
等边浅蛤线粒体全基因组测序和系统发育研究
2021-08-05 12:33夏立萍郭宝英叶莹莹
浙江海洋大学学报(自然科学版) 2021年2期
夏立萍,徐 鸣,郭宝英,叶莹莹
(国家海洋设施养殖工程技术研究中心,浙江舟山 316022)
线粒体基因组(mitochondrial genome)是一种序列长度相对较短,基因信息丰富且易于分离的DNA 分子。具有母系遗传、进化速率快、包含大量同源基因等特点,且广泛存在于真核生物细胞中,因而成为基因组测序的优先目标被广泛应用于系统发育研究中[1]。绝大多数动物线粒体基因组是共价闭合的双链DNA分子,分子量较小,一般介于14~19 kb 之间,基于G+T%碱基含量来区分DNA 双链重轻链。无脊椎动物线粒体基因组通常包括13 个蛋白质编码基因,22 个tRNA 基因和2 个rRNA 基因以及控制基因复制和转录的控制区[2-3]。少数种类在基因组成上存在变化:如太平洋牡蛎Crassostrea gigas 线粒体基因组中存在3 个rRNA 基因(2 个12S rRNA 基因);绣球海葵Metridium senile 的线粒体基因组中只有2 个tRNA 基因[4];双壳纲贝类一些物种线粒体基因组缺失了ATP8 基因,如文蛤Meretrix meretrix,短文蛤M.petechialis 等[5]。
等边浅蛤Macridiscus multifarious(Kong,Matuskuma &Lutaenko,2012)隶属于软体动物门Mollusca,双壳纲Bivalvia,帘蛤目Veneroida,帘蛤科Veneridae,浅蛤属Macridiscus。等边浅蛤分布于日本、朝鲜、越南、印度尼西亚和印度等地,在我国北起辽宁,南至海南均有分布,常栖息在潮间带中潮区、低潮区至浅海的砂质海底,尤以低潮区最为常见[6]。壳长在1~5 cm 之间,壳高在8 mm~4 cm 之间[7]。因其味道鲜美及丰富的营养而成为中国海岸线养殖中重要的双壳类动物之一。目前浅蛤属中有3 个物种,分别是等边浅蛤、黑浅蛤M.melanaegis (Römer,1860)、半布目浅蛤M.donacinus (Koch,1843)。浅蛤属物种的分类地位在历史上经历了多次变革,包括学名也有较大的争议。Macridiscus 最初在1902 年由DALL 提出,并将其归为花蛤属Gomphina 的亚属,直至MIKKELSEN,et al[8]基于形态学与线粒体16S rRNA 基因、线粒体COI 基因、28S rRNA 核基因、以及H3 核基因4 种DNA 分子标记进行了系统发育研究,首次将这一类群提升为浅蛤属。2012 年KONG Lingfeng,et al[9]基于形态学及线粒体Cytb 基因及核基因ITS 区2 种分子标记发现了除黑浅蛤与半布目浅蛤之外的隐种等边浅蛤。虽然等边浅蛤是一种常见的物种,但相关的遗传学研究较少,线粒体基因组研究尚属空白。过往研究只有对该物种形态,群体组成,繁殖周期,生态习性以及染色体核型等方面的研究[10-14],研究补全了等边浅蛤线粒体全基因组和基于线粒体全基因组的系统发育树,为等边浅蛤的种别分析和保护研究提供了研究参考。
1 材料与方法
1.1 样品采集和DNA 提取
本研究于2014 年1 月在舟山市南沙沙滩(122.2°E,30.3°N)进行等边浅蛤样品的采集,取等边浅蛤闭壳肌组织保存于无水乙醇中,在-20 ℃冰箱保存。盐析法提取DNA[15],1%琼脂糖凝胶电泳检测DNA 质量及完整性。
1.2 线粒体基因组测序
委托上海元莘生物有限公司使用二代测序方法[16]基于Illumina 高通量测序平台,对等边浅蛤DNA 样品进行高通量测序。样品基因组DNA 检测合格后,用超声波将DNA 打断,然后对打断后DNA 进行纯化构建测序文库,步骤依次为:-DNA 末端修复,-3’端加A,连接测序头,琼脂糖凝胶电泳后回收目的片段,连接测序头琼脂糖凝胶电泳法回收目的片段,对目的片段进行PCR 扩增,最终建成测序文库。测序前对建好的文库进行质检,合格后采用Illumina HiSeqTM平台测序。
1.3 序列拼接与分析
利用MITObim (https://github.com/chrishah/MITObim)拼接软件对测序读段序列进行多次迭代拼接,得到最优的组装结果。基于denovo 拼接的线粒体基因组序列,利用MITOS 工具(http://mitos2.bioinf.unileipzig.de/index.py)进行基因组注释。tRNA scan-SE 软件(http://lowelab.ucsc.edu/tRNAscan-SE/)进行定位tRNA。利用在线软件CHLOROBOX (https://chlorobox.mpimp-golm.mpg.de/OGDraw.html)绘制线粒体基因组环状图。使用软件BioEdit 统计序列长度、碱基组成、GC 含量并计算AT 偏移度。
从GenBank 中下载帘蛤科15 个物种的线粒体基因组序列,缢蛏作为外群,选择包括等边浅蛤在内的17 个物种的全部蛋白质编码基因[17-22],使用MEGA7.0 将序列进行多重比对,并利用邻接法构建neighborjoining (NJ)系统进化树。
2 结果与分析
2.1 线粒体基因组结构与组成
等边浅蛤线粒体基因组全长20 248 bp,GenBank 登录号为MH932410,由13 个蛋白编码基因,22 个tRNA 基因和2 个rRNA 基因组成,全部在重链编码。13 个蛋白质编码基因包括:基因ATP8、NAD4、COX1、Cytb、ATP6、NAD3、NAD5、NAD6、COX3、NAD1、NAD2、NAD4L 和COX2。其中D-loop 控制区位于COX1 基因和Cytb 基因之间,长度为1 944 bp。在13 个蛋白质编码基因中,ATP8、COX1、NAD6 和NAD2的起始密码子是ATT,NAD4、NAD5 和NAD1 起始密码子是ATA,Cytb、ATP6、NAD3、COX3、NAD4L 和COX2 起始密码子是ATG。对于终止密码子,NAD3、NAD5、COX3 和NAD4L 为TAG,其余8 个蛋白质编码基因都是以TAA 终止(图1、表1)。
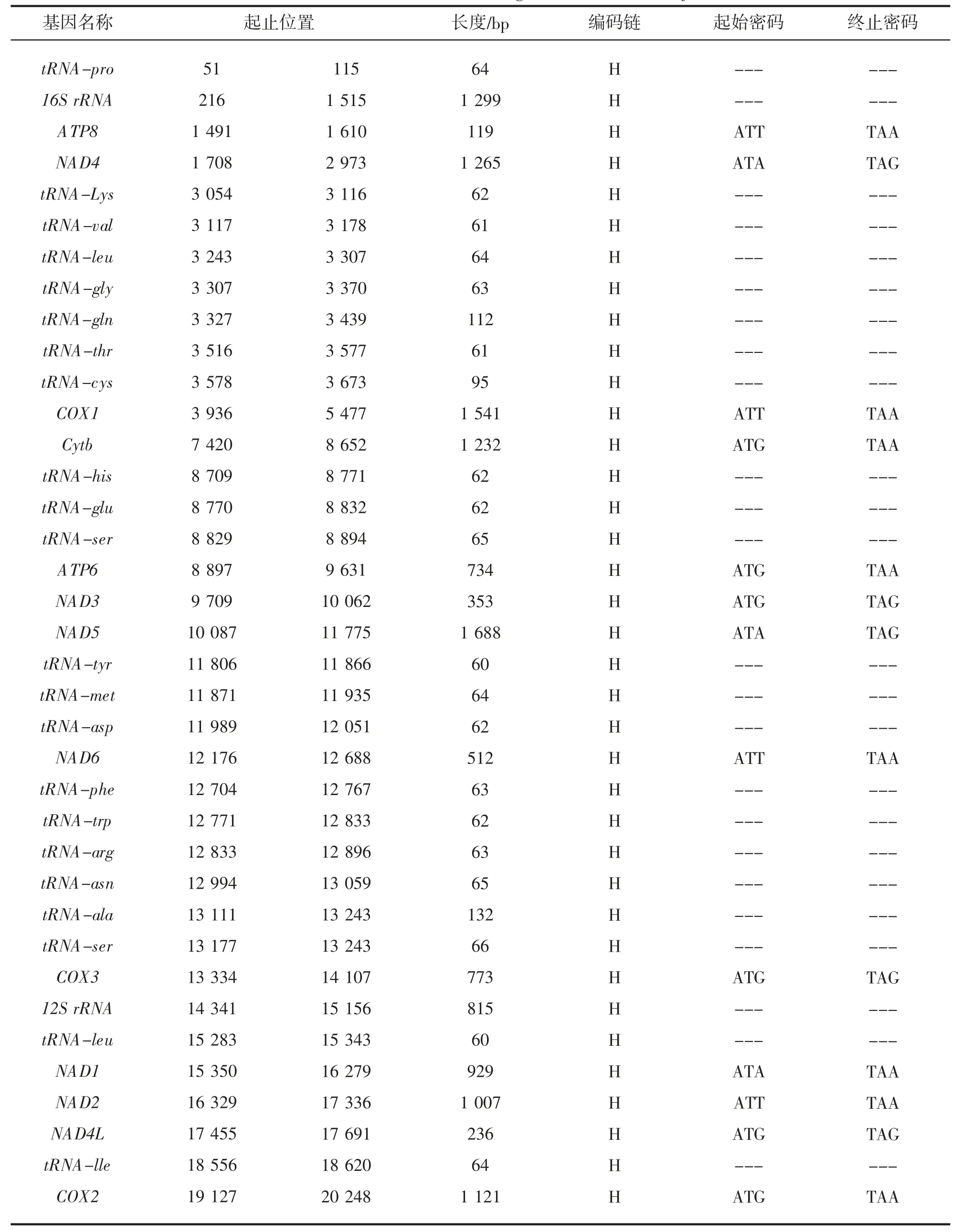
表1 等边浅蛤线粒体基因组结构特点Tab.1 Characteristics of the mitogenomes of M.multifarius
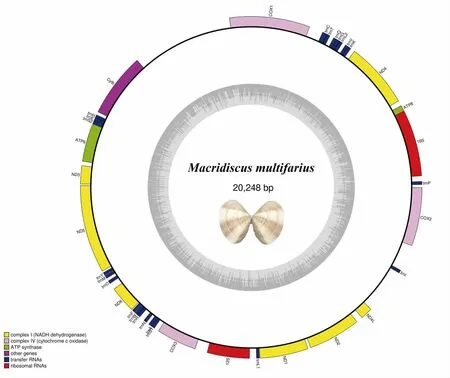
图1 等边浅蛤线粒体基因组环状图Fig.1 The mitochondrial genome mapping of M.multifarius
2.2 核苷酸组成
由表2 可以看出,帘蛤科贝类基因组A+T(63.26%~70.48%)含量均高于G+C(26.91%~36.74%)含量,核苷酸具有明显的AT 偏向性。等边浅蛤线粒体全基因组碱基含量由高到低依次是T(39.33%)>A(28.55%)>G(21.72%)>C(10.4%),A+T 含量居中为67.88%,A+T skew 碱基组成和G+C skew 碱基组成分别为-0.159 和0.352。
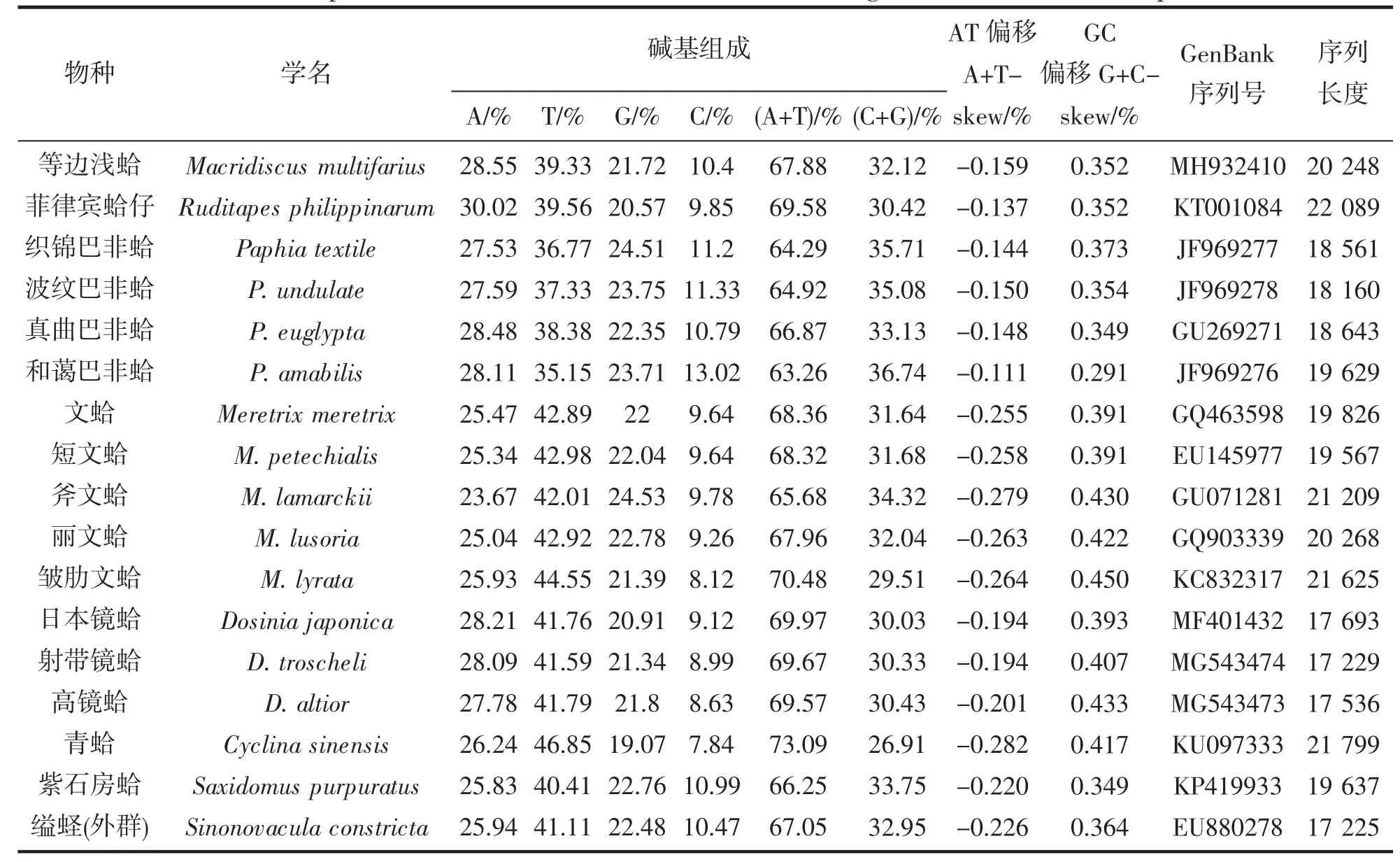
表2 帘蛤科物种线粒体基因组碱基组成Tab.2 Composition and skewness in the mitochondrial genomes of Veneridae species
2.3 蛋白质编码基因
等边浅蛤线粒体基因组具有13 个蛋白质编码基因,其碱基组成见表3。13 条蛋白质编码基因的A+T含量均高于50%,可见其在蛋白编码基因组中也具有AT 偏好性。
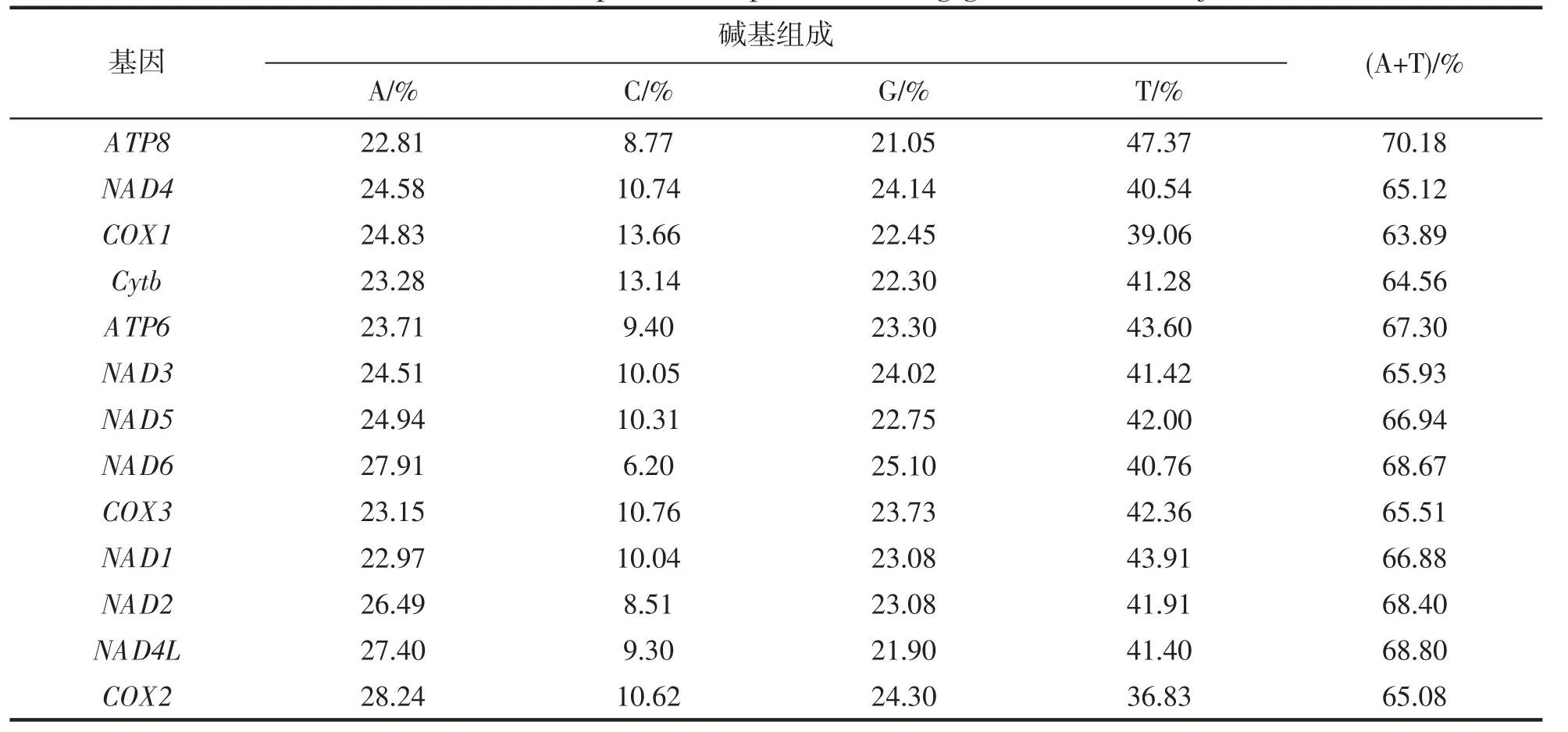
表3 等边浅蛤蛋白编码基因碱基组成Tab.3 Nucleotide composition of protein-coding genes in M.multifarius
2.4 等边浅蛤线粒体基因组进化分析
由于文蛤和短文蛤缺少ATP8 基因,我们将等边浅蛤与其他16 种贝类的线粒体基因组的12 个(ATP8除外)进行多重序列比对和分析,构建NJ 系统发育树,以研究浅蛤属的分类地位及进化关系。等边浅蛤与其他15 种贝类均属帘蛤科Veneridae,等边浅蛤属于浅蛤属Macridiscus,菲律宾蛤仔Ruditapes philippinarum 属于蛤仔属Ruditapes;织锦巴非蛤Paphia textile、波纹巴非蛤P.undulate、真曲巴非蛤P.euglypta、和蔼巴非蛤P.amabilis 属于巴非蛤属Paratapes;文蛤Meretrix meretrix、短文蛤M.petechialis、斧文蛤M.lamarckii、丽文蛤M.lusoria、皱肋文蛤M.lyrata 属于文蛤属Meretrix;日本镜蛤Dosinia japonica、射带镜蛤D.troscheli、高镜蛤D.altior 属于镜蛤属Dosinia;青蛤Cyclina sinensis 属于青蛤属Cyclina;紫石房蛤Saxidomus purpuratus 属于石房蛤属Saxidomus;缢蛏Sinonovacula constricta 则属于竹蛏科Solenoidea,缢蛏属Sinonovacula 作为外群。物种登录号及序列长度等详细信息见表2。
由图2 可见,等边浅蛤与菲律宾蛤仔聚为1 个小支,与巴非蛤属的4 个物种织锦巴非蛤、波纹巴非蛤、真曲巴非蛤和蔼巴非蛤聚为1 个支,它们又与镜蛤属3 个物种日本镜蛤、射带镜蛤、高镜蛤聚为一大支;而文蛤属5 个物种文蛤、短文蛤、斧文蛤、丽文蛤、皱肋文蛤与紫石房蛤、青蛤聚为另一大支;缢蛏在两大分支外独立作为一支。等边浅蛤与菲律宾蛤仔亲缘关系最近。

图2 基于线粒体基因组12 个蛋白质编码基因构建的NJ 系统进化树Fig.2 The neighbor-joining phylogenetic tree based on mitochondrial genome of 12 protein-coding genes
3 讨论
3.1 等边浅蛤线粒体基因组具有AT 偏好性
等边浅蛤与其他15 种帘蛤科贝类的AT 含量均高于50%。其中等边浅蛤A+T 含量为67.88%,12 种蛋白质编码基因的A+T 含量也全部高于50%,具有明显的AT 偏好性。AT 偏移值和GC 偏移值分别为-0.159 和0.352。已有的帘蛤科贝类报道中,同样发现具有较高的AT 含量[23]。且查阅文献发现不只是帘蛤科生物,在所有动植物,甚至细菌、真菌都具有AT 碱基偏好性特性[24-32]。这或许与生物的进化有关,但关于AT 碱基偏好性的产生机制暂无明确的研究结果,还有待继续研究。
3.2 帘蛤科系统进化关系
系统进化树显示(图2),作为外群的缢蛏在进化树上单独作为一支,其余部分主要由两大分支构成,巴非蛤属4 个物种、等边浅蛤、菲律宾蛤仔及镜蛤属2 个物种聚为一大支,青蛤、紫石房蛤及文蛤属5 个物种聚为另一大支,BPs 值均接近100%。其中等边浅蛤与菲律宾蛤仔分在同一细支,亲缘关系最近,两物种的CDS 区全部在重链编码,这可能是等边浅蛤与菲律宾蛤仔亲缘关系更近的原因之一。在帘蛤科16 个物种中,文蛤及短文蛤缺少ATP8 基因,其余14 个物种包括等边浅蛤具有全部的13 个编码蛋白基因。该结果与程汉良等[33]利用线粒体16S rRNA 基因序列研究帘蛤科贝类分子系统发育以及韩冷[34]利用核糖体18S rDNA、ITS 以及28S rDNA 基因序列研究了5 种帘蛤科贝类的亲缘关系均基本一致,结果均显示蛤仔属与镜蛤属亲缘关系最近,其次是青蛤属,文蛤属与石房蛤属亲缘关系最近聚为另外一支。但以上这2 个研究并未涉及到浅蛤属的样品。本研究补充了等边浅蛤线粒体基因组信息,为解决帘蛤科传统分类的一些难点问题提供新的思路和理论依据,同时在帘蛤科贝类系统重建中具有重要意义。
猜你喜欢
临床肺科杂志(2022年3期)2022-11-26
中华实用诊断与治疗杂志(2022年1期)2022-08-31
军事文摘(2022年16期)2022-08-24
今日农业(2022年4期)2022-06-01
中国卒中杂志(2021年7期)2021-11-29
今日农业(2021年14期)2021-10-14
今日农业(2021年11期)2021-08-13
初中生世界·七年级(2019年10期)2019-11-25
食品与健康(2019年7期)2019-07-18
中国体育科技(2018年6期)2018-12-13
