
甜菜铵转运蛋白BvAMT1-3基因的克隆及生物信息学分析
2021-10-17 01:18魏多刘铭曦高卓李佳佳刘大丽马龙彪
中国糖料 2021年4期
魏多,刘铭曦,高卓,李佳佳,刘大丽,马龙彪
(1.黑龙江省普通高等学校甜菜遗传育种重点实验室/黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2.中国农业科学院北方糖料作物资源与利用重点实验室/中国农业科学院甜菜研究所,哈尔滨 150080;3.哈尔滨师范大学生命科学与技术学院,哈尔滨 150025;4.黑龙江大学生命科学学院,哈尔滨 150080)
0 引言
氮素(N)是植物所需的一种重要营养物质,在植物生长发育中起关键作用。氮是植物体内许多重要有机化合物的组分,也是遗传物质的基础[1]。氮不仅是蛋白质、核酸、核蛋白、叶绿素和许多酶的组成成分,同时也参与合成一些维生素,生物碱和植物激素中也都含有氮[2-4]。植物吸收的氮素以铵态氮()和硝态氮()的形式为主,由于植物对两种氮素的吸收和代谢水平不同,会使吸收不同氮素的植物在生长发育过程中产生较为明显的差异[5]。植株体内氮素的同化产物和营养水平也会对铵转运蛋白(AMT)的活性产生影响[6]。研究表明,同化铵态氮需要的能量低于硝态氮,因此植物缺氮时会优先吸收铵态氮,而植物主要通过调控AMT 的合成来调控对氮的吸收[7-8]。农作物根部铵转运因子的过度表达,对农作物的生长有积极的影响,例如在微型番茄根部使AMT过表达,一方面可以提高农作物的质量,另一方面也能提高作物抵抗低氮胁迫的能力[9]。如今大多数铵转运蛋白主要属于AMT1和AMT2两个基因家族[10]。
目前已经从拟南芥、水稻、番茄、百脉根、油菜、山杨、苜蓿、绿藻、小麦、大豆、高粱等作物中都发现了铵转运蛋白的存在[11]。研究表明,拟南芥中含有6 个AMT基因,水稻里最少有12 个AMT基因[12]。AMT基因家族的表达活性受不同氮素形态和铵浓度的影响,当NH4+浓度过高时会抑制植物叶片和根系的生长[13]。同时,缺氮也会对植物AMT基因的表达造成一定的影响。在低氮胁迫3 d 后,拟南芥中AtAMT1-1和AtAMT1-3的表达呈现上升趋势[14-15]。
氮肥对作物的生长、产量和品质起着重要作用,增加氮肥的施用量是提高产量的一个重要途径[16-17],但增施氮肥的同时也增加了环境污染[16,18]。因此,为了对铵转运蛋白进行进一步的研究,提高氮素利用率,本试验利用RT-PCR方法克隆了甜菜BvAMT1-3基因,并采用生物信息学相关技术对该基因进行了较为全面的分析,以期为培育对氮素吸收和利用效率高的甜菜品种及为BvAMT基因后续功能的研究奠定一定的基础。
1 材料与方法
1.1 试验材料
甜菜‘780016B/12 优’,从国家甜菜种质资源中期库取得。准备4 片真叶期的甜菜叶片组织来提取总RNA以供后续的试验。
1.2 试验方法
1.2.1 RNA的提取
采用Trizol(Invitrogen)法,从1 g 甜菜的叶片组织中提取得到总RNA。利用紫外分光光度计NavoVue plus在OD260和OD280下测定RNA 浓度及纯度。1.0%Agarose electrophoresis进一步检测RNA的质量。
1.2.2 反转录cDNA及PCR扩增
取3µg总RNA,利用First strand cDNA synthesis kit(TOYOBO)将其反转录为cDNA。根据甜菜Beta vulgaris ammonium transporter 1-3(BvAMT1-3,LOC104899971)基因的碱基序列分别设计引物如下:
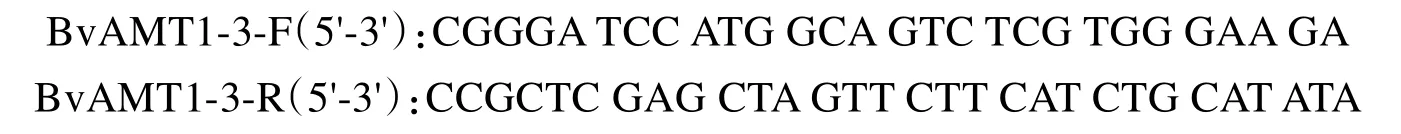
以cDNA 为模板,利用KOD-Plus-Neo高保真酶(TOYOBO)PCR扩增该基因CDS全长。
1.2.3 克隆载体的构建及目的基因鉴定
取2µLPCR 产物,于25 ℃连接到载体pEASY-Blunt 上,把连接好的产物转化入T1 感受态细胞,在含有IPTG 和X-gal的平板培养基进行Amp抗性筛选,选择阳性重组子质粒DNA,利用BamHI和XhoI进行限制性内切酶酶切和PCR 鉴定。把所获得的电泳结果无误差的样品送至上海生工生物工程公司测序。将所得到的质粒DNA 溶液置于-20 ℃保存用于进一步试验。
1.2.4 生物信息学分析
利用NCBI 的ORF finder(https://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析BvAMT1-3的开放阅读框和CDS,运用在线软件pfam(http://pfam.xfam.org/)预测该基因所编码蛋白的保守结构域。Expasy 网站的Prot-Param(https://web.expasy.org/)在线计算氨基酸理化性质,各项指标包括:氨基酸数目、分子量、理论等电点和不稳定指数。用NCBI 中的在线软件BLAST(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行同源性分析。利用SignalP5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)进行信号肽预测。通过TMHMM2.0 在线软件(http://www.cbs.dtu.dk/services/TMHMM/)预测跨膜结构域。利用ProtScale(https://web.expasy.org/protscale/)进行亲疏水性的预测,通过PSORT(https://www.genscript.com/psort.html)预测蛋白的亚细胞定位。
1.2.5 甜菜BvAMT1-3蛋白系统发生分析
将候选甜菜AMT 家族蛋白序列与拟南芥、菠菜、黄花蒿、番茄和水稻AMT 家族序列用ClustalX 和MEGA6.0软件进行多序列比对和Neighbor-Joining进化树的构建。
2 结果与分析
2.1 甜菜BvAMT1-3基因的克隆
根据Beta vulgaris ammonium transporter 1-3(BvAMT1-3,LOC104899971)基因的碱基序列,设计该基因CDS 两端的特异性引物,并在起始密码子和终止密码子两端分别引入BamHI 和XhoI 酶切位点。利用特异性引物来进行PCR扩增。在1.39 kb左右的位置获得了BvAMT1-3目的条带(图1)。将此PCR产物与Marker显示的标准条带对比发现位置无误,可进行下一步试验。
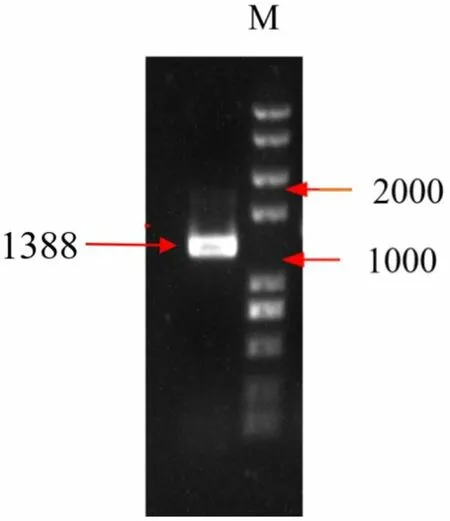
图1 BvAMT1-3 PCR扩增电泳Fig.1 PCR amplification of BvAMT1-3 gene
将BvAMT1-3基因与克隆载体进行连接,转化到大肠杆菌T1中,转化完成后,采用蓝白斑筛选与抗性筛选得到阳性克隆,并对所得的阳性克隆进行质粒提取,双酶切后电泳检测,表明在1%Agarose 凝胶的3.93 kb和1.39 kb 的位置上,获得了相对应于载体pEASY-Blunt 和目的基因片段BvAMT1-3的条带(图2)。通过与Marker标准条带对比发现位置无误差,可判断连接正确。
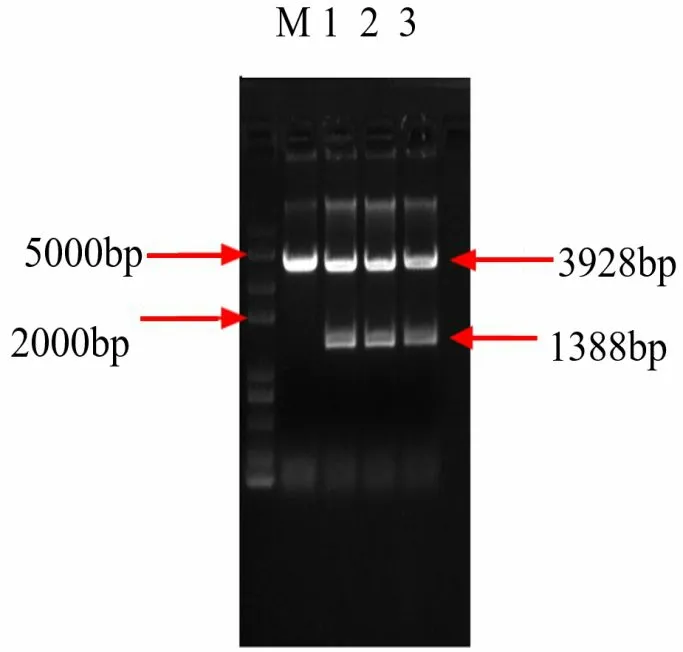
图2 pEASY-Blunt-BvAMT1-3重组质粒的鉴定Fig.2 Identification of recombinant plasmid of pEASYBlunt-BvAMT1-3
2.2 甜菜BvAMT1-3基因生物信息学分析
2.2.1 甜菜BvAMT1-3基因的序列分析
甜菜BvAMT1-3基因的CDS 全长为1 388 bp,编码462 个氨基酸(图3)。根据ProtParam 分析计算,该基因所编码蛋白的分子量为49.729 kDa,等电点(Theoretical pI)为5.30。不稳定系数分析表明,蛋白的不稳定指数(Instability index)小于40,属于稳定蛋白。根据SignalP-5.0 分析该蛋白没有信号肽,不属于分泌蛋白。利用在线软件SOPMA 和POSRT 对甜菜BvAMT1-3 二级结构和亚细胞定位进行分析,结果表明BvAMT1-3蛋白定位于内质网膜结构上,以α-螺旋为主,占44.16%,其次为无规则卷曲,占27.49%,而延伸链结构和β-转角所占比例较小(表1)。通过对甜菜BvAMT1-3基因跨膜结构域分析,发现BvAMT1-3 含有9 个跨膜螺旋区,属于跨膜蛋白(图4),存在一个Ammonium transport保守结构域,位置在15~442 aa(图5)。

图3 甜菜BvAMT1-3基因序列信息Fig.3 Gene sequence of Beta vulgaris BvAMT1-3

续图3 甜菜BvAMT1-3基因序列信息Continued Fig.3 Gene sequence of Beta vulgaris BvAMT1-3
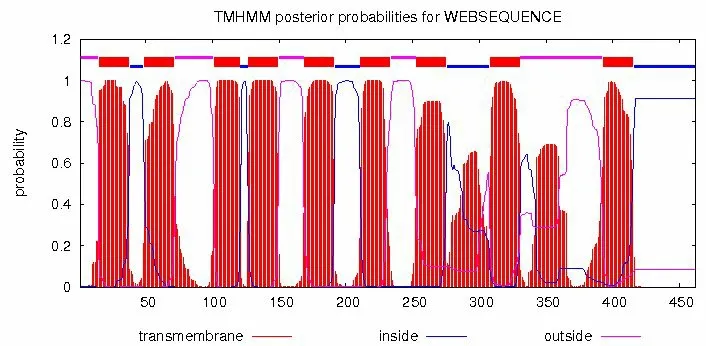
图4 BvAMT1-3蛋白的跨膜螺旋区分析Fig.4 Transmembrane helix region analysis of BvAMT1-3 protein

图5 甜菜BvAMT1-3蛋白保守结构域Fig.5 Conserved domain assay of BvAMT1-3 protein

表1 BvAMT1-3 蛋白的二级结构分析和亚细胞定位Table 1 Secondary structure and subcellular localization of BvAMT1-3 protein
2.2.2 BvAMT1-3蛋白的亲水性和疏水性预测
用ProtScale软件对蛋白质进行疏水性/亲水性分析,正值越大表示越疏水,负值越大表示越亲水,介于+0.5~-0.5 之间的主要为两性氨基酸。结果表明,BvAMT1-3 蛋白的疏水区域大于亲水区域(图6),预测该蛋白为疏水蛋白,这与ProtParam分析结果一致。
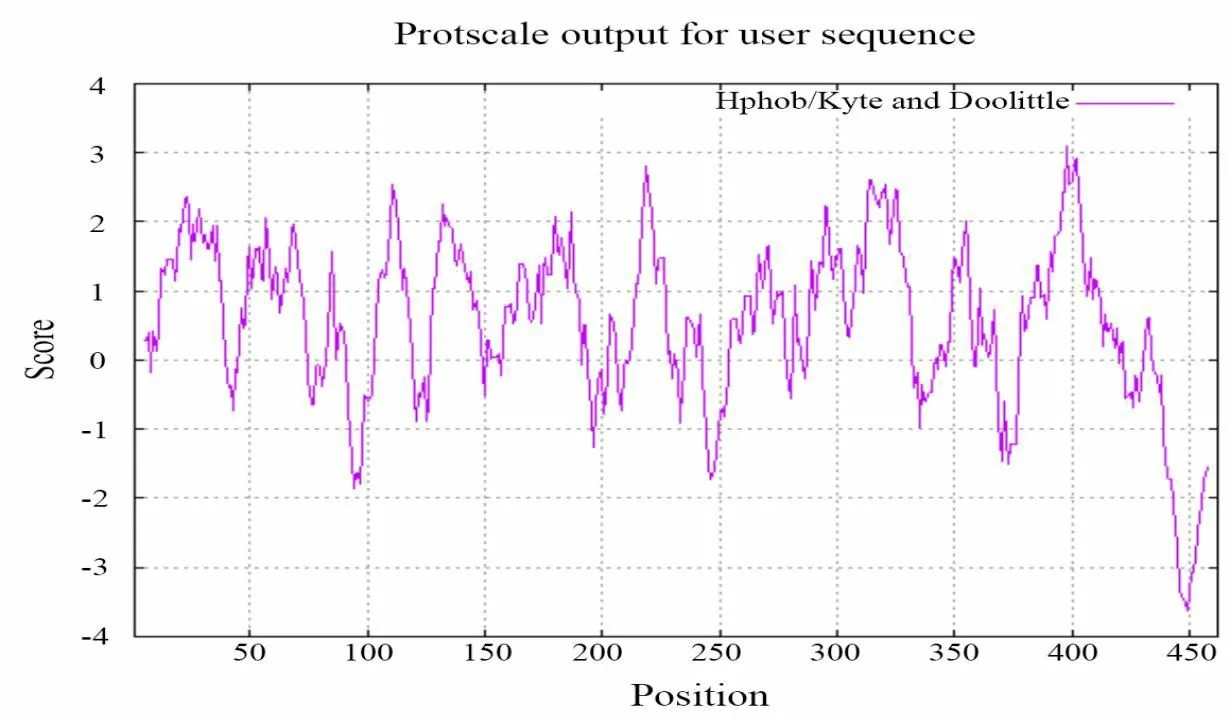
图6 BvAMT1-3蛋白亲疏水性预测Fig.6 Prediction of hydrophilicity and hydrophobicity of BvAMT1-3 protein
2.2.3 甜菜BvAMT1-3蛋白三级结构预测
将甜菜BvAMT1-3 的氨基酸序列提交到SWISSMODEL 在线软件,采用同源建模法,预测该蛋白的三级结构(图7)。结果显示,该蛋白的三级结构主要由无规则卷曲、延伸链和α-螺旋构成,与二级结构预测结果基本一致。
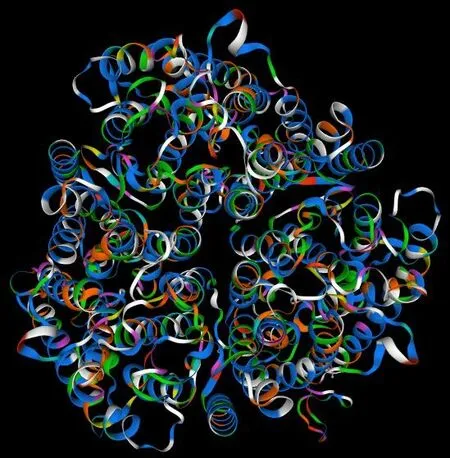
图7 BvAMT1-3蛋白的三级结构预测图Fig.7 Prediction for tertiary structure of BvAMT1-3 protein
2.2.4 甜菜BvAMT1-3基因启动子顺式作用元件分析
转录因子通过与启动子区域的顺式作用原件结合从而调控相关基因的表达,为进一步预测甜菜BvAMT1-3基因可能潜在的功能,利用Plantcare在线软件对甜菜BvAMT1-3基因启动子顺式作用元件进行分析。结果显示,该基因共预测到了5 类顺式作用元件(图8),包含有参与干旱诱导、赤霉素、胚乳表达、光反应、MeJA(茉莉酸乙酯)反应等顺式作用元件。
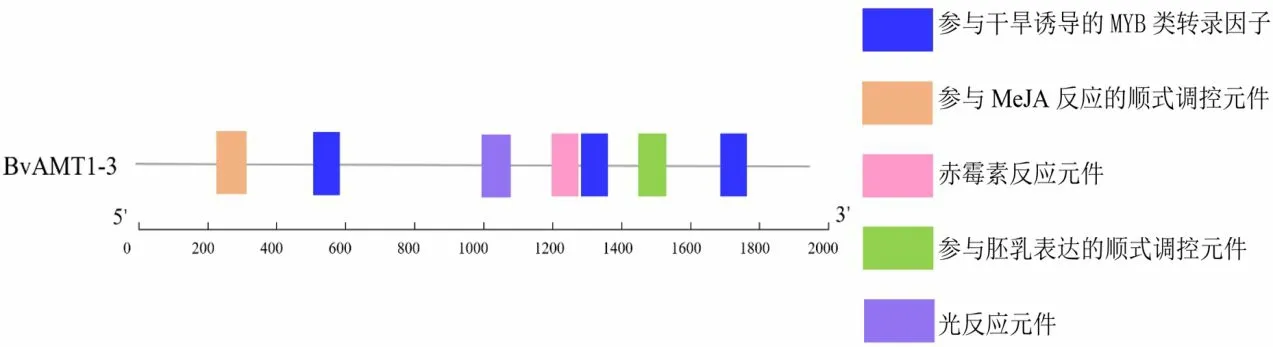
图8 甜菜BvAMT1-3基因启动子区域分析Fig.8 Promoter region analysis of BvAMT1-3 gene
2.2.5 甜菜AMTs基因系统进化树构建
通过系统进化树可以进一步分析不同物种间AMTs基因家族的亲缘关系,对选取的5 个甜菜AMTs、4 个黄花蒿AMTs、6 个拟南芥AMTs、4 个菠菜AMTs、5 个番茄AMTs和9 个水稻AMTs基因的氨基酸序列,利用MEGA7.0 进行多重序列比对构建了系统进化树(图9)。结果表明,来源于甜菜、拟南芥、菠菜、黄花蒿、番茄和水稻中的总共33 个AMTs基因被划分为AMT1、AMT2 和AMT3 这3 个亚族,其中甜菜的5 个AMT基因分为AMT1和AMT3两个亚家族。其中,仅有BvAMT3-3单独位于AMT3亚家族;包括BvAMT1-3在内的其余4个基因均为AMT1 亚家族。目的基因BvAMT1-3和SoAMT1-3聚为同一分支,表明BvAMT1-3与SoAMT1-3同源性较高。
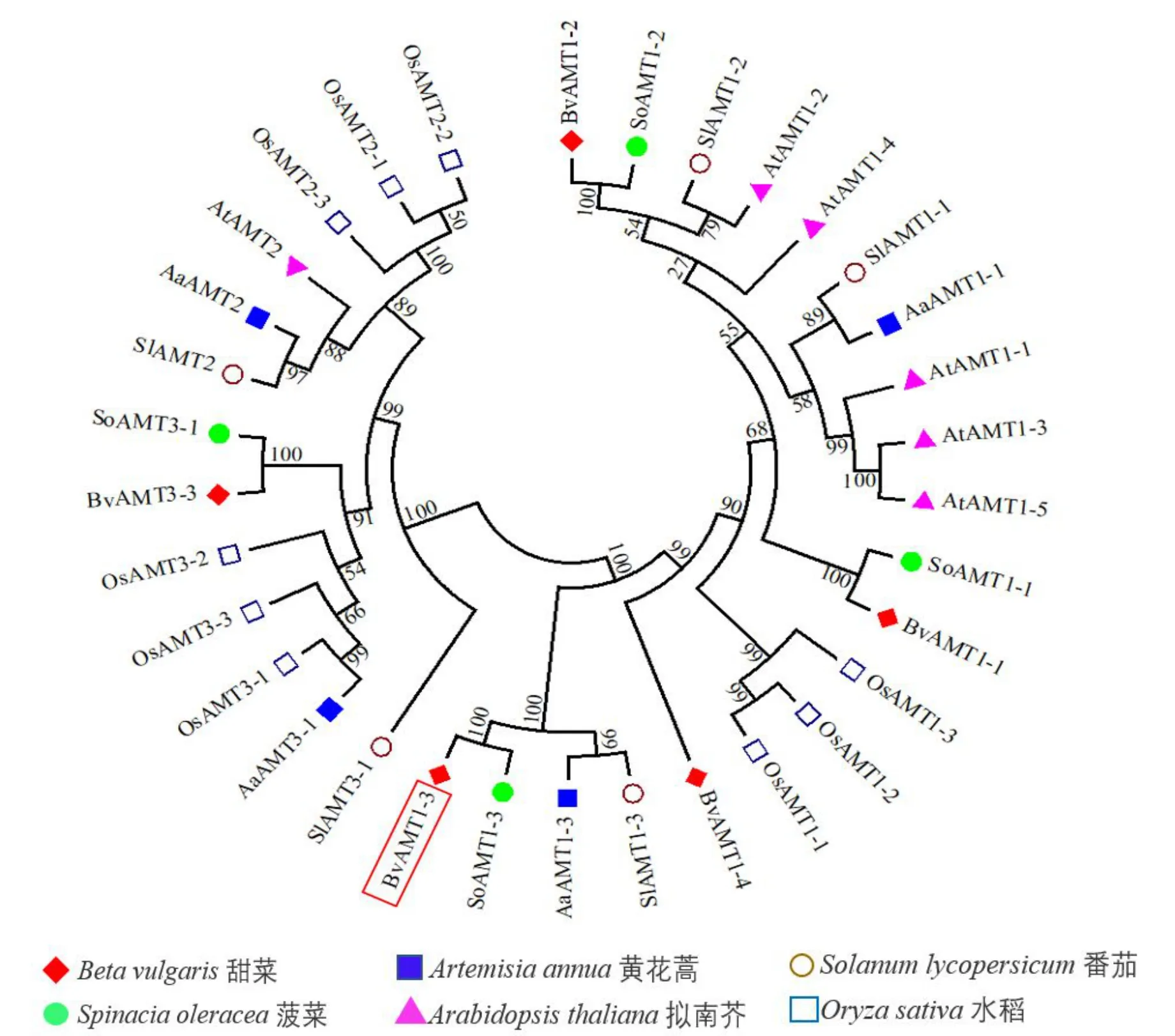
图9 甜菜与其它物种之间的AMT基因家族进化树Fig.9 Phylogenetic tree of the AMT genes family between Beta vulgaris and other species
3 讨论与结论
甜菜作为我国主要的产糖作物,被广泛种植于我国东北、西北和华北地区,是继甘蔗之后的第二大产糖作物[19]。糖的作用是多方面的,除了能作为食物提供能量参与人体的日常代谢外,也普遍应用于食品、酿酒、药品生产等工业。氮素是植物生长发育所需的一种重要营养物质,氮素是决定甜菜含糖的一个重要因素,甜菜通过铵转运蛋白(AMTs)调控对铵态氮的吸收和转运。
铵转运蛋白介导铵态氮在植物体内的转运机制主要是通过H+-ATP供给的细胞膜内外的渗透差。AMTs不仅介导植株对的吸收,同时也调控的外排,通过这一机制来维持植物体内的稳态[20]。而又可以通过反馈调节调控AMTs 的活性来控制植株氮代谢的平衡[21]。为了预测该基因的结构和特性,本研究利用RT-PCR技术克隆了BvAMT1-3基因,并运用生物信息学的方法对该基因进行了初步的分析。分析结果表明,甜菜中BvAMT1-3基因编码462个氨基酸,介于431~532个,这与之前大多数植物AMT 蛋白氨基酸个数的研究结果相一致[22]。蛋白质的α-螺旋和β-转角是二级结构中的稳定结构,是判断蛋白质结构稳定与否的关键,而无规则卷曲具有不稳定性,甜菜BvAMT1-3蛋白中α-螺旋和β-转角所占的比例大于无规则卷曲,因此说明该蛋白为稳定蛋白,也印证了理化性质分析的结果正确。BvAMT1-3基因存在Ammonium transport 保守结构域,这保证了BvAMT1-3 蛋白在转运铵盐中发挥稳定的作用。对甜菜中5 个AMTs基因的系统进化分析结果表明,甜菜BvAMT1-3基因与菠菜SoAMT1-3具有高度同源性,这一结果可能与植物进化史长久以来甜菜与菠菜的进化有关。对BvAMT1-3基因启动子顺式作用元件的预测结果表明,含有参与干旱诱导、生长素、胚乳表达、光反应、茉莉酸乙酯反应等顺式作用元件,说明该基因可能对甜菜生长和抗逆境胁迫有重要意义。
甜菜BvAMT1-3基因属于AMT1亚族,有报道表明,在AMT基因中存在特定的保守磷酸化位点。这种磷酸化位点在水稻和拟南芥的AMT1 家族中都被发现存在,其中有6 个保守磷酸化位点存在于拟南芥At-AMT1-1基因中[23]。通过对拟南芥AtAMT1-1的研究发现其中含有8 个蛋白激酶C 磷酸化位点和3 个酪蛋白激酶磷酸化位点[24]。这些磷酸化位点的存在保证了AMT1蛋白在铵态氮转运过程中与植物体内各器官之间的信号传导。研究表明,水葫芦EcAMT1在受到氮胁迫时的表达量会上调,棉花根系中GhAMT1-3表达量在低氮处理3d后升高,这与拟南芥、水稻、番茄和百脉根中的AMT1受氮胁迫而促进表达相匹配[25-26],表明氮胁迫可能会促进AMT1家族的基因表达。亚细胞定位显示其位于内质网膜,具有一定的转运功能。由此推断,该基因编码的蛋白可能会作为转运蛋白参与到甜菜对氮素的吸收和转运网络中,在受到氮胁迫时,该基因的表达量上调,增加植株对铵态氮的吸收从而降低甜菜受氮胁迫的损害。
目前对甜菜铵转运蛋白的研究还处于起步阶段,通过对甜菜BvAMT1-3基因进行克隆和进行生物信息学分析,可以对甜菜铵转运蛋白的功能进行初步研究,为深入了解甜菜铵转运蛋白在氮胁迫下转运铵态氮的分子机制和今后的转基因试验奠定基础。
猜你喜欢
土壤学报(2022年2期)2022-06-09
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
中国糖料(2022年2期)2022-04-06
小哥白尼(趣味科学)(2021年12期)2021-03-16
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
悦游 Condé Nast Traveler(2021年4期)2021-01-13
园艺与种苗(2020年12期)2021-01-08
农业环境科学学报(2020年10期)2020-11-11
环球时报(2017-09-19)2017-09-19
飞碟探索(2015年9期)2015-11-05
