
濒危种观光木小枝生物量分配与功能性状的纬度变异规律及其影响因素
2022-04-08 01:08蒙检罗应华于瀛刘朝阳林建勇
广西植物 2022年3期
蒙检 罗应华 于瀛 刘朝阳 林建勇




摘 要:為了探讨观光木当年生枝条生物量的分配规律和叶片功能性状的变化规律及其影响因素,该研究以濒危物种观光木(Michelia odora)为对象,测定了广西地区5个不同纬度上观光木当年生小枝及叶片功能性状。结果表明:(1)随着纬度的增加小枝总重和总叶重总体呈异速生长关系,其生物量更多趋于对小枝的构建。(2)观光木叶功能性状呈现一定规律,即从南向北随纬度的增加,其叶面积、叶重呈逐渐降低的趋势,叶厚度、叶干物质含量、气孔长、气孔大小、气孔密度呈逐渐上升的趋势,比叶面积、叶含水率、气孔宽先增大后减小,总体呈逐渐减低的趋势,叶组织密度先减小后增大,反映观光木可通过叶片形态可塑性以应对地理环境的变化。(3)比叶面积与叶含水率呈显著正相关,而与叶鲜重、叶干物质含量、气孔密度呈显著负相关;叶干物质含量与叶含水率呈显著负相关,说明观光木可以通过叶片性状组合的调整和平衡从而响应环境变化。(4)CCA分析显示,坡度是影响观光木功能性状的最显著变量,年平均温度对观光木功能性状也具有显著影响,年均降水量和相对湿度对植物功能性状具有一定强度的影响,但不起显著作用。该研究结果有助于理解观光木对环境的适应特征,其对纬度变异的响应机制等具有重要意义。
关键词: 小枝, 纬度, 生物量分配, 功能性状, 环境因子
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2022)03-0394-12
Latitudinal variation of branchlet biomass allocation and
functional traits of endangered species Michelia odora
and its influencing factors
MENG Jian1, LUO Yinghua1,2*, YU Ying1, LIU Zhaoyang1, LIN Jianyong3
( 1. Forestry College of Guangxi University, Nanning 530004, China; 2. Forestry College of Guangxi University, Guangxi Key Laboratory of
Forest Ecology and Conservation, Nanning 530004, China; 3. Guangxi Zhuang Autonomous Region
Forestry Research Institute, Nanning 530002, China )
Abstract:In this study, the functional traits of annual branchlets and leaves of Michelia odora, an endangered species, were sampled and measured along five different latitude gradients in Guangxi. The variation of biomass distribution and leaf functional traits along the latitude gradient were discussed. The results were as follows: (1)The total branchlet weight and total leaf weight tended to allometric growth relationship with the increase of latitude, and its biomass tended to the construction of branchlets. (2) The functional traits of the leaves showed a certain rule, i. e. increased with latitude from south to north, and its leaf area and leaf mass decrease of with the increasing latitude; leaf thickness, leaf dry matter content, stomatal length, stomatal area, stomatal density showed a trend of gradual increase; specific leaf area, leaf water content and stomatal width increased at first and then decreased, and overall showed a trend of gradual decrease, leaf tissue density decreased and then increased, which indicates that M. odora can respond to changes in the geographical environment through the plasticity of the leaf shape. (3) Specific leaf area was significantly positively correlated with leaf moisture content, but negatively correlated with leaf fresh weight, leaf dry matter content, and stomatal density. There was a significantly negative correlation between leaf dry matter content and leaf water content, which means that M. odora can respond to the change of geographical environment by adjusting and balancing the character combination of leaves. (4) CCA analysis showed that slope was the most significant variable affecting the functional traits of M. odora, the mean annual temperature also had a significant impact on the functional traits of M. odora, the mean annual precipitation, and relative humidity had a certain intensity effect on plant functional traits, but not significant. All the results are helpful to understand the adaptation characteristics of M. odora to the environment, and its response mechanism to latitude variation is of great significance.
Key words: twig, latitude, biomass allocation, functional traits, environmental factors
植物与环境之间的相互关系一直都是生态学研究的核心问题之一。植物通过对小枝、叶片和一些性状的调节形成对环境变化的适应。植物小枝生物量分配的权衡,影响小枝结构构建、养分的传输及吸收效率,反映植物生活史进化的生态策略(Dai et al., 2020)。生境变化对植物的分布规律、功能性状乃至生态系统功能等产生重大影响(Collins et al., 2016)。植物叶片具有很大的表型可塑性,叶片形态和解剖结构特征随着光照、水分和温度等环境因素的变化而形成不同类型的适应性(Niu et al., 2018)。植物叶功能性状是决定植物生理生化循环的关键因素(Roa-Fuentes et al., 2015),它与植物对资源的获取和利用密切相关(Zirbel et al., 2017)。植物各功能性状不是独立发挥作用的,各性状间存在一定的相关性(何念鹏等,2018a)。揭示各性状间的关联性,有助于了解植物适应外界环境生长、生存的策略(Guittar et al., 2016)。当年生小枝是植物分枝系统中最具有活力的部分,植物叶片和果实大都生长在这些小枝(twig)上,研究其特性对于理解植物生理生态变化和植物的生活史策略具有重要意义(杨冬梅等,2011)。植物通过权衡资源配置策略,以适应不同生长发育阶段需要,使有限资源倾向于收益和成本较高的特性(陈莹婷和许振柱,2014)。此前有关环境因子对植物叶片功能性状影响的研究主要集中在群落尺度上(Petter et al., 2015; Weng et al., 2017),而忽略了种内性状变异(Petter et al., 2015)。郭卫红等(2017)研究表明,物种间各功能性状存在显著差异,同种植物的功能性状变异同样不可忽视。
观光木(Michelia odora),为木兰科的单属种,是中国特有的古老孑遗树种,零星分布于云南、广西、广东等省(区)海拔500~1 000 m常绿阔叶林中,适宜生长于温暖湿润气候、深厚肥沃土壤。随着森林破坏及乱砍乱伐,观光木野外种群数量减少,已被列入国家珍稀濒危二级保护植物。有关观光木的研究主要集中在育苗栽培(欧斌等,2017)、净光合速率(陈凯等,2019)等方面,但有关观光木功能性状方面研究目前还未见相关文献报道。为此,本研究以广西区内5个不同纬度自然生长的成年观光木为研究对象,分析其一年生小枝生物量分配与功能性状的纬度变异规律,以期解决以下几个问题:(1)不同纬度下,观光木的小枝生物量具有何种分配模式?(2)不同纬度间叶片主要功能性状是否存在差异且存在一定的规律?(3)叶片主要功能性状之间存在何种关系、主要功能性状受何种环境因子影响?本研究结果有助于理解观光木对环境的适应特征,其对纬度变异的响应机制等具有重要意义。
1 研究地区与研究方法
1.1 样品采集区概况和地理环境因子数据的获取
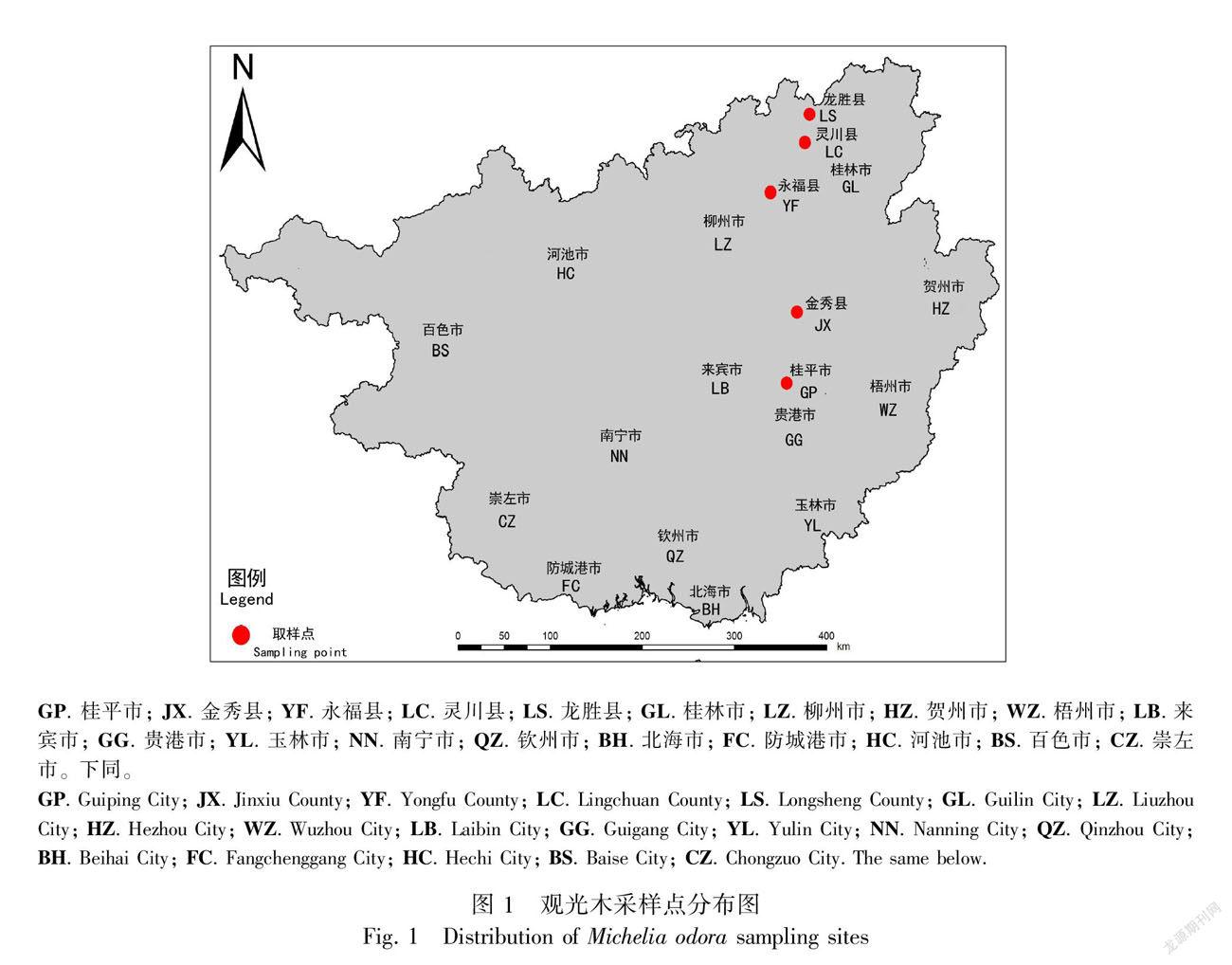
选取广西区内5个不同纬度梯度的自然生长的观光木进行研究(图1),其地理分布范围(109.99°—110.21° E、23.52°—25.89° N)从南向北依次是桂平市(GP)、金秀县(JX)、永福县(YF)、灵川县(LC)、龙胜县(LS),该区域属于亚热带季风气候,夏季炎热多雨,冬季温暖少雨,由南向北呈现明显的温度梯度和降水梯度变化,年均温为17.6~21.4 ℃,最冷月1月份平均气温7.9~13 ℃,最热月7月份平均气温24.3~29.5 ℃。年平均降水量一般在1 534.7~1 992.6 mm之间。全年平均无霜期为345 d,各个采样地的基本环境概况见表1,气象数据来源于国家气象局气象数据中心(http: //data. cma.cn/)。在样品采集过程中,利用手持 GPS测定各樣地所处的经度、纬度,用坡度仪测量坡度,同时用冠层分析仪测量郁闭度。
1.2 样品采集和处理
2019年7月,于各天然分布点,分别选取3株长势良好的成年观光木,在每株标准木的东、南、西、北4个方向,截取冠层全光照下具有完好成熟叶片的 4根当年生小枝。小枝样品放入密封袋并编号,立即放入有冰袋的保温箱中,在黑暗的环境中储藏12 h,将样本带回室内实验室,对叶片、小枝进行分离。从密封袋中随机选取健康完整的11片完全展开、大小相似、未受损的健康叶片,并对叶片进行标记编号,其中10片用于测量叶性状指标,1片用来测气孔性状。
1.3 叶片功能性状测定
用CanoScan LiDE 300扫描仪结合Photoshop测定其叶面积(leaf area,LA)。用精度为0.01 mm的电子游标卡尺在叶片非叶脉的前、中、末端分别测量叶片厚度(leaf thickness,LT),取平均值作为叶厚度。用电子天平称量叶鲜量(leaf fresh weigh,LFW),精确到0.01 g,将取出的叶片放入70 ℃烘箱内烘干至恒重后,称量干重(leaf dry weight, LDW)。同时,将枝条样品烘干至恒重,得出总叶片干重(total leaf dry mass, TLDW)、(小枝上全部叶片和叶柄的总干重)、小枝干重(twig dry weight, TDW)。
植物叶片性状包括比叶面积(specific leaf area, SLA)、叶干物质含量(leaf dry matter content, LDMC)、叶含水率(leaf water content, LWC)和叶组织密度(leaf tissue density, LTD)。
气孔属性性状包括气孔长度(stomatal length, SL)、气孔宽度(stomatal width, SW)、气孔面积(stomatal area, SA)、气孔密度(stomatal density, SD)。测定具体步骤:将无色指甲油均匀涂抹于叶片背面(约1 cm2,注意避开叶脉),待其风干后用透明胶带粘住指甲油涂抹部分并取下铺展在载玻片上制片,采用光学显微镜在40倍物镜和10倍目镜下观察并拍摄图片,每一样品随机选择3个视野记录气孔数目,根据视野内的气孔数量计算SD。同时在每个视野图片中随机选择3个气孔,利用ImageJ 1.8软件分析测定气孔长度、气孔宽度,本文以气孔器保卫细胞的长轴长度(μm)和短轴长度(μm)代表气孔长度和宽度、气孔面积代表气孔大小。各性状的计算公式如下:
比叶面积(SLA, m2·kg-1)=叶面积/叶干重;叶干物质含量(LDMC, g·kg-1)=叶干质量/叶饱和鲜重;叶含水率(LWC, %)=(叶鲜重-叶干重)/叶鲜重×100;叶组织密度(LTD, mg·mm-3)=叶干重/(叶面积×叶厚度)。
1.4 数据处理
采用R 3.5.1进行数据统计分析,利用单因素方差分析(one-way ANOVA)和LSD法进行多重比较(α=0.05),分析观光木5个天然分布点的各性状差异显著性以及不同纬度梯度上功能性状的差异性。用 Pearson相关性分析法对各功能性状间关系进行相关分析,同时使用R 3.5.1软件的“vegan”包进行典型相关分析(CCA),分析功能性状与环境因子的关系。采用异速生长方程对当年生小枝和枝条上总叶片生物量分配进行描述。表达式y=bxa,对数转化为lgy=lgb+algx。式中:x与y表示2个特征参数,利用lgx和lgy作图;b为异速生长常数;a为异速生长指数,即线性关系的斜率(当a=1时为等速生长,a≠1时为异速生长)。使用R 3.5.1的“smatr”包进行标准化主轴回归分析(standardized major axis,SMA)(Warton et al., 2006)。分析纬度变化过程中小枝功能性状间的异速生长关系,通过观光木当年生小枝和小枝总叶片生物量特征值即小枝和总叶片的干重,计算出异速指数, 并通过Warton(Warton & Weber, 2002)的方法对斜率进行异质性检验。采用R 3.5.1和SigmaPlot 10.0绘制图像。文中图表数据为平均值 ± 标准差。
2 结果与分析
2.1 小枝功能特征与异速生长关系
各研究地区中,GP、JX、YF、LC、LS观光木当年生小枝干重和枝条上的总叶片干重如表2所示。
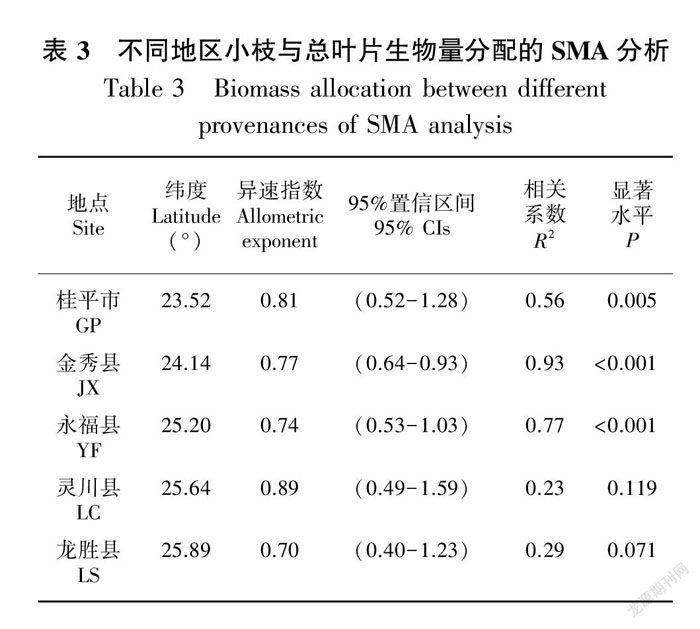
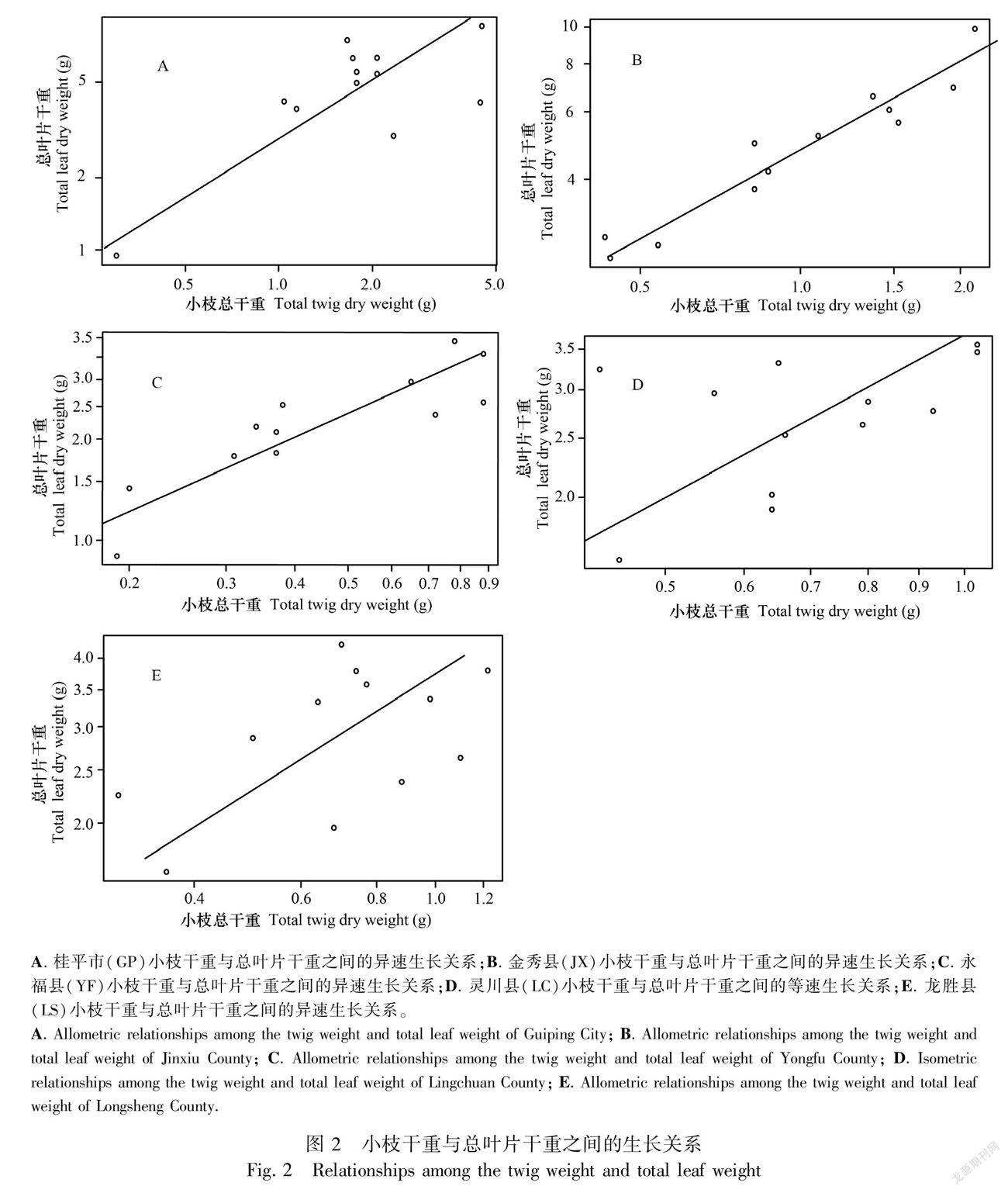
从南向北在不同纬度上,观光木小枝与总叶片生物量分配的回归参数如表3所示,GP小枝干重与总叶干重的异速生长指数显著小于1.0(P=0.005)(图2:A); JX小枝干重与总叶干重的异速生长指数显著小于1.0(P<0.001)(图 2:B);YF小枝干重与总叶干重的异速生长指数显著小于1.0(P<0.001)(图2:C);LS小枝干重与总叶干重的异速生长指数显著小于1.0(P=0.071)(图2:E),4个研究地区的小枝干重与总叶干重均呈异速生长关系,即小枝干重的增加速率大于总叶干重。而LC小枝干重与总叶干重的异速生长指数与1.0无显著差异(P=0.119)(图2:D),呈等速生长关系。
2.2 不同纬度地区观光木功能性状差异比较
方差分析表明,不同纬度地区观光木的叶重、叶面积、比叶面积、叶含水率、气孔大小等呈显著差异,气孔密度差异不显著。由图(3)可知,从南向北随着纬度的上升:叶面积、叶鲜重、叶干重呈逐渐降低的趋势;叶厚度、叶干物质含量、气孔长、气孔大小、气孔密度呈逐渐上升的趋势;比叶面积、叶含水率、气孔宽先增大后减小,总体呈逐渐减低的趋势;叶组织密度先减小后增大。
2.3 观光木各功能性状间的相关性
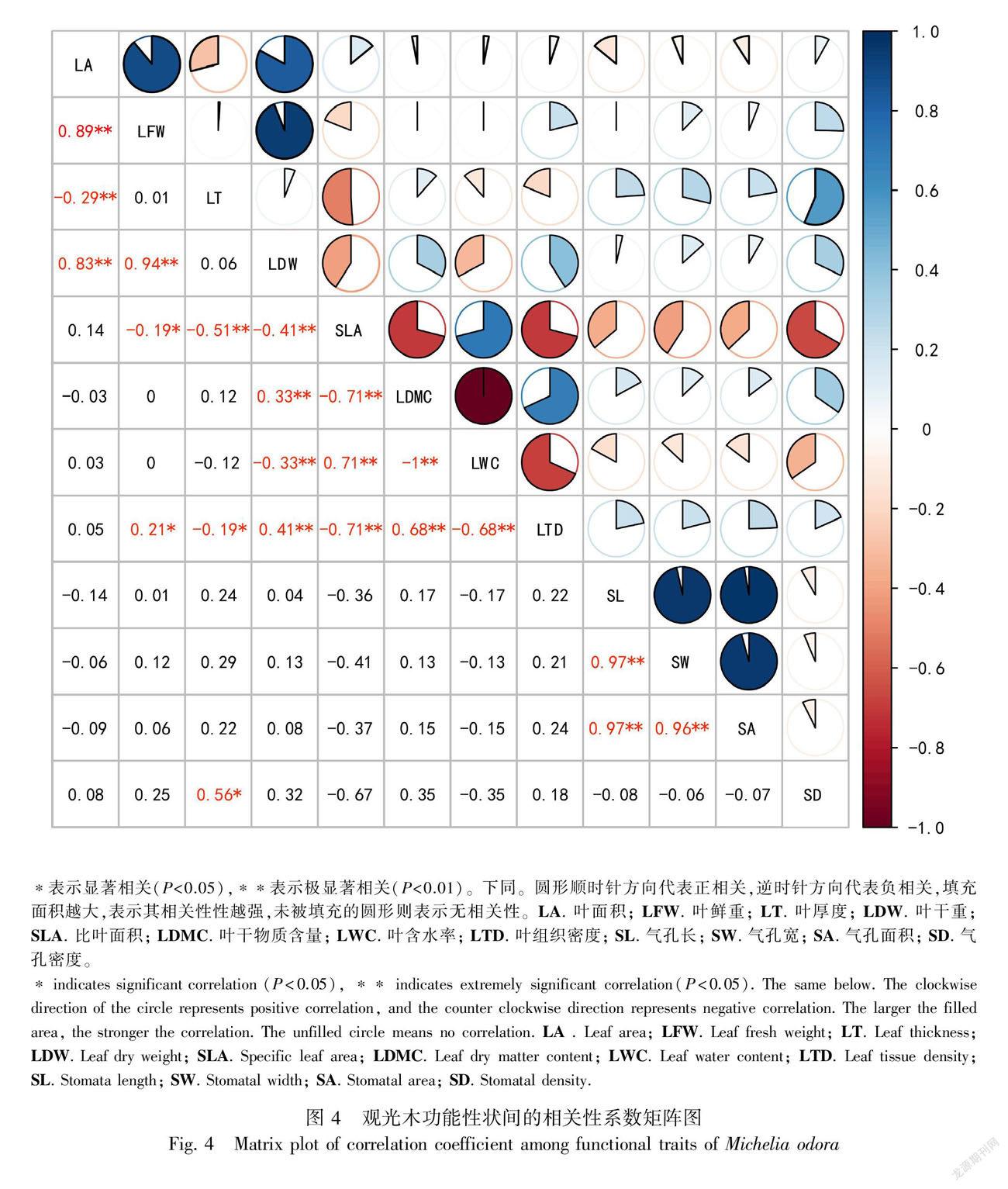
對不同纬度地区的观光木的功能性状进行相关分析结果表明,叶面积与叶鲜重、叶干重呈显著正相关,与叶厚度呈显著负相关;比叶面积与叶含水率呈显著正相关,与叶干物质含量、叶组织密度、气孔密度、叶厚度呈显著负相关;气孔大小与气孔长度、气孔宽度呈显著正相关;叶干物质含量与叶含水率呈极显著负相关,达到极值。
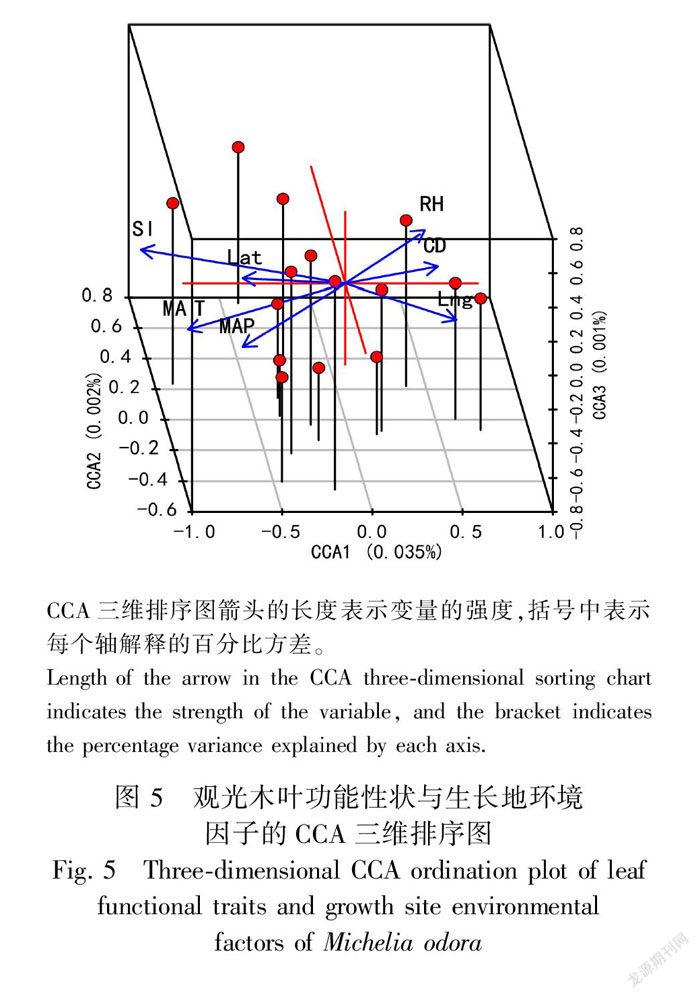
2.4 观光木叶功能性状与环境因子的关系
对CCA结果进行蒙特卡罗检验,其显著性为0.001,说明排序的结果可以接受环境因子对植物功能性状的解释量,但只是7个环境因子综合分析的结果,并不代表每个环境因子都对功能性状具有显著影响,因此需要对环境变量进一步预选,经计算对观光木功能性状具有显著影响的是坡度和年均温(表4)。
CCA的三维排序图显示(图5),坡度是影响观光木功能性状的最显著变量;年平均温度对观光木功能性状也具有显著影响;年均降水量和相对湿度对植物功能性状具有一定强度的影响,但不起显著作用;经度和纬度对植物功能性状影响较小。
3 讨论与结论
3.1 小枝生物量分配策略
不同纬度地区中,观光木当年生小枝重与总叶重除LC地区呈等速生长关系外,其余研究地区均呈异速生长关系, 且随着纬度上升异速指数逐渐减小,即在研究区内随纬度上升,生物量分配更多趋于小枝构建,本研究结果与Sun等(2006)研究植物各器官(叶、茎、根)年增加速率保持恒定的规律不一致。这可能是观光木分布地接近河岸所致,使其可获得充足的水分和养分,叶片蒸腾散失的水分能得到迅速补充,生物量分配更加倾向小枝构建,河岸附近长年受到风力以及人为等因素的影响,小枝需要更多生物量的投资以应对大叶片受到风力、重力、人为因素等的影响
(Niinemets et al., 2007)。而在LC地区观光木当年生小枝干重与总叶片干重呈等速生长关系,与孙蒙柯等(2018)研究一致。由于LC地区观光木处饮用水源保护区,其自然环境相对稳定且能得到较好的保护,该地区可能是更适宜观光木生长的生境。因此,当小枝生物量在有限的条件下,植物更倾向选择构建小枝以达到最优经济分配效率 (Moles & Westoby, 2000)。在不同生境中,观光木通过增大对叶和小枝的投资,体现植物为避开遮荫的限制,对有限资源的权衡以满足植物生长(卢艺苗等,2019),有利于构建群落的稳定性和植物的共存性。
3.2 不同纬度地区的主要叶功能性状的表现特征
观光木种群分布区域广泛,自然生长地的立地与气候环境差异必然会导致种群在生理生态特性方面的分化,使叶片性状存在显著差异(Roscher et al., 2018)。经过长期对当地生境的适应,处于不同生境的同种物种会形成不同的地理种源(安海龙等,2015)。观光木功能性状表现相对不稳定,可能是由于不同纬度地区水热条件差异所致,使其生物量在小枝的分配权衡和资源利用效率也存在差异,导致最能直观表现叶片形态结构的叶面积和叶重发生较大变化。不同研究地区观光木叶面积、叶重、比叶面积、叶干物质含量和气孔大小等均呈显著差异,这可能是其对自然生长地环境影响较为敏感,导致小枝与叶片生物量的分配不均,也表明观光木具有较强的叶片形态可塑性以应对地理环境变化(陈嘉静等,2019)。气孔属性主要与水分利用效率和蒸腾作用以及光合速率有关(Franks et al., 2017),不同环境因子对气孔属性的影响不同,如光照、干旱胁迫下使植物叶片气孔属性发生变化(Yoo et al., 2013)。本研究发现:气孔密度差异不显著,原因可能是研究地水源、光源充足,气孔密度对外界因子变化的响应相同;从南向北随着纬度的上升,由于水热条件的降低使植物变现倾向小枝生物量投资的偏好,导致叶片的生物量逐渐减少,最终使叶面积、叶鲜重、叶干重呈逐渐降低的趋势,与龙嘉翼等(2018)研究一致;叶厚度、叶干物质含量、气孔长、气孔大小、气孔密度呈逐渐上升的趋势,与倪隆康等(2019)研究一致,反映植物面对环境胁迫,通过增强对水分和养分的保存能力和增强光合速率和水分利用效率以应对叶片生物量丧失。
3.3 主要叶功能性状间关系
本研究中,比叶面积与叶组织密度和叶干物质含量存在极显著的负相关,与前人研究一致(郑颖等,2014;郭茹等,2015;戚德辉等,2015),表现出比叶面积-叶组织密度和比叶面积-叶干物质含量间的权衡关系,反映观光木通过对叶片形态结构改变以适应不同纬度环境。叶含水率与比叶面积呈显著正相关,与叶干物质含量呈显著负相关,与段媛媛等(2017)认为,叶含水率与比叶面积呈显著正相关,与叶干物质含量呈显著负相关一致,表明生长在水源和养分充足地区的观光木的叶含水量能保持在相对稳定水平且养分能及时更替,使叶片有较高的资源利用效率。叶厚度与比叶面积呈显著负相关,与Roa-Fuentes等(2015)研究一致,这种以减少叶片厚度碳构建而增加叶片面积碳投资的策略,可以增加植物对水分、养分和光等资源的利用率,可提高光合作用能力(郑芬等,2020)。气孔密度与叶片厚度呈显著正相关,与比叶面积呈显著负相关,说明叶片厚度的增大时使叶片细胞体积相应变化,植物需要相应的使气孔密度的增加来权衡这一变化,且能更高效地利用资源,提高光合速率最终使比叶面积减小,与前人研究的较高气孔密度往往代表较高气孔导度和光合速率一致(Drake et al., 2013),这种权衡关系调控了植物气孔属性对生长环境长期的适应性进化(Franks et al., 2009)。
3.4 功能性状与研究地环境因子的关系
CCA分析表明,观光木叶功能性状与自然生长地的坡度、年均温具有显著相关性,可以认为观光木叶功能性状主要受年均温和坡度的影响。本研究5个自然生长地中,从南向北呈明显的纬度梯度分布,观光木叶片功能性状呈现出一定的纬度格局,这与孟婷婷等(2007)研究结果一致,水热变化能直接影响植物的生理生态特征。
坡度对观光木功能性状的影响最为显著,坡度的差异影响微生境的空间变异和其对植物功能性状的筛选,刘旻霞和马建祖(2013)研究表明,坡度越大环境条件越恶劣,土壤含水量低、土壤矿化严重,而坡度较低时有相对高的土壤营养资源和土壤含水量。坡度大的地区积温少,同时坡度影响坡面径流冲刷强度,坡度越大,土壤水分和养分流失量越多而含量越低;坡度和坡向的组合决定植物能够接受的光照强度和面积。总体而言,5个自然生长地由于坡度的差异,使观光木对其营养元素和有机质含量吸收效率不同,从而影响植物的功能性状,当坡度较大时,植物倾向于长出较厚的叶子来抵御由于冲蚀引起的水分和养分流失(丁佳等,2011),同时调节比叶面积和叶干物质含量的关系,从而提高植物体内养分保存能力。年均溫与植物叶功能性状呈显著相关性,与Kamiyama等(2014)研究一致。这可能是观光木小枝在水热条件好的条件下生物量能更多向叶片分配,且水热条件充足时植物叶片水分能保持相对稳定且养分能及时更替,使叶片有较高的资源利用效率;也表明观光木可以通过改变叶片形态和生理特征来适应高温环境、降低蒸腾强度防止体内水分散失,维持正常的生理代谢活动,与孟婷婷等(2007)、孙坤等(2014)研究一致。高纬度年均温较低地区植物叶片需要有更高的水分、气孔密度以及更好组织结构以应对环境胁迫和防止叶片水分散失;气温的变化是影响植物生长的重要因子(Costa et al., 2017),低温限制光合作用所需要的碳吸收以及重要元素在植物体内的流动,导致比叶面积、叶重变小;气孔属性与年均温呈显著相关性,可以推断当温度升高时,植物蒸腾作用加快,植物为了减少水分流失,使气孔关闭,是对胁迫环境的一种适应策略。这进一步说明观光木的生长、生存不仅取决于自身的生理特性,且与生长地地理环境因子间关系密切。年平均温度的显著贡献表明气候的重要性,从更大意义上描述了植物对温度变化的脆弱性,这可能在气候变化和全球气温上升的情况下产生的负面影响,这也证明了气候异质性和植物功能性状之间关键联系的正确性。
叶柄是叶片与枝条相互连接的关键部位,与小枝和叶片的生物量分配都有密切联系,探究生物量分配策略时叶柄应作为单独研究指标,本研究未将叶柄单独分开,结果可能会产生偏差。本文由于选取的5个研究地近似分布于同一经线上,且研究地海拔梯度差异不大,所以仅选取纬度以及与温度和水分相关的环境因子进行研究。
参考文献:
CHEN JJ, YU H, XU CB, et al., 2019. Effects of provenance and common garden environment on leaf functional traits of Machilus pauhoi seedlings[J]. Chin J Appl Environ Biol, 25(3): 648-654. [陈嘉静, 余华, 徐朝斌, 等, 2019. 种源与同质园环境对刨花楠幼苗叶功能性状的影响[J]. 应用与环境生物学报, 25(3): 648-654.]
CHEN K, YANG M, LIU SN, 2019. Effects of different lights on growth and photosynthetic physiological characteristics of T. odorum seedlings[J]. J Beihua Univ(Nat Sci Ed), 20(4): 537-541. [陈凯, 杨梅, 刘世男, 2019. 不同光照对观光木幼苗生长及光合生理特性的影响[J]. 北华大学学报(自然科学版), 20(4): 536-541.]
CHEN YT, XU ZZ, 2014. Review on research of leaf economics spectrum[J]. Chin J Plant Ecol, 38(10): 1135-1153. [陈莹婷, 许振柱, 2014. 植物叶经济谱的研究进展[J]. 植物生态学报, 38(10): 1135-1153.]
COLLINS CG, WRIGHT SJ, WURZBURGER N, 2016. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees[J]. Oecologia, 180(4): 1037-1047.
COSTA DS, CLASSEN A, FERGER S, et al., 2017. Relationships between abiotic environment, plant functional traits, and animal body size at Mount Kilimanjaro, Tanzania[J]. PLoS ONE, 12(3): e0174157.
DAI L, GUO X, KE X, et al., 2020. Biomass allocation and productivity-richness relationship across four grassland types at the Qinghai Plateau[J]. Ecol Evol, 10(1): 506-516.
DING J, WU Q, YAN H, et al., 2011. Effects of topographic variations and soil characteristics on plant functional traits in a subtropical evergreen broad-leaved forest[J]. Biodivers Sci, 19(2): 158-167. [丁佳, 吴茜, 闫慧, 等, 2011. 地形和土壤特性对亚热带常绿阔叶林内植物功能性状的影响[J]. 生物多样性, 19(2): 158-167.]
DRAKE PL, FROEND RH, FRANKS PJ, 2013. Smaller, faster stomata: scaling of stomatal size, rate of response, and stomatal conductance[J]. J Exp Bot, 64(2): 495-505.
DUAN YY, SONG LJ, NIU SQ, et al., 2017. Variation in leaf functional traits of different-aged Robinia pseudoacacia communities and relationships with soil nutrients[J]. Chin J Appl Ecol, 28(1): 28-36. [段媛媛, 宋丽娟, 牛素旗, 等, 2017. 不同林龄刺槐叶功能性状差异及其与土壤养分的关系[J]. 应用生态学报, 28(1): 28-36.]
FRANKS PJ, BERRY JA, LOMBARDOZZI DL,et al., 2017. Stomatal function across temporal and spatial scales: deep-time trends, land-atmosphere coupling and global models[J]. Plant Physiol, 174(2): 583-602.
FRANKS PJ, DRAKE PL, BEERLING DJ, 2009. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: an analysis using Eucalyptus globulus[J]. Plant Cell Environ, 32(12): 1737-1748.
GUITTAR J, GOLDBERG D, KLANDERUD K, et al., 2016. Can trait patterns along gradients predict plant community responses to climate change?[J].Ecology, 97(10): 2791-2801.
GUO R, WEN ZM, WANG HX, et al., 2015. Relationships among leaf traits and their expression in different vegetation zones in Yanhe River basin, Northwest China[J]. Chin J Appl Ecol, 26(12): 3627-3633. [郭茹, 温仲明, 王红霞, 等, 2015. 延河流域植物葉性状间关系及其在不同植被带的表达[J]. 应用生态学报, 26(12): 3627-3633.]
GUO WH, WANG H, YU MK, et al., 2017. Latitude variation mechanism of leaf traits of Metasequoia glyptostroboides in eastern coastal China[J]. Chin J Appl Ecol, 28(3): 772-778. [郭卫红, 王华, 虞木奎, 等, 2017. 沿海地区水杉叶片性状的纬度变化机制[J]. 应用生态学报, 28(3): 772-778.]
HE NP, LIU CC, ZHANG JH, et al., 2018a. Perspectives and challenges in plant traits: From organs to communities[J]. Acta Ecol Sin, 38(19): 6787-6796. [何念鹏, 刘聪聪, 张佳慧, 等, 2018a. 植物性状研究的机遇与挑战: 从器官到群落[J]. 生态学报, 38(19): 6787-6796.]
KAMIYAMA C, KATABUCHI M, SASAKI T,et al., 2014. Leaf-trait responses to environmental gradients in moorland communities: contribution of intraspecific variation, species replacement and functional group replacement[J]. Ecol Res, 29(4): 607-617.
LIU MX, MA JZ, 2013. Feature variations of plant functional traits and environmental factor in south-and north-facing slope[J]. Res Soil Water Conserv, 20(1): 102-106. [刘旻霞, 马建祖, 2013. 阴阳坡植物功能性状与环境因子的变化特征[J]. 水土保持研究, 20(1): 102-106.]
LONG JY, ZHAO YM, KONG XQ, et al., 2018. Trade-offs between twig and leaf traits of ornamental shrubs grown in shade[J]. Acta Ecol Sin, 38(22): 8022-8030. [龙嘉翼, 赵宇萌, 孔祥琦, 等, 2018. 观赏灌木小枝和叶性状在林下庇荫环境中的权衡关系[J]. 生态学报, 38(22): 8022-8030.]
LU YM, WANG MT, CHEN XP, et al., 2019. Effects of the current-year shoot stem configuration on leaf biomass in different canopy heights of woody plants in evergreen broad-leaved forest in Jiangxi Province, China[J]. Chin J Appl Ecol, 30(11): 3653-3661. [卢艺苗, 王满堂, 陈晓萍, 等, 2019. 江西常绿阔叶林木本植物不同冠层高度当年生小枝茎构型对叶生物量的影响[J]. 应用生态学报, 30(11): 3653-3661.]
MENG TT, NI J, WANG GH, 2007. Plant functional traits, environments and ecosystem functioning[J]. Chin J Plant Ecol, 31(1): 150-165. [孟婷婷, 倪健, 王国宏, 2007. 植物功能性状与环境和生态系统功能[J]. 植物生态学报, 31(1): 150-165.]
MOLES AT, WESTOBY M, 2000. Do small leaves expand faster than large leaves, and do shorter expansion times reduce herbivore damage[J]. Oikos, 90(3): 517-524.
NI LK, GU DX, HE W, et al., 2019. Research advances in plant ecological adaptability in karst area[J]. Chin J Ecol, 38(7): 2210-2217. [倪隆康, 顾大形, 何文, 等, 2019. 岩溶区植物生态适应性研究进展[J]. 生态学杂志, 38(7): 2210-2217.]
NIINEMETS, PORTSMUTH A, TENA D, et al., 2007. Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy[J]. Ann Bot, 100: 283-303.
NIU SL, CLASSEN AT, LUO YQ, 2018. Functional traits along a transect[J]. Funct Ecol, 32(1): 4-9.
OU B, PENG L, LIAO CX, et al., 2017. Study on cultivation techniques and quality grading indexes for seedlings of Tsoongiodendron odorum[J]. S Chin For Sci, 45(6): 30-32. [歐斌, 彭丽, 廖彩霞, 等, 2017. 观光木实生苗培育技术及苗木质量分级指标研究[J]. 南方林业科学, 45(6): 29-32.]
PETTER G, WAGNER K, WANEK W,et al., 2015. Functional leaf traits of vascular epiphytes: vertical trends within the forest, intra- and interspecific trait variability, and taxonomic signals[J]. Funct Ecol, 30(2): 188-198.
QI DH, WEN ZM, YANG SS, et al., 2015. Trait-based responses and adaptation of Artemisia sacrorum to environmental changes[J]. Chin J Appl Ecol, 26(7): 1921-1927. [戚德辉, 温仲明, 杨士梭, 等, 2015. 基于功能性状的铁杆蒿对环境变化的响应与适应[J]. 应用生态学报, 26(7): 1921-1927.]
ROA-FUENTES LL, TEMPLER PH, CAMPO J, 2015. Effects of precipitation regime and soil nitrogen onleaf traits in seasonally dry tropical forests of the Yucatan Peninsula, Mexico[J]. Oecologia, 179(2): 585-597.
ROSCHER C, SCHUMACHER J, LIPOWSKY A, et al., 2018. Functional groups differ in trait means, but not in trait plasticity to species richness in local grassland communities[J]. Ecology, 99(10): 2295-2307.
SUN K, JIANG BY, ZHANG SH, et al., 2014. Study on the response of leaf size traits in Hippophae tibetana Schlecht with precipitation and temperature[J]. J NW Norm Univ (Nat Sci Ed) , 50(6): 71-76. [孫坤, 蒋碧玉, 张世虎, 等, 2014. 西藏沙棘叶片性状对水热条件的响应研究[J]. 西北师范大学学报(自然科学版), 50(6): 71-76.]
SUN MK, CHENG L, WANG MT, et al., 2018. Twig biomass allocation of woody species in evergreen broad-leaf forest, Wuyi Mountain[J]. Chin J Ecol, 37(6): 1815-1823. [孙蒙柯, 程林, 王满堂, 等, 2018. 武夷山常绿阔叶林木本植物小枝生物量分配[J]. 生态学杂志, 37(6): 1815-1823.]
WARTON DI, WEBER NC, 2002. Common slope tests forbivariate errors-in-variables models[J]. Biom J, 44(2): 161-174.
WARTON DI, WRIGHT IJ, FALSTER DS,et al., 2006. Bivariate line-fitting methods for allometry[J]. Biol Rev Camb Philos Soc, 81(2): 259-291.
WENG E, FARRIOR CE, DYBZINSKI R,et al., 2017. Predicting vegetation type through physiological and environmental interactions with leaf traits: evergreen and deciduous forests in an earth system modeling framework[J]. Glob Change Biol, 23: 2482-2498.
YANG DM, MAO LC, PENG GQ, 2011. Within-twig biomass allocation in evergreen and deciduous broad-leaved species: Allometric scaling analyses[J]. Bull Bot Res, 31(4): 472-477. [杨冬梅, 毛林灿, 彭国全, 2011. 常绿和落叶阔叶木本植物小枝内生物量分配关系研究: 异速生长分析[J]. 植物研究, 31(4): 472-477.]
YOO CY, PENCE HE, JIN JB, et al., 2010. The Arabidopsis GTL1 transcription factor re-gulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1[J]. Plant Cell, 22(12): 4128-4141.
ZHENG F, LI ZJ, QIU ZJ, et al., 2020. Effects of understory light on functional traits of evergreen broad-leaved forest saplings in Nanling Mountains, Guangdong Province[J]. Acta Ecol Sin, 40(13): 4516-4527. [郑芬, 李兆佳, 邱治军, 等, 2020. 广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响[J]. 生态学报, 40(13): 4516-4527.]
ZHENG Y, WEN ZM, SONG G, et al., 2014. The influence of environment and phylogenic background on variation in leaf and fine root traits in the Yanhe River catchment, Shaanxi, China[J]. Acta Ecol Sin, 34(10): 2682-2692. [郑颖, 温仲明, 宋光, 等, 2014. 环境及遗传背景对延河流域植物叶片和细根功能性状变异的影响[J]. 生态学报, 34(10): 2682-2692.]
ZIRBEL CR, BASSETT T, GRMAN E, et al., 2017. Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration[J]. J Appl Ecol, 54 (4) :1070-1079.
(责任编辑 李 莉)
收稿日期:2021-02-03
基金项目:国家自然科学基金(31200330); 南宁树木园优质用材树种科研项目培育与深化项目 [Supported by National Natural Science Foundation of China (31200330); Cultivation and Deepening Project of Nanning Arboretum High-Quality Timber Tree Species Research Project]。
第一作者: 蒙检(1995-),硕士研究生,主要从事森林生态学研究,(E-mail)812107833@qq.com。
通信作者:罗应华,博士,副教授,主要从事生物多样性研究,(E-mail)liliaceaeluo@163.com。
猜你喜欢
数学学习与研究(2018年3期)2018-03-14
齐鲁周刊(2017年29期)2017-08-08
中学政史地·教学指导版(2016年12期)2017-05-16
中国市场(2016年36期)2016-10-19
天津农业科学(2015年12期)2015-12-03
河北渔业(2015年10期)2015-10-15
中学政史地·教学指导版(2009年9期)2009-05-19
