[9]等。本研究旨在探讨MALAT1在肝癌细胞中的表达,及其对肝癌细胞增殖,迁移以及体外成瘤的影响,并探讨其发生机制。
1 材料与方法
1.1 材料
肝癌细胞系HepG2、Huh-7及正常肝细胞L-O2均由中国科学院(上海)细胞类型培养研究所购得。Akt,PI3K一抗购自美国abcam公司;MEM和DMEM培养基购自美国HyClone公司;MALAT1 siRNA 干扰序列购自GenePharma(上海吉玛公司);转染试剂GP-transfect-Mate购自GenePharma(上海吉玛公司);引物由大连宝生物科技有限公司设计以及合成;RT-PCR仪购自美国ABI公司;蛋白电泳及转膜设备购自美国Bio-Rad公司。
1.2 细胞培养转染及分组
L-O2和Huh-7细胞均培养于含10%胎牛血清、1%双抗的DMEM培养基,置于37℃、5%CO2、饱和湿度的培养箱中,0.25%胰蛋白酶消化,按1∶2-1∶3 的比例传代。HepG2细胞培养于含10%胎牛血清、1%双抗的MEM培养基,同样置于37℃、5%CO2、饱和湿度的培养箱中,0.25%胰蛋白酶消化,按1∶2-1∶3的比例传代。实验所用细胞均为对数生长期细胞。取肝癌细胞系培养后随机分为三组:肝细胞癌组(Blank 组),阴性转染组(si-NC组),MALAT1siRNA组(si-MALAT1组)。将对数生长期的HepG2和Huh-7细胞以转染前一天2×105接种到6孔板中,培养过夜,当细胞密度达到60%-80%通过转染试剂GP-transfect-Mate ,按照说明书方法分别转染,4-6小时后查看转染效率(图1)。48 h后用于提取RNA,72 h后用于提取蛋白。

图1 Huh-7细胞转染效率 (A:转染前 B:转染后)

1.4 CCK-8法检测细胞增殖转染后24 h收集转染细胞,按密度为1×103/孔接种于96孔板中。细胞在37℃孵育0 h、24 h、48 h和72 h,在指定的时间点每孔加入10 μL CCK-8试剂,在37°C孵育2 h后,用酶标仪在450 nm波长下测光密度值(OD值),整理统计数据,综合分析数据,得出结果。
1.5 集落形成试验检测肝癌细胞体外成瘤转染后24 h收集转染细胞,机械悬浮成单个细胞悬液。然后,将转染细胞计数并接种到6孔板中,平均每孔2000个细胞。在37℃、5%CO2的潮湿气氛中孵育2周后,用磷酸盐缓冲液(PBS)洗涤3遍,无水甲醇固定20 min,结晶紫染色15 min,用PBS洗涤3遍,然后空气中干燥。在光学显微镜下拍摄染色细胞并计数,每孔随机选择5个视野。
1.6 划痕愈合检测肝癌细胞迁移HCC细胞以5×106细胞/孔的密度接种在六孔板。长满后,用枪头沿直尺垂直划痕,每孔3条划痕,再用PBS清洗3遍,去除划下的细胞,加入无血清培养基, 放入37℃ 5%CO2培养箱,培养。按0 h,24 h,48 h取样,拍照。
1.7 Western blot检测PI3K、Akt蛋白表达量提取转染72 h后的细胞总蛋白,SDS-PAGE电泳分离,湿法转膜,用含5%脱脂奶粉的TBST室温封闭1 h,用TBST清洗2遍,加一抗(PI3K1∶1 000,Akt1∶1 000,Beta-actin1∶500),4℃孵育过夜后用TBST洗涤3遍,加二抗(1∶10 000),室温孵育1 h,用TBST清洗3遍,用 ECL 法化学发光,用 ImageJ分析扫描灰度值,Beta-actin设置为内参照,分析蛋白表达水平。
1.8 数据统计分析计量资料采用均值±标准差(Mean±SD)表示,并用SPSS20.0软件进行统计学分析。两组间数据比较采用t检验,三组间比较选用方差分析,以P<0.05为差异具有统计学意义。
2 结果
2.1 lncRNA MALAT1 在肝癌细胞中的表达结果
运用PCR技术检测正常肝细胞与肝癌细胞中的MALAT1表达情况,结果显示,与正常肝细胞(L-O2)相比,肝癌细胞(HepG2和Huh-7)中MALAT1的表达水平显著增高,差异具有统计学意义(P<0.05),图2A;转染后,肝癌细胞中lncRNA MALAT1的表达水平显著下降,差异具有统计学意义(P<0.05),图2B。
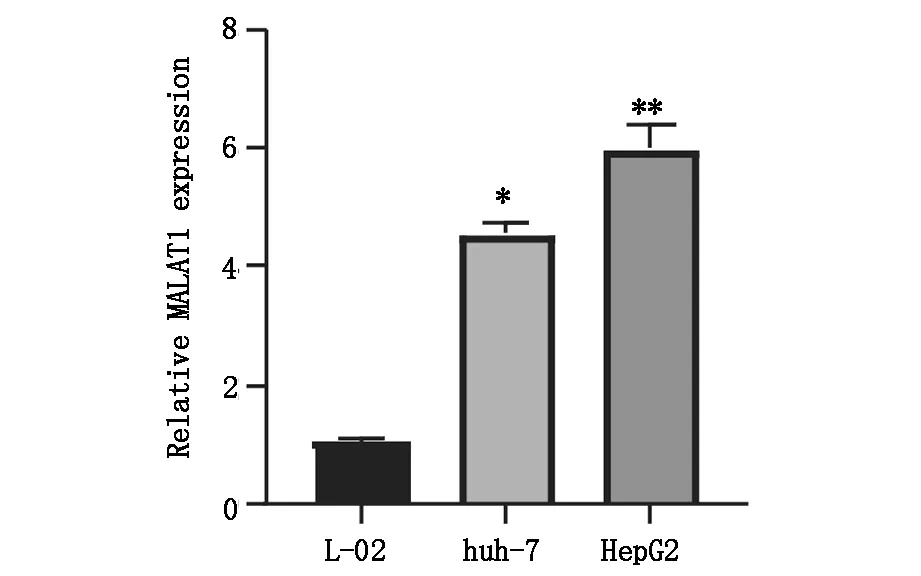
图2A lncRNA MALAT1在肝癌细胞系及正常肝细胞系中的表达,*P<0.05,**P<0.05.
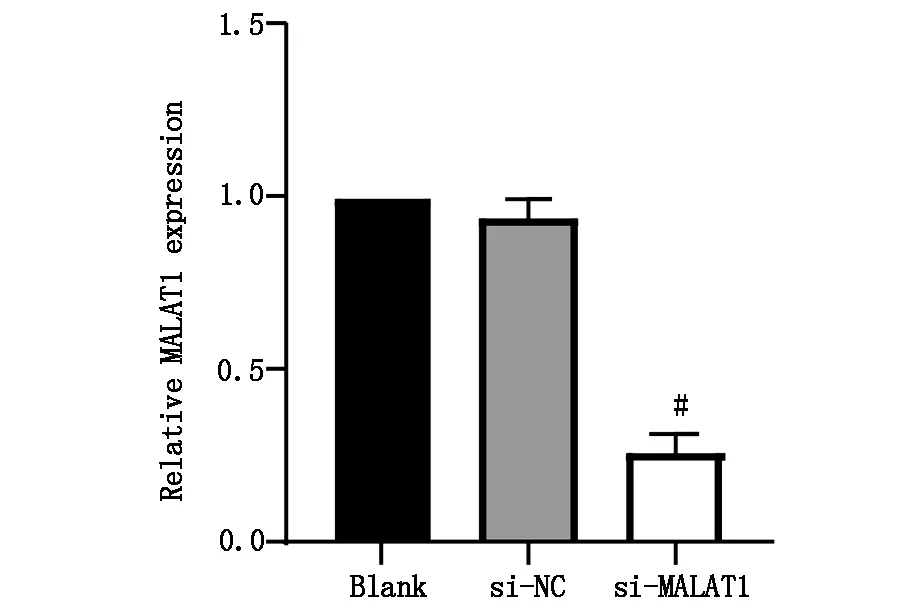
图2B siRNA转染后肝癌细胞lncRNA MALAT1的表达,#P<0.05
2.2 下调lncRNA MALAT1表达对肝癌细胞增殖的影响
CCK-8检测三组肝癌细胞增殖情况,与Blank 组(5.212±0.132)相比,si-NC组细胞增殖率(4.741±0.211)无明显差异(t=2.976,P>0.05),si-MALAT1组细胞增殖率(2.785±0.174)明显降低,差异具有统计学意义(t=4.016,P<0.05),图3A,图3B。
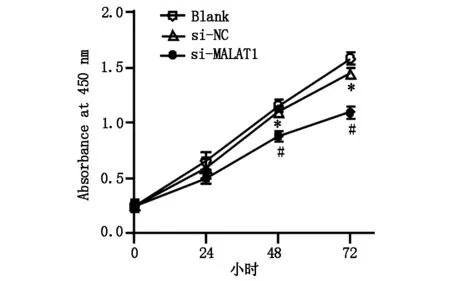
图3A Blank 组、si-NC组、si-MALAT1组细胞增殖曲线,
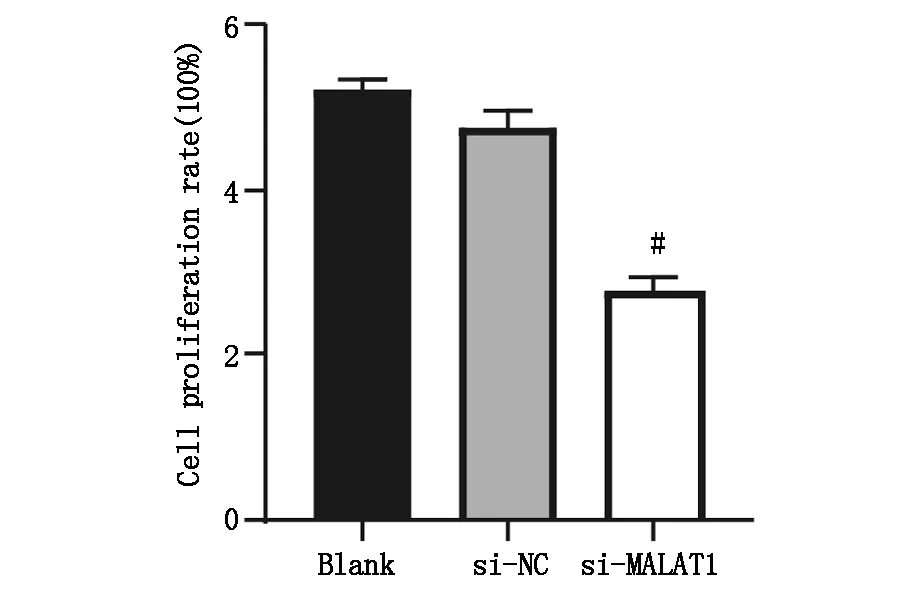
图3B Blank 组、si-NC组、si-MALAT1组细胞增殖率,
2.3 下调lncRNA MALAT1表达对肝癌细胞迁移的影响
划痕愈合实验结果表明,与Blank 组(63.477±4.324)%相比,si-NC组细胞迁移愈合率(58.108±5.341)%无明显差异(t=1.897P>0.05),si-MALAT1组细胞迁移愈合率(31.420±6.235)%明显降低,差异具有统计学意义(t=5.316P<0.05),图4A,图4B。
2.4 下调lncRNA MALAT1表达对肝癌细胞体外成瘤能力的影响
集落形成实验结果表明,与Blank 组细胞集落形成率(52.141±5.320)%相比,si-NC组细胞集落形成率(46.880±4.571)%无明显差异(t=1.774,P>0.05),si-MALAT1组细胞集落形成率(22.753±6.382)%明显降低,差异具有统计学意义(t=5.962,P<0.05),图5A,图5B。

图4A Huh-7 细胞在200倍视野下0 h、48 h的迁移愈合情况

图4B Blank 组、si-NC组、si-MALAT1组细胞迁移愈合率,#P<0.05

图5A Huh-7细胞集落形成实验
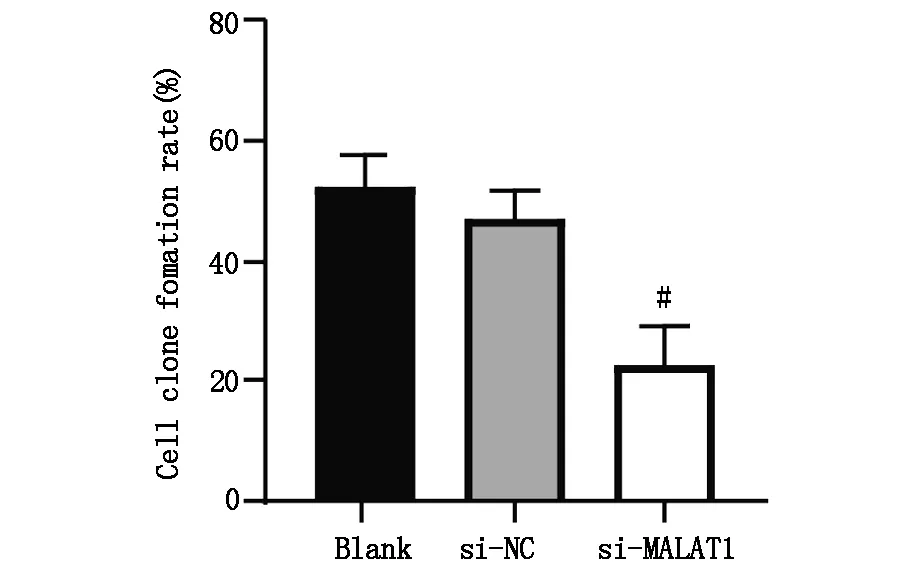
图5B Blank 组、si-NC组、si-MALAT1组细胞集落形成率,#P<0.05
2.5 下调lncRNA MALAT1对肝癌细胞PI3K、Akt蛋白表达的影响
蛋白免疫印迹实验检测三组细胞PI3K、Akt蛋白表达情况,与Blank组相比,si-NC组各蛋白表达无明显差异(P>0.05),si-MALAT1组肝癌细胞内PI3K、Akt蛋白表达明显降低,差异具有统计学意义(P<0.05),图6A,图6B1,B2。
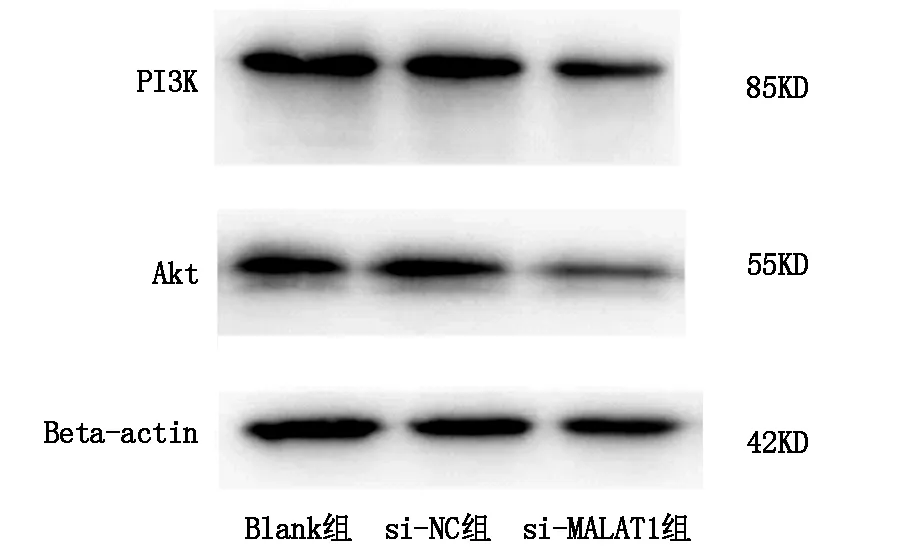
图6A 蛋白免疫印迹检测转染siRNA后对细胞PI3K、Akt表达的影响

图6B1 Blank 组、si-NC组、si-MALAT1组细胞PI3K蛋白表达,#P<0.05
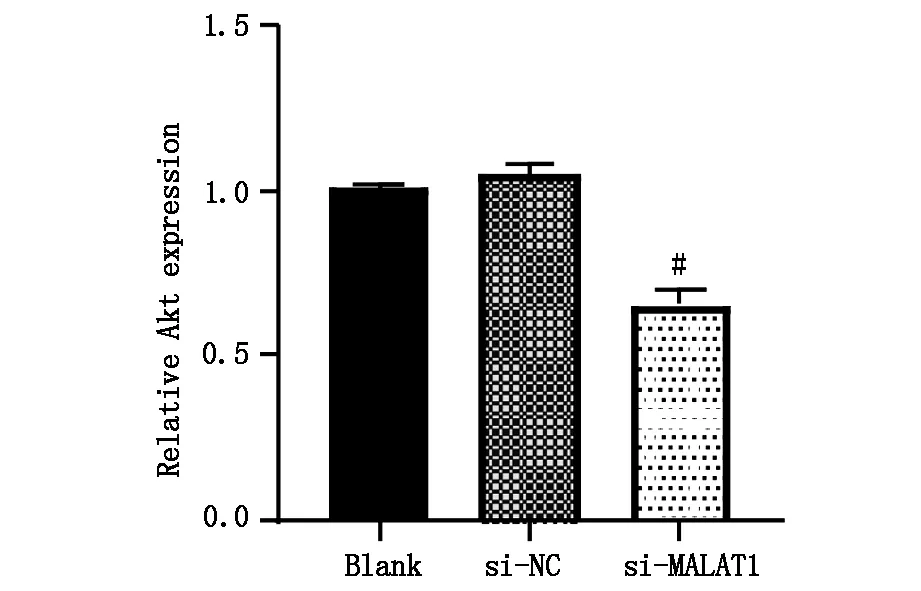
图6B2 Blank 组、si-NC组、si-MALAT1组细胞Akt蛋白表达,#P<0.05
3 讨论
肝癌在临床上是极其常见的恶性肿瘤[1]。据统计,我国肝癌的病死率在肿瘤中排第二位[10]。许多危险因素,包括酒精性肝病,非酒精性脂肪性肝病,肝硬化,HBV和HCV感染,均会影响肝癌发生与发展[11-12]。然而,肝癌的潜在发病机制目前尚未明确。尽管目前国内外在肝癌的诊断和治疗方面有了很大进展,但是肝癌患者的治疗效果以及远期预后仍不理想,其5年生存率仅为15%[13]。肝癌患者的不良预后主要归因于术后肿瘤复发和转移的发生率高,且缺乏有效的辅助治疗[14]。近年来大量文献表明,lncRNA在肝癌的发生发展以及预后中起到重要作用[15-16]。
长链非编码RNA(lncRNAs)是一种长度超过200nt、缺乏编码蛋白质能力的RNA序列,起初被认为是基因组转录的“噪音”,不具有生物学功能[17]。近年来研究表明,lncRNAs参与了多种细胞过程的调控,包括细胞的生长和增殖,细胞凋亡和代谢[4-5]。其在肿瘤细胞生长、代谢过程中作用日渐受到重视。MALAT1是一个进化高度保守的lncRNA基因,由8000 nt组成,编码基因定位于染色体11q13.1[6]。最早是在研究非小细胞肺癌中被发现[18]。Schmidt 等[19]对非小细胞肺癌的研究发现,MALAT1 具有促进肺癌形成的能力,且高表达的MALAT1与肺癌的预后不良有关。它在胃癌、乳腺癌、结肠癌等多种肿瘤组织中均有不同程度的表达[7-9]。本研究结果显示:与正常肝细胞(L-O2)相比,肝癌细胞中MALAT1表达升高;si-MALAT组的肝癌细胞的增殖、迁移率均低于空白对照组(Blank 组),阴性对照组(si-NC组)。提示MALAT1在肝癌中高表达且与肝癌的发生发展密切相关。
PI3K/Akt信号通路是重要的凋亡抑制信号转导通路,其在胃癌、结直肠癌、宫颈癌、鼻咽癌等多种肿瘤生长代谢中发挥重要作用[20]。PI3K 活化后可激活下游蛋白Akt,活化后的Akt可调节其下游多种凋亡相关基因,促进凋亡基因失活,进而抑制细胞凋亡。Zhang SH等人[21]研究证实,LncRNA-MALAT1介导PI3K/Akt 信号通路调控高糖诱导的内皮细胞的增殖、凋亡、迁移和血管生成。Jin Y 等人[22]研究表明,LncRNA-MALAT1介导PI3K/Akt 信号通路促进卵巢癌上皮细胞增殖、代谢。本研究进一步对MALAT1与PI3K、Akt之间的相互作用机制进行探究,通过抑制MALAT1的表达,siRNA 组细胞 PI3K、Akt 蛋白表达量明显下降;siRNA 阴性对照组无明显变化。实验结果表明:下调 MALAT-1可以抑制 PI3K/Akt 信号通路上关键靶点PI3K与Akt的表达,进而抑制肝癌细胞增殖、迁移和体外成瘤能力。
本研究仍然存在一些不足之处,如: MALAT1是如何对PI3K/Akt 信号通路上关键靶点进行影响,MALAT1是否会改善肝癌患者的预后,都值得下一阶段进一步探究。
综上所述,MALAT1 在肝癌中高表达,且MALAT1能够促进肝癌细胞增殖、迁移以及体外成瘤能力。下调肝癌细胞中 MALAT1后,可抑制其生物学行为。其机制可能与其影响PI3K/Akt信号传导通路上关键靶点的表达有关,为临床上治疗肝癌提供了新的思路。
猜你喜欢
传染病信息(2022年6期)2023-01-12
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年5期)2021-10-12
石河子大学学报(自然科学版)(2018年6期)2018-04-12
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
郑州大学学报(医学版)(2016年4期)2016-08-11
癌症进展(2016年10期)2016-03-20
癌变·畸变·突变(2016年3期)2016-02-27
华南农业大学学报(2015年5期)2015-12-04
