
基于优化的MaxEnt模型预测赤水蕈树的潜在适宜区
2022-04-08 01:08文国卫叶兴状施晨阳赖文峰刘邦友蒋天雨朱晓如张国防
广西植物 2022年3期
关键词:环境因子
文国卫 叶兴状 施晨阳 赖文峰 刘邦友 蒋天雨 朱晓如 张国防


摘 要:赤水蕈樹(Altingia multinervis)具有较高的经济和药用价值,但由于气候变化和人类活动的强烈干扰,使其野生种质资源已近枯竭,被列为国家二级保护植物。为重建末次间冰期以来赤水蕈树地理分布格局变化过程、了解环境因子对潜在适宜区的制约机制,为赤水蕈树及其周围野生动植物的栖息地保护和引种栽培提供可靠的科学依据,该文运用MaxEnt模型结合地理信息系统ArcGIS软件模拟赤水蕈树潜在适生区及其空间变化格局,利用贡献率(PC)、置换重要值(PI)和刀切图评估制约现代地理分布的主导因子。结果表明:(1)优化的MaxEnt模型预测精度极高,AUC值大于0.97;赤水蕈树现代潜在适生区主要集中分布在贵州、四川和重庆交界的长江流域周围,其中以贵州赤水河流域为最佳适宜区;昼夜温差月均值(bio2)、年降水量(bio12)、降水季节性变化(bio15)和温度年较差(bio7)四个关键环境因子影响赤水蕈树的地理分布。(2)现代赤水蕈树潜在中高适生区面积2.692 6×104 km2,末次间冰期、全新世中期、未来时间段(2041—2060年、2061—2080年)低浓度(RCP2.6)和高浓度(RCP8.5)的中高适生区面积分别是2.277 3×104、2.831 0×104、2.159 6×104、2.605 1×104、2.330 4×104、2.460 4×104 km2。(3)在未来4种排放情景下,赤水蕈树新增的面积集中在四川和贵州区域,而在过去的全新世中期则集中分布在重庆区域。综上结果表明,赤水蕈树分布范围狭窄,而赤水河流域独特的地形优势可能是其避难的主要场所。
关键词: 赤水蕈树, MaxEnt模型, 潜在适生区, 环境因子, 避难所
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2022)03-0363-10
Potential suitable area of Altingia multinervis
predicted by optimizated MaxEnt model
WEN Guowei1, YE Xingzhuang1, SHI Chenyang1, LAI Wenfeng1, LIU Bangyou2,
JIANG Tianyu1, ZHU Xiaoru1, ZHANG Guofang1*
( 1. College of Forestry, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 2. Guizhou
Chishui Alsophila National Nature Reserve administration, Zunyi 564700, Guizhou, China )
Abstract:Altingia multinervis has been listed as a second-class protected plant in China., with high economic and medicinal values. However, its wild germplasm resources have been nearly exhausted due to the strong disturbance of climate change and human activities. Here, we aim to simulate the changes in the geographical distribution pattern of A. multinervis since the last interglacial (LIG) and to explore how climatic factors restrict the potential suitable areas, to provide a reliable scientific basis for habitat protection and cultivation of A. multinervis and its surrounding wildlife. In this study, based on optimized MaxEnt model and ArcGIS software, we simulated the potential suitable areas of A. multinervis and its spatial change pattern. The importance of environmental factors that constrains current geographical distribution was evaluated by percent contribution(PC), permutation importance(PI) and Jackknife test. The results were as follows: (1) The prediction accuracy of the optimized MaxEnt model was extremely high, as indicated by the value of the area under the receiver operator characteristic curve over 0.97. The potential suitable areas of A. multinervis for the present distribution were mainly distributed in the Yangtze River basin at the junction of Guizhou, Sichuan and Chongqing, of which the best suitable area was around Chishui River basin in Guizhou. The key environmental factors affecting the potential distribution of A. multinervis were mean diurnal range (bio2), annual precipitation (bio12), seasonality of precipitation (bio15) and range of annual temperature (bio7). (2) The potential middle-high suitable area of A. multinervis in current was 2.692 6×104 km2, in last interglacial (LIG) was 2.277 3×104 km2, in the mid-Holocene (MH) was 2.831×104 km2, in four future scenarios were 2.159 6×104 km2 (2050s RCP2.6), 2.605 1×104 km2 (2050s RCP8.5), 2.330 4×104 km2 (2070s RCP2.6), 2.460 4×104 km2 (2070s RCP8.5), respectively. (3) Under the four future discharge scenarios, the newly increased area of A. multinervis was concentrated in Sichuan and Guizhou, while it was concentrated in Chongqing in the mid-Holocene. All the above results indicate that the distribution range of A. multinervis is narrow and the potential suitable area is very small, and the unique topographic advantage of Chishui River Basin may be the main refuge place of A. multinervis.
Key words: Altingia multinervis, MaxEnt model, potential suitable area, environmental factors, refuge
近年来,全球气候变化和人类活动对环境、生物圈和生物多样性的影响日益显著,主要表现在降水的次数、强度和频率极不稳定,以及热浪、干旱、雷暴、洪水和飓风等极端事件的频发(D′Amato et al., 2020)。其中,极端气候事件强度和频率的增加,将直接对植物分布产生较大的负面影响(Mga et al.,2021)。由于气候变化的复杂性和不可预测性,以及地理位置的特殊性,植物对气候变化的适应就显得尤为重要(Ceccarelli & Grando, 2020)。为应对气候变化,特别是在气候环境恶劣的情况下,微避难所就极有可能是支持当地物种生存的栖息地(Mclaughlin et al., 2017)。目前,虽然对一些物种的灭绝机制已有新的进展,但仍未知哪些气候因子变化引起了物种灭绝,以及有多少物种会面临灭绝风險(Roman-Palacios & Wiens, 2020)。在气候和人为土地利用变化的双重作用下,植物生存和繁衍所需的栖息地类型大幅减少,并呈现碎片化(Sales et al., 2020; Karl et al., 2020)。而减缓气候变化和人类活动对原始森林生态系统的干扰,有益于物种间进行自然更新、基因交流和空间迁移等过程(Matteo et al., 2020)。因此,在气候变化的背景下保护生物多样性,需要制定出保护濒危物种切实可行的保护策略(Suggitt et al., 2018)。
全球气候变暖是21世纪生物多样性保护的重大挑战之一,而生态位模型(environmental niche models,ENMs)对气候变化下物种适宜潜在分布的变化性预测,已成为多个领域研究的热点(Li et al., 2020)。生态位模型在生态、进化和生物地理学中被广泛应用,因其能够对物种进行连续性预测和潜在适宜分布范围的变化描述(Galante et al., 2017;Mariano et al., 2019)。目前常用生态位模型主要有:最大熵模型(MaxEnt)(Sobek-Swant et al., 2012)、规则集遗传算法模型(GARP)(Zhang et al., 2020)、CLIMEX模型(Szyniszewska et al., 2020)等。其中,MaxEnt模型是最常用的小生境建模工具之一(Feng et al., 2019),是由于该模型对极少的样本数量(n≥5)就能较好地模拟出物种的适生范围,且成本低、操作简单、运行时间短(Pearson et al., 2007)。因此,MaxEnt模型常被国内外研究者应用于珍惜濒危动植物保护(Farashi & Shariati, 2017; Han et al., 2019)、物种入侵(Kariyawasam et al., 2020; 陈剑等,2021)、作物种植区划(Zhang et al., 2016)、病虫害防治(Li et al., 2019)等领域。
赤水蕈树(Altingia multinervis)是金缕梅科(Hamamelidaceae)蕈树属(Altingia)的常绿乔木之一,已被列为国家二级濒危保护植物(袁守良和白小节, 2009)。赤水蕈树树干挺直,木材质地坚硬,可供建筑、制作家具使用,也可砍倒作为优良的食用菌培养基和放养香菰的母树,其树皮流出的树脂可作香料和供药用,具有很好的经济和药用价值(Ye et al., 2020)。遗憾的是,对赤水蕈树还未进行系统和深入的研究,许多价值还未挖掘出来,该物种就已经处于极度濒危的状态,因此,对其开展试验研究并制定合理有效的管理保护措施刻不容缓。
末次间冰期(last interglacial,LIG)、末次盛冰期(last glacial maximum,LGM)和全新世中期(mid-holocene,MH)代表过去时期全球气候的三种极端状态,由于经过反复高温高湿-寒冷干燥-高温高湿的交替波动,对物种地理分布格局造成巨大影响,导致物种从高纬度向低纬度地区迁移,或低纬度向高纬度地区迁移,同时气候的剧烈变化严重阻碍了物种的进化,甚至导致物种大面积减少(Manthey & Box, 2010)。本研究利用R语言的ENMeval语言包优化MaxEnt模型,模拟末次间冰期、全新世中期、现代(current)和未来时间段(2041—2060年、2061—2080年)低浓度(RCP 2.6,温室气体排放浓度较低的情况下)和高浓度(RCP 8.5,温室气体排放浓度较高的情况下)两种气候情景下赤水蕈树的潜在适生区,探究其分布空间变化格局,进一步分析影响其分布的重要环境因子,以及探究该物种分布狭窄和能在一定范围内生存繁殖的原因,旨在为更好地开展赤水蕈树天然种质资源的保护提供参考和理论依据。
1 材料与方法
1.1 研究区域
赤水蕈树是贵州特有种植物,集中分布于赤水桫椤保护区的五柱峰、金沙沟,以及赤水的天台山,分布范围为105° 41′ 18″—106° 00′ 59″ E、28° 23′ 54″—28° 35′ 39″ N;海拔为600~1 000 m。该地区年降水量为1 200~1 300 mm,年均温度为16 ℃,相对湿度为70%~90%,森林覆盖率为90%以上,土壤以酸性的黄壤土和沙质土为主(袁守良和白小节, 2009)。鉴于赤水蕈树天然分布区域依然保持着原始的生态系统和分布窄的特点,可能还存在尚未被发现的区域,以及人为活动破坏等造成该物种的消失。因此,本研究适当扩大赤水蕈树的分布区域,为将来赤水蕈树的保护区设置提供参考。
1.2 数据的搜集与整理
赤水蕈树种群个体数量少,采集和收录样本的机构较少。本研究主要通过对赤水桫椤国家级自然保护区进行实地调查,并通过检索中国数字植物标本馆(CVH,http://www.cvh.ac.cn/)和查阅已出版的文献资料,获得赤水蕈树的天然分布记录。赤水蕈树的文献和标本中仅有分布地点的记录,便运用经纬度查询(http://map.jiqrxx.com/jingweidu/)定位,并对获得的数据进行处理,删除人工种植点、重复的点和经纬度缺失的点。此外,为降低群集效应所产生的误差,将分布点以物种名、经度、纬度和地点整理成CSV格式文件导入ArcGIS中,进行缓冲区分析,以1 km范围内的多个样本点,只保留一个点的原则,最终获得24个赤水蕈树天然种源分布样点。
1.3 环境因子筛选
本研究共有环境变量37个,分别为气候变量19个、土壤变量16个、海拔变量(elev)和人类活动变量(hfp)各1个。气候和海拔变量数据均来自WorldClim数据库(http://worldclim.org),土壤变量数据则来自于国家青藏高原科学数据中心(http://westdc.westgis.ac.cn)的“基于世界土壤數据库(HWSD)(http://www.fao.org/ faostat/en/#data.)的中国土壤数据集(V1.1)(2009)”,人类活动变量数据下载自国际地球科学信息网络中心(http://www.ciesin.org/)。其中,19个气候变量分别为年均温(bio1)、昼夜温差月均值(bio2)、等温性(bio3)、温度季节变动系数(bio4)、最热月最高气温(bio5)、最冷月最低气温(bio6)、温度年较差(bio7)、最湿季平均气温(bio8)、最干季平均气温(bio9)、最暖季平均气温(bio10)、最冷季平均气温(bio11)、年降水量(bio12)、最湿月降水量(bio13)、最干月降水量(bio14)、降水季节性变化(bio15)、最湿季降水量(bio16)、最干季降水量(bio17)、最暖季降水量(bio18)、最冷季降水量(bio19),16个土壤变量分别为表层土砾石含量(t-gravel)、表层沙含量(t-sand)、表层淤泥含量(t-silt)、表层粘土含量(t-clay)、土壤质地分类(t-usda-tex-class)、土壤容重(t-ref-bulk-density)、表层土壤有机碳含量(t-oc)、表层土壤酸碱度(t-pH-H2O)、粘性层土壤的阳离子交换能力(t-cec-clay)、土壤的阳离子交换能力(t-cec-soil)、表层土壤基本饱和度(t-bs)、表层土壤交换性盐基(t-teb)、表层土壤碳酸盐或石灰含量(t-caco3)、表层土壤硫酸盐含量(t-caso4)、表层土壤电导率(t-ece)、表层土壤容重(t-bulk-density)。由于赤水蕈树的分布范围狭窄,需较高空间分辨率的数据才能较好地模拟,却因末次盛冰期只有2.5′分辨率的气候数据,分辨率较低。因此,为了确保模拟结果的可靠性,放弃末次盛冰期的模拟,以保证所有时期的图层分辨率气候数据均为30″(30 arc-second)(地面约1 km2)。为减少因环境变量的多重共线性造成MaxEnt模型过度拟合的问题,将37个环境变量数据和最终分布点导入ArcGIS 10.5中进行点插值提取出来,在SPSS 22中对点插值数据做Spearman相关分析和多重共线性VIF方差膨胀因子分析,当两个环境因子之间的相关性大于等于0.8,只选择其中一个因子的原则,初步筛选出15个环境因子参与建模。
1.4 优化模型和预测精度评价
本研究采用R语言的ENMeval语言包优化MaxEnt模型(Phillips et al., 2017),即优化了调控倍频(regularization multiplier,RM)和特征组合(feature combination multiplier,FC)两个参数。其中,将调控倍频设置为0.5~8,依次递增0.5,共16个;并采用11个特征组合(L、QT、H、HP、PT、QH、LQH、LPT、QHP、LQPT和LQHPT),其中L、Q、H、P、T分别为线性(linear)、二次型(quadratic)、片段化(hinge)、乘积型(product)、阈值型(threshold)(朱耿平和乔慧捷,2016)。将上述176种参数组合进行综合测试,之后使用Akaike信息量准则(akaike information criterion correction, AICc)的delta.AICc模型评估OR10和AUCDIFF检验模型的拟合度和复杂度,最终可获取最低AICc值(delta.AICc=0)时的最佳模型参数组合,用于MaxEnt模型建模(Phillips et al., 2006)。然后,将掩膜好的ASCII文件和24个赤水蕈树现代分布点的CSV格式文件,通过MaxEnt 3.4.1软件来模拟预测不同时期、不同情景下赤水蕈树的潜在分布区和适生区变化,其中,75%的数据作为训练集(train set),剩下25%的数据作为验证集(test set),其他参数为默认值,重复10次(Robert et al, 2011)。根据赤水蕈树的适宜指数,将输出后平均值的ASCII文件导入ArcGIS 10.5,采用人工(manual)分级方法(刘艳梅等,2018),将适宜度分为高度适生区(0.7~1)、中度适生区(0.5~0.7)、低度适生区(0.3~0.5)和非适生区(0~0.3)4级,并利用ArcGIS栅格计算工具统计适生区面积。
其中,MaxEnt模型使用受试者工作特征曲线(receiver operating characteristic curve,ROC curves)的AUC(area under curve)值对模型进行准确度评价。AUC值范围在0到1之间,值越大说明预测的准确性就越好。若AUC < 0.7,则预测结果不理想,可信度低,一般不能使用;0.7~0.8为预测结果一般;0.8~0.9表示预测结果良好;0.9~1.0表示预测结果极佳(Jessica et al., 2014)。
1.5 适生区空间分布格局
首先,根据赤水蕈树的适宜指数将其分布存在概率值大于0.5作为适宜分布区,反之为非适宜区,分别赋值为“1”和“0”。然后,基于(0,1)矩阵,以现代潜在适生区为基础,进一步推测和分析赤水蕈树在过去和未来气候变化情景下适宜分布区的变化过程,并定义了四种适生区类型,依次为保留适生区(1→1)、新增适生区(0→1)、丧失适生区(1→0)和非适生区(0→0)。最后,将矩阵变换值导入ArcGIS 10.5,实现赤水蕈树适生区空间格局变化的可视化,并利用ArcGIS栅格计算工具统计保留区、增加区和丧失区的面积。
2 结果与分析
2.1 MaxEnt优化模型及准确性评价
本研究以24个赤水蕈树现代分布点和筛选后的15个环境变量图层为基础,并根据AICc信息准则,使用MaxEnt模型对其过去、现代和未来不同情景下的潜在分布区进行模拟。其中,15个环境变量分别是昼夜温差月均值、等温性、温度季节变动系数、温度年较差、最湿季平均气温、年降水量、最干月降水量、降水季节性变化、最干季降水量、土壤质地分类、土壤容重、表层土壤有机碳含量、表层土壤酸碱度、海拔、人类活动。根据R语言获得的AIC信息量准则可知,当delta. AICc=0时,FC= HP,RM= 6.5,为此次模拟中最优的参数。然后,使用优化后的参数进行MaxEnt模拟,预测现代赤水蕈树潜在地理分布,在10次重复中,训练AUC平均值为0.980 0 ± 0.004 8,而测试AUC平均值为0.970 8 ± 0.010 5,两者的AUC值均大于0.9,表明优化后的MaxEnt模型能够准确模拟赤水蕈树的潜在地理分布。
2.2 环境变量的重要性
环境变量的重要性主要由贡献率(percent of contribution,PC)和置换重要值(permutation importance,PI)指标来评定,环境变量指标数值越大,就越重要。基于赤水蕈树现代分布点和15个环境变量图层的MaxEnt模拟结果,因有5个环境变量的贡献率和置换重要值为零或极小,因此,该研究仅提供10个主导环境变量的参数(表1)。贡献率前5个因子依次是昼夜温差月均值(bio2,36.09%)、年降水量(bio12,25.38%)、降水季节性变化(bio15,12.29%)、温度年较差(bio7,9.50%)和表层土壤酸碱度(t-pH-H2O,3.82%),共占总贡献率的77.08%。置换重要值前5个因子依次是昼夜温差月均值(39.41%)、年降水量(19.22%)、降水季节性变化(16.34%)、最干月降水量(bio14,7.69%)和温度年较差(bio7,6.91%),共占置换重要值的89.57%。在刀切图中(图1),使用仅此变量(with only variable)分析时,AUC值最佳的环境因子是昼夜温差月均值,其后三个因子分别是等温性(bio3)、最干月降水量(bio14)和最干季度降水量(bio17),表明这四个环境因子对赤水蕈树的分布有很大的影响。综上,昼夜温差月均值(bio2)和年降水量(bio12)是影响赤水蕈树现代潜在分布的主导因子,降水季节性变化(bio15)、温度年较差(bio7)、最干月降水量(bio14)和表层土壤酸碱度(t-pH-H2O)对赤水蕈树的潜在分布也起着重要作用。
2.3 现代、过去和未来潜在适生区
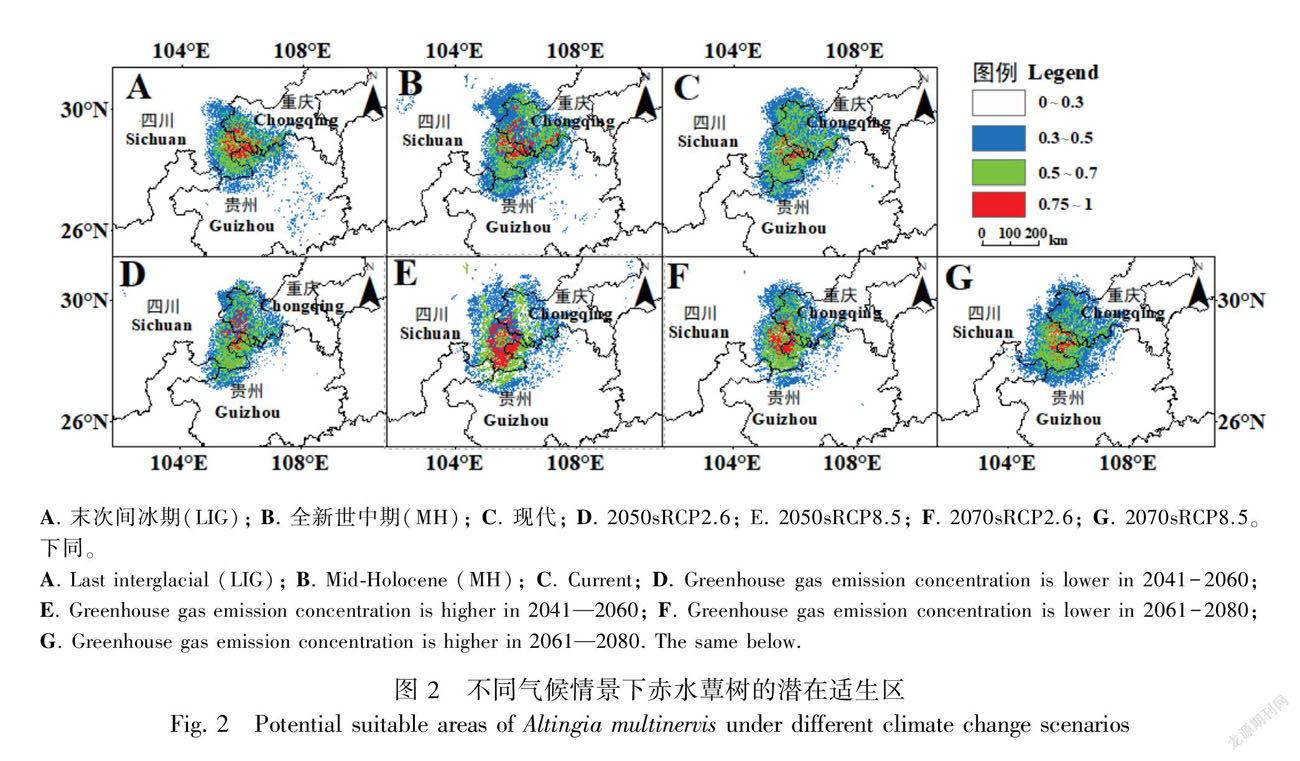
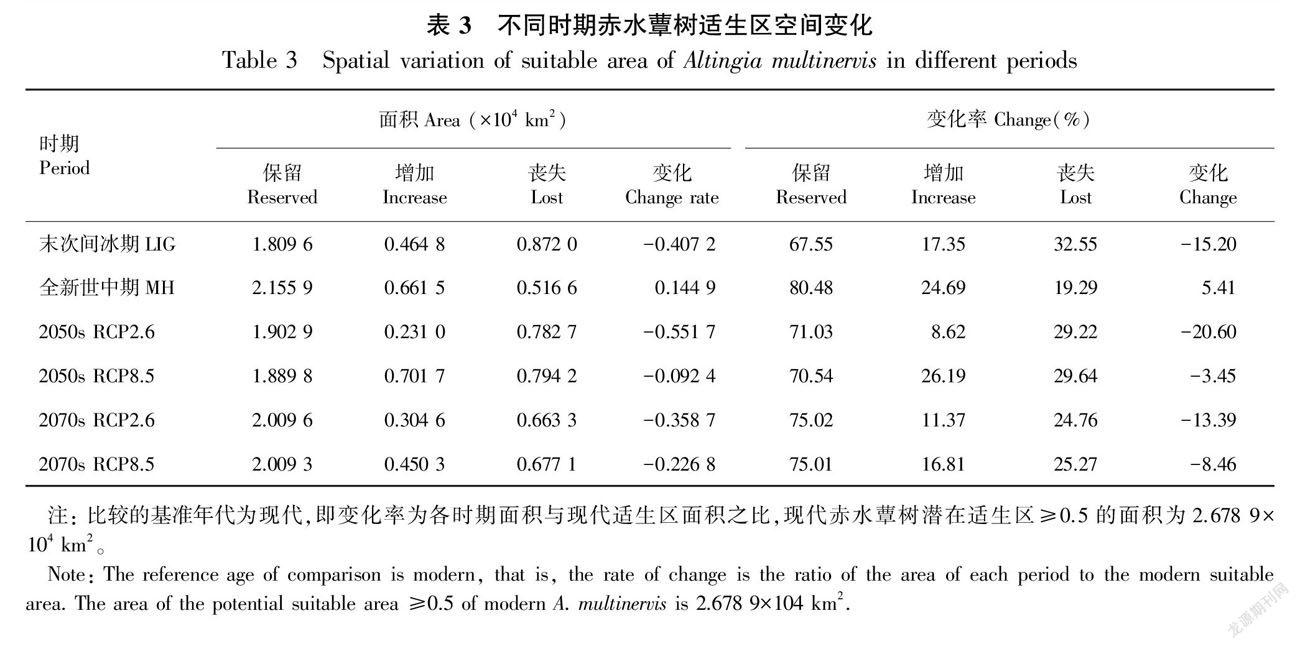
根据MaxEnt模拟结果,赤水蕈树现代潜在总适生区面积为6.547 6×104 km2,包括高適生区0.252 7×104 km2、中适生区2.439 9×104 km2、低适生区3.854 9×104 km2,潜在适生区主要集中分布在贵州、四川和重庆交界的长江流域周围,其中又以赤水河流域分布最为密集(表2,图2:C)。从末次间冰期到现代,赤水蕈树潜在总适生区由最初的5.409 4×104 km2(LGM)增加到7.540 3×104 km2(MH),然后下降到6.547 6×104 km2(current);同样,高适生区从0.535 3×104 km2(LGM)增加到0.579 7×104 km2(MH),之后大幅下降到0.252 7×104 km2(current),比过去两个时期少了两倍多(表2,图2:A-C)。未来4种不同气候情景(2050s RCP2.6、RCP8.5,2070s RCP2.6、RCP8.5)下,赤水蕈树总适生区分别为5.170 4×104、6.413 1×104、5.352 6×104、6.999 7×104 km2,这表明较高浓度的排放情景有利于赤水蕈树适生区范围的扩大。
2.4 过去和未来空间格局变化
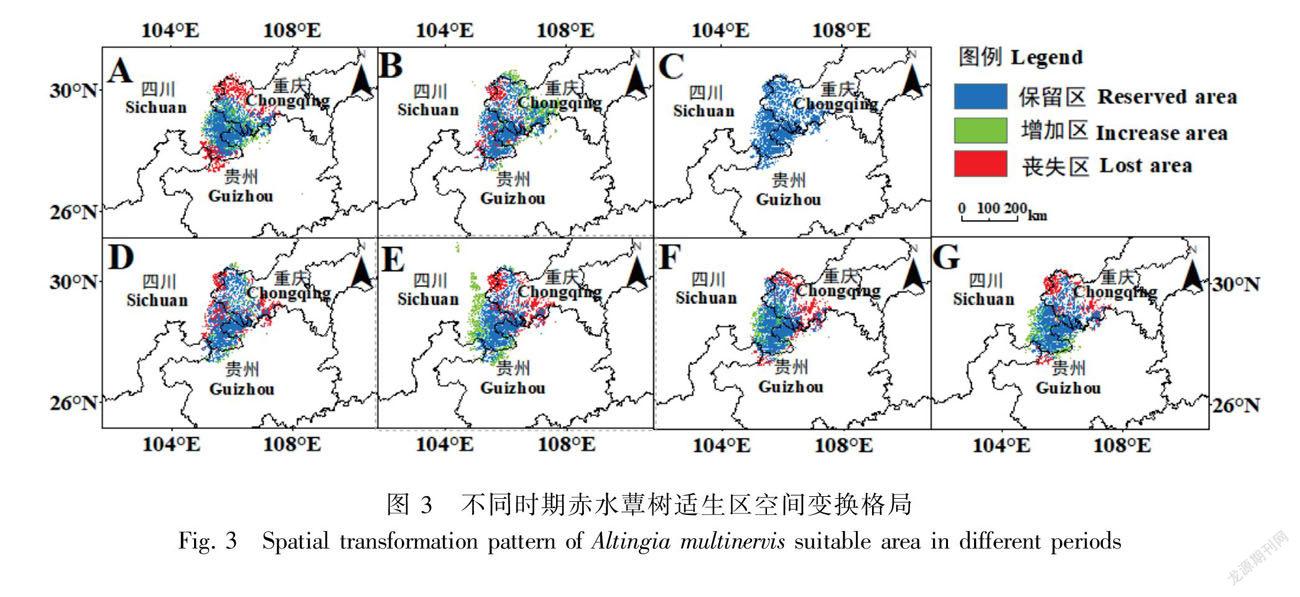
基于现代赤水蕈树的潜在分布范围,末次间冰期潜在适生区的丧失面积最大,为0.872 0×104 km2,占现代潜在适生区面积的32.55%,丧失面积主要分布在现代赤水蕈树的潜在适生区的南北两端(图3:A,表3)。此外,除全新世中期外,末次间冰期、2050s RCP2.6、2050s RCP8.5、2070s RCP2.6和2070s RCP8.5的潜在适生区新增面积均小于丧失面积,分别丧失了0.407 2×104、0.551 7×104、0.092 4×104、0.358 7×104、0.226 8×104 km2,依次占现代潜在适生区面积的15.20%、20.60%、3.45%、13.39%、8.46%(表3)。全新世中期新增面积主要在集中在重庆部分,与其他时期相反。在未来两个时期两种不同浓度排放情景下,赤水蕈树潜在适生区丧失的面积主要集中在重庆部分区域,而新增的面积集中在四川和贵州部分区域(图3)。值得注意的是,6个时期中,末次间冰期潜在适生区新增面积小于丧失面积(变化率15.20%),且总体潜在适生区的北部比南部丧失面积更多,而全新世中期的潜在适生区新增面积大于丧失面积(变化率5.41%),增加部分主要集中在现代赤水蕈树的潜在适生区的北端(重庆部分)(表3,图3:A、B),这表明全新世中期更有利于赤水蕈树的生存和繁殖。此外,未来新增的面积均小于丧失的面积,低浓度排放情景(RCP2.6)丧失面积由2050s的20.60%下降到2070s的13.39%,而高浓度排放情景(RCP2.6)丧失面积由2050s的3.45%扩大到2070s的8.46%(表3),这表明长期的RCP2.6排放情景或短期内的RCP8.5排放情景下,赤水蕈树的潜在适生区丧失面积有下降的趋势。
3 讨论与结论
3.1 赤水蕈树潜在地理分布及其主导环境因子
基于赤水蕈树现代分布点,使用MaxEnt模型预测了其潜在适生区范围,除了贵州赤水河流域为最佳适生区外,还在贵州与四川、重庆交界的部分区域有一定零散分布。由于赤水蕈树本身对生态环境的要求极为苛刻,常与其他植物共生(袁守良和白小节, 2009),模拟范围比实际要大一些,这与Lin等(2020)对12种白松的模拟结果相一致。此外,本研究发现赤水蕈树可能更适宜较高浓度的排放情景下的气候条件,且赤水河流域独特的地形优势极有可能是赤水蕈树避难的主要场所。正是因为赤水蕈树本身的特性和所在生境的特殊性,使其分布范围极为狭窄。因此,在不同时期,影响赤水蕈树分布的土壤因子基本相同,不会有较大波动;而造成赤水蕈树分布区变化可能的原因是气候因子的剧烈波动,以及人类活动的强烈干扰等。
与现代相比,气候变化对赤水蕈树未来潜在适宜区的影响,主要体现在在未来两种排放情景下,总适生区均为增加趋势。其中,高度适生区范围在低浓度(RCP2.6)排放情景下随着时间的推移而有所增加,高浓度(RCP8.5)排放情景则相反。根据MaxEnt模拟结果,昼夜温差月均值(bio2)、年降水量(bio12)、降水季节性变化(bio15)和温度年较差(bio7)4个因子对赤水蕈树的分布起到了主导作用,与冯雪萍等(2017)对樟科(Lauraceae)、壳斗科(Fagaceae)和大戟科(Euphorbiaceae)等5科物候期的研究结论相一致。另外,人类对土地利用变化是碳排放的主要来源之一,也是气候变化的驱动因素(Tang et al., 2021)。但本研究中,人类活动因子并未成为影响赤水蕈树潜在分布的主导因子之一,这可能是因为人类活动是间接影响赤水蕈树的潜在分布,具体原因还需进一步分析和探讨。
3.2 赤水蕈树种质资源的保护
现代赤水蕈树适生区在过去、未来气候变化下仍有共同部分(保留区)。保留区作为赤水蕈树分布的较高适宜区域,在过去已证明保留区是气候变化条件下赤水蕈树的避难所,而在未来气候变化条件下,保留区既能作为赤水蕈树应对气候变化的生长地与避难所,又可作为赤水蕈树试验研究的主要区域。因此,保留区内应做到两点:其一,应加强赤水蕈树野生种质资源的保护和管理,实现赤水河周围脆弱生态环境的保护,以提高该地的生物多样性,更好地发挥自身的生态效益;其二,加强赤水蕈树高适宜分布区的栖息地管理工作,注重因地制宜地营造满足其生存和繁衍后代的需求。对于增加区,除全新世中期主要分布于适宜分布区的重庆边界地域外,其他时期均主要分布于适宜分布区的四川和贵州边界地域,应在增加制定可持续的土地利用规划,并在新增适生区的边缘地带,减少此范围内的人为干扰强度和频率,为赤水蕈树的传播提供足够适宜的空间,以增加赤水蕈树分布范围扩大的可能性。对于丧失区来说,主要集中分布在赤水蕈树适宜分布区的重庆边缘地带,此范围内应以野生资源保护为主,积极开展赤水蕈树的资源调查和动态监测活动,提高赤水蕈树对气候变化的适应能力,同时将该范围内的赤水蕈树科学迁入其适宜种植范围,进行迁地保护。此外,迁地保护作为珍稀濒危野生种质资源最重要的科学保护策略之一,这對濒危植物的进一步研究和近缘种起源和进化的探索具有十分重要的意义(Xiang et al., 2018)。总之,赤水蕈树的保留区、增加区和丧失区分别相当于保护区的核心区、缓冲区和试验区,这对保护区的设置提供了大概的结构框架,在未来对营建全球的生物多样性都具有重大的意义。
赤水蕈树等极小种群的分布现状是由全球气候变化、现代人类社会发展和所生长的地理环境条件,以及本身的特性共同造成的,可能出现当前生境并不是其最佳适生区。因此,开展当前气候情境下赤水蕈树的潜在分布范围调查,进一步分析其潜在地理空间分布,探寻其种群传播的可替代区域,从而扩大种植面积,为就地和迁地保护打好基础。王卫等(2019)利用MaxEnt模型预测极小种群植物丹霞梧桐的潜在适生区,并分析影响其分布的主导因子,对保护区就地和迁地保护的措施制定提供依据,这与本研究的初衷具有一致性。赤水蕈树生长倾向于人迹罕至的山地地区,最适分布集中于赤水河附近的赤水桫椤国家级自然保护区内,且模拟的潜在分布区与该描述吻合度高。根据模拟结果与分析,我们建议可在贵州赤水桫椤自然保护区、赤水竹海国家森林公园、雷公山国家级自然保护区、习水国家级自然保护区、梵净山国家级自然保护区、重庆金佛山国家级自然保护区、重庆四面山自然保护区和四川黄荆老林国家森林公园率先展开迁地工作。
参考文献:
CECCARELLI S, GRANDO S, 2020. Evolutionary plant breeding as a response to the complexity of climate change[J]. iScience, S2589-0042(20): 31012
CHEN J, WANG SH, ZHU F, et al, 2021. Study on the risk of invasive plant Pachyderma in Yunnan [J]. Guihaia, 41(5); 789-798.[陈剑, 王四海, 朱枫, 等, 2021. 外来入侵植物肿柄菊在云南的扩散风险研究[J]. 广西植物, 41(5); 789-798.]
D′AMATO G, HERBERTO JC, ORTEGA OPM, et al., 2020.
The effects of climate change on respiratory allergy and asthma induced by pollen and mold allergens[J]. Allergy, 75(9): 2219-2228.
FARASHI A, SHARIATI M, 2017. Biodiversity hotspots and conservation gaps in Iran[J]. J Nat Conserv, 39: 37-57.
FENG X, PARK DS, LIANG Y, et al., 2019.Collinearity in ecological niche modeling: Confusions and challenges[J]. Methods Ecol Evol, 9(3): 10365-10376.
FENG XP, LIU JF, HE ZS, et al., 2017. Effects of the climate factors on phenological phase of flowers and fruits of the main woody plants in Dai Yun Mountain [J]. J NW For Univ, 32(3): 56-61. [冯雪萍, 刘金福, 何中声, 等, 2017. 气候因子对戴云山木本优势科植物花果期物候的影响[J]. 西北林学院学报, 32(3): 56-61.]
GALANTE PJ, ALADE B, MUSCARELLA R, et al., 2017. The challenge of modeling niches and distributions for data-poor species: a comprehensive approach to model complexity[J]. Ecography, 41(5): 726-736.
HAN YH, DONG SK, WU XY, et al., 2019. Integrated modeling to identify priority areas for the conservation of the endangered plant species in headwater areas of Asia[J]. Ecol indic, 105(Oct.): 47-56.
JESSICA BS, CHRISTINE RW, KAREN VR., 2014. Developingmacrohabitat models for bats in parks using maxent and testing them with data collected by citizen scientists[J]. Int J Biodivers Conserv, 6(2): 171-183.
KARIYAWASAM CS, KUMAR L, RATNAYAKE SS, 2020. Potential risks of plant invasions in protected areas of Sri Lanka under climate change with special reference to threatened vertebrates[J]. Climate, 8(4): 51.
KARL H,KUTTNER M, MOSER D, et al., 2020. Habitat availability disproportionally amplifies climate change risks for lowland compared to alpine species[J]. Glob Ecol Conserv, 23: e01113.
LI AN, WANG JW, WANG RL, et al., 2019. MaxEnt modeling to predict current and future distributions of Batocera lineolata (Coleoptera: Cerambycidae) under climate change in China[J]. Ecoscience, 27(1): 1-9.
LI X, LI M, HOU L, et al., 2018. De novo transcriptome assembly and population genetic analyses for an endangered Chinese endemic Acer miaotaiense (Aceraceae)[J]. Genes, 9(8): 378.
LI Y, LI M, LI C, et al., 2020. Optimized MaxEnt model predictions of climate change impacts on the suitable distribution of Cunninghamia lanceolata in China[J]. Forests, 11(3): 302.
LIN L, HE J, XIE L, et al., 2020. Prediction of the suitable area of the Chinese white pines (Pinus subsect. Strobus) under climate changes and implications for their conservation[J]. Forests, 11(9): 996.
LIU YM, ZHOU SD, XIE DF, et al., 2018. Potential distribution of Fritillaria unibracteata predicted by the MaxEnt model [J]. Guihaia, 38(3): 352-360. [劉艳梅, 周颂东, 谢登峰, 等, 2018. 基于最大熵模型(MaxEnt)预测暗紫贝母的潜在分布[J]. 广西植物, 38(3): 352-360.]
MANTHEY M, BOX EO, 2010. Realized climatic niches of deciduous trees: comparing western Eurasia and eastern North America[J]. J Biogeogr, 34(6): 1028-1040.
MARIANO SG, CAROLINA CA, ANDERSON RP, 2019. Sufficient versus optimal climatic stability during the Late Quaternary: using environmental quality to guide phylogeographic inferences in a Neotropical montane system[J]. J Mammal, 100(6): 1783-1807.
MATTEO P, MAURIZIO M, MARCO M, et al., 2020. Potential impact of climate change on the forest coverage and the spatial distribution of 19 key forest tree species in Italy under RCP4.5 IPCC Trajectory for 2050s[J]. Forests, 11(934): 1-19.
MCLAUGHLIN BC, ACKERLY DD, KLOS PZ, et al., 2017. Hydrologicrefugia, plants, and climate change[J]. Glob Change Biol, 23(8): 2941-2961.
MGA B, AHC D, PC A, et al., 2021. Are pine-oak mixed stands in Mediterranean mountains more resilient to drought than their monospecific counterparts? — Science Direct[J]. For Ecol Manag, 484: 118955.
PEARSON RG, RAXWORTHY CJ, NAKAMURA M, et al., 2007. ORIGINAL ARTICLE: Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar[J]. J. Biogeogr, 34(1): 102-117.
PHILLIPS SJ, ANDERSON RP, MIROSLAV D, et al., 2017. Opening the black box: An open-source release of MaxEnt[J]. Ecography, 40(7): 887-893.
PHILLIPS SJ, ANDERSON RP, SCHAPIRE RE, 2006. Maximum entropy modeling of species geographic distributions[J]. Ecol Model, 190(3-4): 231-259.
ROBERTO M, ZAMORA R, MOLINA JR, et al, 2011. Predictive modeling of microhabitats for endemic birds in South Chilean temperate forests using maximum entropy(Maxent)[J]. Ecol Inform, 6: 364-370.
ROMAN-PALACIOS C, WIENS JJ, 2020. Recent responses to climate change reveal the drivers of species extinction and survival[J]. Proc Natl Acad Sci USA, 117(8): 201913007.
SALES L, GALETTI M, PIRES M, 2020. Climate and land-use change will lead to a faunal ‘savannization’ on tropical rainforests[J]. Glob Change Biol, 26(12): 7036-7044.
SOBEK-SWANT S, KLUZA DA, CUDDINGTON K, et al., 2012. Potential distribution of emerald ash borer: What can we learn from ecological niche models using Maxent and GARP?[J]. For Ecol Manag, 281(none): 23-31.
SUGGITT AJ, WILSON RJ, ISAAC N, et al., 2018. Extinction risk from climate change is reduced bymicroclimatic buffering[J]. Nat Clim Change, 8(8): 713-717.
SZYNISZEWSKA AM, LEPPLA NC, MANOUKIS NC, et al., 2020. CLIMEX and MED-FOES models for predicting the variability in growth potential and persistence of Mediterranean fruit fly (Diptera: Tephritidae) Populations[J]. Ann Entomol Soc Am, 113(2): 114-124.
TANG X, WOODCOCK CE, OLOFSSON P, et al., 2021. Spatiotemporal assessment of land use/land cover change and associated carbon emissions and uptake in the Mekong River Basin[J]. Remote Sens Environ, 256: 112336.
WANG W, YANG JJ, LUO XY, et al., 2019. Assessment of potential habitat for Firmiana danxiaensis,a plant species with extremely small populations in Danxiashan National Nature Reserve based on MaxEnt model [J]. Sci Silv Sin, 55(8): 19-27. [王卫, 杨俊杰, 罗晓莹, 等, 2019. 基于MaxEnt模型的丹霞山国家级自然保护区极小种群植物丹霞梧桐的潜在生境评价[J]. 林业科学, 55(8): 19-27.]
YE XZ, ZHANG MZ, JIANG YT, et al., 2020. The complete chloroplast genome of Altingia chinensis (Hamamelidaceae)[J]. Mitochondrial DNA B, 5(2): 1808-1809.
YUAN SL, BAI XJ, 2009. Distribution and conservation strategies of the rare plant Altingia multinervis[J]. Environ Protect Technol, 15(1): 42-43. [袁守良, 白小节, 2009. 珍稀植物赤水蕈树的分布及保护对策探索[J]. 环保科技, 15(1): 42-43.]
ZHANG K, SUN L, TAO J, 2020. Impact of climate change on the distribution of Euscaphis japonica (Staphyleaceae) Trees[J]. Forests, 11(5): 525.
ZHANG LL, CAO B, BAI CK, et al., 2016. Predicting suitable cultivation regions of medicinal plants with MaxEnt modeling and fuzzy logics: A case study of Scutellaria baicalensis in China[J]. Environ Earth Sci, 75(5): 361.
ZHU GP, QIAO HJ, 2016. Effect of the MaxEnt model’s complexity on the prediction of species potential distributions [J]. Biodivers Sci, 24(10): 1189-1196. [朱耿平, 乔慧捷, 2016. MaxEnt模型复杂度对物种潜在分布区预测的影响[J]. 生物多样性, 24(10): 1189-1196.]
(責任编辑 李 莉)
收稿日期:2021-05-09
基金项目:福建农林大学科技创新基金(KFA17069A);宁夏自治区重点研发计划项目(KJA19H01A);福州定西水土流失综合治理项目(KH180062A)[ Supported by Science and Technology Innovation Fund of Fujian Agriculture and Forestry University(KFA17069A); Key Research and Development Project of Ningxia Autonomous Region(KJA19H01A); Fuzhou Dingxi Project of Soil Erosion Comprehensive Control (KH180062A)]。
第一作者: 文国卫(1997-),硕士,主要从事濒危物种保护研究,(E-mail)1200428018@fafu.edu.cn。
通信作者:张国防,教授,博士生导师,主要从事森林培育、经济林栽培和生态学等相关研究,(E-mail)fjzgfzgf@126.com。
猜你喜欢
湖北农业科学(2017年3期)2017-03-21
江苏农业科学(2016年11期)2017-03-21
中国中药杂志(2017年1期)2017-03-06
中国市场(2016年36期)2016-10-19
天津农业科学(2016年4期)2016-04-20
天津农业科学(2015年12期)2015-12-03
河北渔业(2015年10期)2015-10-15
湖北农业科学(2014年1期)2014-03-22
