
外源GABA对西伯利亚白刺幼苗盐胁迫伤害缓解效应
2021-06-29 01:00郑曌钰肖雅楠
东北农业大学学报 2021年5期
刘 威,王 贺,王 骁,赵 野,郑曌钰,肖雅楠
(东北农业大学园艺园林学院,哈尔滨 150030)
土壤盐分是全球农业主要非生物胁迫[1],盐胁迫导致渗透胁迫和离子毒害,影响植物正常生长发育。盐胁迫对植物的危害存在于各方面,包括对种子萌发[2]、光合作用[3]、离子积累及抗氧化系统[4]等方面。植物为应对盐胁迫已形成许多复杂生理机制,如调控体内盐分分配、渗透调节、活性氧清除等。除此之外,激素调节、多胺及植物体信号物质也参与抵御盐胁迫过程。
γ-氨基丁酸(GABA)是一种在植物体内天然存在的非蛋白质氨基酸,更是一种重要的胞内信号分子,参与植物体内与胁迫相关基因表达调控[5]。GABA在植物体内由谷氨酸脱酸酶(GAD)催化L-谷氨酸脱羧和多胺降解两条途径生成;GABA转氨酶(GABA-T)是代谢关键酶,促使其分解代谢进入三羧酸循环(TCA)。正常情况下,植物组织中GABA含量较低,在植物受到逆境胁迫时,刺激GAD活性提升,诱导GABA迅速积累,防止细胞酸中毒,从而对植物起到保护作用。此外,在GABA分解过程中可通过调节胞质pH、诱导激素合成(如乙烯等)和渗透调节等方式提高植物适应性,增强植物抗逆性[6]。外源施用GABA显著增强植物对各种非生物胁迫耐受性,据报道,外源GA⁃BA应用通过促进与次级代谢相关渗透调节物质积累提高匍匐剪股颖抗旱能力[7],也通过改善光合作用保护玉米幼苗免受盐分伤害[8]。
西伯利亚白刺(Nitraria sibiricaPall.)为蒺藜科白刺属(NitrariaL.)落叶具刺灌木,因其具有较强耐盐能力,成为改良盐碱化土壤主要造林树种。当前国内外对西伯利亚白刺响应盐胁迫研究主要集中于盐胁迫下西伯利亚白刺种子萌发[9]、光合特性[4]、离子平衡[10]及转录组学、代谢组学等方面。盐分对白刺的影响与浓度相关,研究表明,低浓度NaCl(200 mmol·L-1)处理使西伯利亚白刺幼苗Na+含量显著增加并降低根中K+/Na+[10];中度NaCl胁迫(300 mmol·L-1)影响唐古特白刺光合作用能力[11];高盐度NaC(l400 mmol·L-1)抑制白刺生长[9]。此外,盐胁迫还影响白刺氮代谢关键酶活性,抑制谷氨酰胺合成,从而降低白刺氮代谢速率,对白刺生长发育造成伤害[11]。
然而,鲜有报道外源GABA对盐胁迫下西伯利亚白刺幼苗生长特性及GABA支路的影响。本试验通过研究200 mmol·L-1NaCl胁迫下,施加不同浓度外源GABA对西伯利亚白刺幼苗生理特性和GABA代谢支路中代谢物和关键酶活性的影响,探究外源GABA提高白刺耐盐性作用机理,为外源应用GABA减轻植物盐胁迫伤害和提高植物抗盐性提供理论依据。
1 材料与方法
1.1 试验材料
供试西伯利亚白刺种子由吉林白城市林业科学院提供。挑选籽粒饱满,大小均一种子,用体积分数10%NaClO溶液消毒5 min,流水和蒸馏水分别冲洗5次后,置于光照培养箱中。待其长至4~5片真叶时定植于花盆中,挑选长势一致一年生实生苗,移植于纯净河沙花盆中,1/2 Hoagland全营养液浇灌培养。缓苗后对幼苗进行盐处理,试验处理如表1所示,每个处理3次重复,共18盆。每天同一时间喷施5、10 mmol·L-1GABA溶液,以全部叶片湿润,但无过量GABA溶液滴下为标准,7 d后取样并测定相关指标。

表1 试验处理Table 1 Experimental treatments
1.2 试验设计
试验于东北农业大学园艺实验站开展。试验设置盐分(NaCl分析纯,0、200 mmol·L-1),GABA(Sigma,0、5、10 mmol·L-1)两因素随机组合,设置6个处理(见表1),3次重复。
1.3 测定指标及方法
测量不同处理下西伯利亚白刺幼苗鲜重和干重,通过计算得出根茎比和叶片肉质化程度。公式如下:根茎比(%)=(地下根部分干重/地上茎部分干重)×100;叶片肉质化程度(%)=(叶鲜重/叶干重)×100;测定不同处理下西伯利亚白刺株高;参照李合生方法测定相对含水量、叶片叶绿素含量、电解质外渗率、丙二醛、脯氨酸和可溶性糖含量[12];考马斯亮蓝法测定可溶性蛋白含量;参照Zhang和Bown方法测定内源GABA含量[13];参照Bartyzel等方法测定GAD活性[14];利用ELISA试剂盒测定GABA-T活性。
1.4 数据处理与分析
GraphPad Prism 7.0软件处理数据并绘图,运用SPSS25.0软件分析数据差异显著性。
2 结果与分析
2.1 外源GABA对盐胁迫下西伯利亚白刺幼苗生理特性的影响
如表2所示,外源喷施不同浓度GABA对西伯利亚白刺幼苗株高和根茎比有抑制作用,随GABA浓度升高,株高和根茎比呈降低趋势。盐胁迫(T3)下西伯利亚白刺幼苗株高和根茎比与对照差异显著,对西伯利亚白刺株高产生抑制作用,株高下降23%。但对西伯利亚白刺幼苗根茎比有显著促进作用,因西伯利亚白刺为一种盐生植物,200 mmol·L-1NaCl导致地下部分生长活跃。在NaCl胁迫下施用5 mmol·L-1GABA(T4)显著增加西伯利亚白刺株高,其高度较NaCl处理(T3)增加17.9%,且5 mmol·L-(1T4)与10 mmol·L-1(T5)GABA对盐胁迫下西伯利亚白刺株高降低的缓解作用无显著差异。但T4和T5处理显著降低根茎比,随外源GABA浓度升高对盐胁迫下根茎比的影响表现为先降后升。说明盐胁迫下适宜浓度外源GABA促进西伯利亚白刺幼苗生长且对茎的促进作用大于根。

表2 NaCl胁迫下不同浓度GABA对西伯利亚白刺幼苗生理特性的影响Table2 Effectsof different concentrationsof GABA on physiological characteristicsof N.siberia seedlingsunder NaCl stress
外源喷施不同浓度GABA溶液,西伯利亚白刺幼苗叶片相对含水量呈先增后降趋势,最大值时GABA浓度为5 mmol·L-(1T1),而随GABA浓度升高,叶绿素含量下降,与对照组差异显著。盐胁迫(T3)导致西伯利亚白刺幼苗相对含水量和叶绿素含量显著下降(P<0.05),分别比对照降低6.15%和32.9%。盐胁迫下喷施不同浓度外源GABA,叶片相对含水量呈增加趋势,与盐胁迫(T3)差异显著,说明外源GABA对盐胁迫西伯利亚白刺幼苗渗透胁迫有缓解作用。不同浓度外源GABA对西伯利亚白刺幼苗在盐浓度为200 mmol·L-1下叶绿素合成具有一定促进作用,当GABA浓度为5 mmol·L-1(T4)时,对西伯利亚白刺幼苗中叶绿素含量增加效果最明显,相比于T3增加39.2%,差异显著(P<0.05)。虽然T5高于T4但差异不明显,可以看出,随GABA浓度增加叶绿素含量在不断积累但变化趋于稳定。
正常栽培条件下施加5、10 mmol·L-1GABA对丙二醛含量和电解质外渗率影响不明显;NaCl处理下(T3)西伯利亚白刺叶片中丙二醛含量显著升高,较对照增加约0.3倍;施加不同浓度GABA后MDA含量显著降低,分别降低50.47%和23.37%,西伯利亚白刺叶片中MDA含量最低时GABA浓度为5 mmol·L-(1T4)。盐胁迫(T3)下西伯利亚白刺幼苗电解质外渗率增加,是对照3.1倍。外源施用GABA降低盐胁迫下西伯利亚白刺幼苗电解质外渗率,当GABA浓度为5 mmol·L-(1T4)时,电解质外渗率最低。说明适宜浓度外源GABA(5 mmol·L-1)可缓解盐胁迫对植物细胞膜伤害,减少离子流动,降低叶片电解质外渗率和丙二醛含量。
2.2 外源GABA对盐胁迫下西伯利亚白刺幼苗渗透调节物质含量的影响
从图1可看出,无NaCl胁迫下,脯氨酸含量随GABA浓度增加呈先升后降趋势,并在GABA浓度为5 mmol·L-1时达到最大值。NaCl处理下脯氨酸含量较对照增加0.3倍,差异显著。喷施GABA后脯氨酸含量随GABA浓度升高呈先降后略微上升趋势,分别较NaCl处理下降11.06%和4.40%,当GABA浓度为5 mmol·L-1时脯氨酸含量最低。说明盐胁迫条件下西伯利亚白刺积累脯氨酸参与渗透调节,防止细胞内水分过度流失,且外源GABA在一定程度上缓解NaCl胁迫下西伯利亚白刺叶片脯氨酸含量增加。
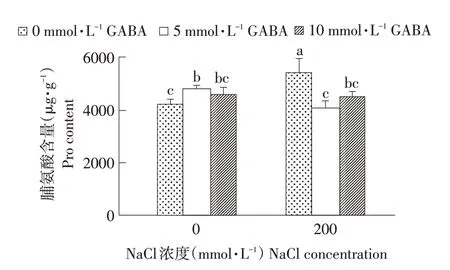
图1 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺幼苗脯氨酸含量的影响Fig.1 Effect of different concentrations of exogenous GABA on prolinecontents in N.siberia seedlingsunder NaCl stress
如图2所示,正常条件下,随GABA浓度升高,西伯利亚白刺叶片可溶性糖含量逐渐减少,当浓度升至10 mmol·L-1时,叶片中可溶性糖含量较对照降低34.2%。然而,盐胁迫使可溶性糖含量大量积累,约为对照1.1倍,差异显著。外源GA⁃BA使盐胁迫下西伯利亚白刺可溶性糖含量大幅下降,且随GABA浓度升高表现出先降后升趋势,当GABA浓度为5 mmol·L-1时西伯利亚白刺体内可溶性糖含量最低。说明在西伯利亚白刺遭受盐胁迫时,加速体内以淀粉形式存在糖的水解作用,增加体内可溶性糖含量,而一定浓度外源GABA缓解西伯利亚白刺叶片淀粉水解作用,降低可溶性糖含量。
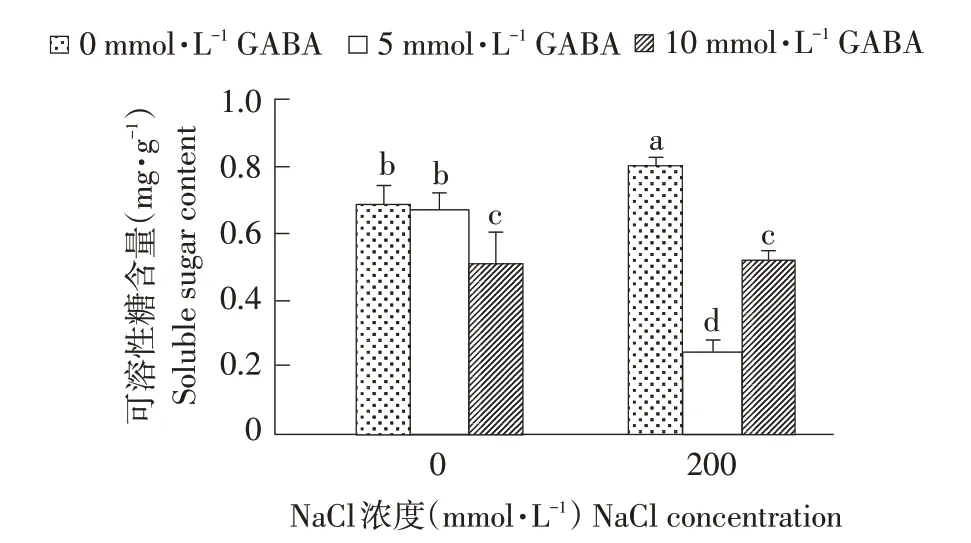
图2 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺幼苗可溶性糖含量的影响Fig.2 Effect of different concentrationsof exogenous GABA on the solublesugar contents in N.siberia seedlings under NaCl stress
NaCl及外源GABA对西伯利亚白刺叶片可溶性蛋白含量影响如图3所示,正常栽培条件下外源GABA提高可溶性蛋白含量,当GABA浓度为5 mmol·L-1时,可溶性蛋白含量达峰值,与对照差异显著(P<0.05);NaCl胁迫降低西伯利亚白刺叶片可溶性蛋白含量,但外源GABA处理使可溶性蛋白含量先略微上升而后显著下降,10 mmol·L-1GA⁃BA处理时可溶性蛋白含量降到最低。表明在盐胁迫条件下,西伯利亚白刺渗透能力减弱,可溶性蛋白含量降低,且适宜浓度外源GABA对盐胁迫下西伯利亚白刺可溶性蛋白含量增加具有一定作用。
2.3 外源GABA对盐胁迫下西伯利亚白刺幼苗GABA支路的影响
由图4可看出,无论正常栽培还是盐胁迫下,适宜浓度外源GABA处理可增加西伯利亚白刺幼苗叶片内源GABA含量。非NaCl胁迫条件下,外源施用10 mmol·L-1GABA提高西伯利亚白刺幼苗叶片内源GABA含量,是对照1.03倍;NaCl处理导致内源性GABA大量积累,而外源GABA降低盐胁迫下西伯利亚白刺幼苗体内GABA含量。随外源GABA浓度增加,内源性GABA含量表现为先降后升趋势,当GABA浓度为5 mmol·L-1时,显著降低盐胁迫下内源GABA含量,且与对照无明显差异。表明适宜浓度外源GABA可增加正常栽培条件下西伯利亚白刺幼苗内源GABA含量,同时在盐胁迫下,适宜浓度外源GABA有效促进内源GABA合成与积累,为GABA支路提供丰富前体物质,降低盐胁迫对西伯利亚白刺伤害。
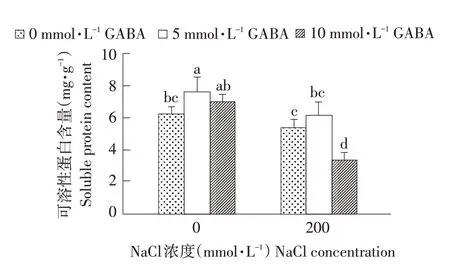
图3 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺幼苗可溶性蛋白含量的影响Fig.3 Effect of different concentrations of exogenous GABA on thesolubleprotein contentsin N.siberia seedlings under NaCl stress
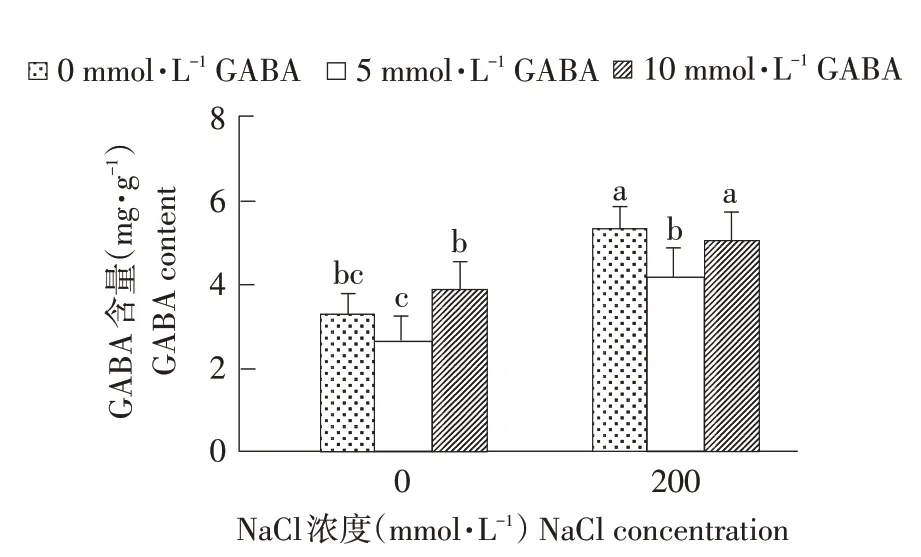
图4 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺幼苗内源GABA含量的影响Fig.4 Effect of different concentrations of exogenous GABA on theendogenous GABA contentsin N.siberia seedlings under NaCl stress
GAD活性变化影响西伯利亚白刺体内GABA含量。由图5可知,正常条件下,外源施加GABA对西伯利亚白刺幼苗叶片内GAD活性无明显影响,200 mmol·L-1NaCl处理下GAD活性达到最大值且显著高于对照(P<0.05);盐胁迫下施加外源5、10 mmol·L-1GABA,GAD活性分别低于单独NaCl胁迫处理组1.1%和0.8%。由此表明,盐胁迫使GAD活性升高,促进L-Glu脱羧生成GABA,促进GABA积累,且外源喷施GABA降低GAD活性。

图5 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺GAD活性的影响Fig.5 Effects of different concentrations of exogenous GABA on GAD activitiesin N.siberia under NaCl stress
GABA-T是GABA分解关键酶,在逆境胁迫中发挥重要作用。由图6可知,外源施用GABA增强GABA-T活性,西伯利亚白刺幼苗内GABA-T活性与GABA浓度呈正相关,当GABA浓度为10mmol·L-1时达到最大值,显著高于对照,是对照1.2倍;NaCl处理降低GABA-T活性,但影响不显著,随GABA浓度增加,GABA-T活性先升后降。当GABA浓度为10 mmol·L-1时,分别低于同组盐处理和5 mmol·L-1GABA处理14.3%和16.7%,与其他处理组差异显著(P<0.05)。由此表明,正常情况下施加外源GA⁃BA提高西伯利亚白刺幼苗叶片内GABA-T活性,但外源GABA降低盐胁迫下GABA-T活性,从而促进内源GABA积累。
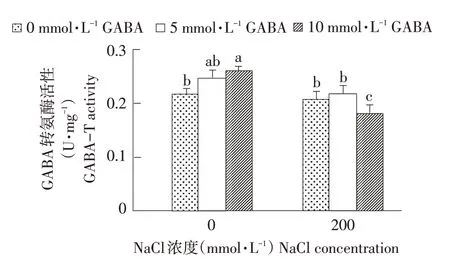
图6 NaCl胁迫下不同浓度外源GABA对西伯利亚白刺GABA-T活性的影响Fig.6 Effectsof different concentrations of exogenous GABA on GABA-T activitiesin N.siberia under NaCl stress
3 讨论与结论
土壤盐分对植物生长有不利影响,植株高度和根茎比是重要生长指标。本研究中盐胁迫对西伯利亚白刺影响表现在株高显著变矮而根茎比显著升高。虽然西伯利亚白刺耐盐性较强,但其地上茎部、地下根部受盐胁迫影响程度不同。这与低浓度盐对黑果枸杞幼苗根生长具有促进作用,对茎生长影响不明显结果一致[15]。植物通过多种生理生化作用相互影响应对盐胁迫,叶片相对含水量是植物体内各种生理指标综合体现。本研究中,盐处理降低西伯利亚白刺幼苗叶片相对含水量,与盐胁迫下菊花幼苗相对含水量显著下降结论一致[16],说明盐处理使土壤水势降低,导致植物吸水困难。
当植物缺水时,细胞将积累脯氨酸、可溶性糖和可溶性蛋白,增强细胞内渗透调节能力,提高细胞保水性并维持水分平衡。本研究中,脯氨酸和可溶性糖含量在盐胁迫下显著增加,可溶性蛋白降低,与酸枣中3种渗透调节物质含量均增加研究结果不一致[17]。可能原因是西伯利亚白刺遭受盐胁迫时,叶片细胞受离子毒害,促使蛋白质分解,使可溶性蛋白含量下降。盐胁迫下植物体内产生大量活性氧自由基,MDA含量高低和电解质外渗率可衡量膜损伤程度。本研究中两者变化趋势与观音竹研究结果一致[18]。植物内源GABA含量变化趋势与GAD和GABA-T活性有关,本研究中盐胁迫下西伯利亚白刺幼苗叶片内源GABA积累是因GAD活性增加,GABA-T活性无显著变化,与Zarei等发现GABA高积累是由于GAD激活和GABA-T抑制引起的研究结果一致[19]。
本研究中,NaCl胁迫下西伯利亚白刺幼苗表现出典型盐分破坏症状,包括株高、相对含水量、叶绿素含量显著降低,丙二醛、电解质外渗率显著升高。外源GABA处理后,5 mmol·L-1GA⁃BA对恢复盐胁迫下西伯利亚白刺叶片相对含水量效果最佳,施加不同浓度外源GABA均增加叶绿素水平以减轻盐胁迫造成损害。上述结果表明,通过添加外源GABA,可减弱西伯利亚白刺幼苗生长受到的抑制,与王馨对西伯利亚白刺研究一致[20]。盐胁迫引起渗透和氧化胁迫,导致细胞膜透性增加,GABA被认为是脂质过氧化过程中MDA形成抑制剂[21]。当前的研究中,盐胁迫下外源GABA处理后MDA和电解质外渗率均显著下降,且5 mmol·L-1缓解效果最佳,证明叶面喷施GABA溶液提高西伯利亚白刺抵抗盐胁迫能力。本研究中盐胁迫下可溶性糖含量大量积累,这与李焕勇等对盐胁迫下小果白刺研究结果一致[22],添加GABA溶液可溶性糖呈下降趋势,说明一定浓度外源GABA可缓解西伯利亚白刺叶片淀粉水解作用,降低可溶性糖含量。盐胁迫导致植物细胞中蛋白质合成代谢增强,提高渗透调节能力以促进植物适应盐胁迫环境。本研究中,盐处理下5 mmol·L-1GABA溶液促进可溶性蛋白增加,提高细胞保水能力,保护细胞膜免受损伤,但过量浓度GABA溶液抑制可溶性蛋白合成。此外,本研究结果显示,脯氨酸在西伯利亚白刺体内积累以适应盐环境,可能是由于盐胁迫影响脯氨酸合成代谢酶活性,致使脯氨酸含量升高以维持细胞内渗透平衡。GABA添加降低脯氨酸含量,这与利用GABA使甜瓜抵抗Ca(NO3)2胁迫结果一致[23]。
近年来,综合研究GABA支路生理和遗传学表明,其功能对于植物响应非生物胁迫(例如极端温度和盐分)正常生长所必需。本研究结果表明,Na⁃Cl处理下西伯利亚白刺叶片内源GABA含量显著增加,外源GABA处理虽未进一步增加但也显著高于正常条件下GABA含量,与盐碱胁迫下施加GABA对甜瓜品种“金辉”胚芽中GABA含量研究结果一致[24]。GAD和GABA-T是GABA合成和分解关键酶,在植物对逆境的防御反应中发挥关键作用。本研究中,盐胁迫显著增加GAD活性但对GABA-T活性无显著影响。外源添加不同浓度GABA后GAD活性均降低,而GABA-T呈先升后降趋势,其中GA⁃BA-T降低较显著,与干旱胁迫下白三叶GAD和GABA-T活性均增加,但第10天加入外源GABA后,两种酶活性均降低结果一致[25]。在盐胁迫时添加GABA后西伯利亚白刺体内GABA含量仍较高,可能是由于较低GABA降解速率,并非与GAD活性增加有关,这与Rujira研究结果一致[26]。
综上所述,NaCl胁迫对西伯利亚白刺幼苗株高、根茎比、相对含水量、叶绿素含量、MDA含量、电解质外渗率、渗透调节物质含量和GABA代谢关键物质含量和关键酶活性产生影响,且外源喷施GABA后,通过对渗透调节以及GABA支路的影响产生缓解NaCl胁迫的效果。上述研究结果为改善盐碱地和培育盐生植物提供理论基础。
猜你喜欢
中老年保健(2022年3期)2022-11-21
舰船科学技术(2022年11期)2022-07-15
草业科学(2022年3期)2022-03-26
安徽化工(2021年3期)2021-05-29
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
红领巾·成长(2019年3期)2019-04-16
中学生物学(2008年6期)2008-08-29
