
藏南布丹拉山南坡种子植物区系海拔格局分析
2022-04-08 01:08王俊伟明升平杨坤何敏拉琼
广西植物 2022年3期
关键词:植物多样性
王俊伟 明升平 杨坤 何敏 拉琼

摘 要:生態群落交错区通常因物种丰富、区系成分复杂而被视为关键带。藏南布丹拉山处在半湿润向半干旱的生态环境过渡带上,因其特殊的自然地理环境而有着丰富的山地植物多样性,但这一重要生态过渡区的种子植物组成和区系成分海拔分布格局目前尚缺乏了解。为了理清布丹拉山南坡种子植物区系成分及其垂直分布变化格局,该文通过野外植物群落的样方调查、实验室标本鉴定与相关文献资料查阅,系统地研究了布丹拉山南坡种子植物属种区系地理成分性质,探讨了优势属丰富度和属区系地理成分沿海拔梯度的变化趋势。结果表明:(1)该区种子植物约有45科156属316种,其中单种属和小属最丰富,区系地理成分在属种水平上温带分布型占主导地位。(2)在地理成分垂直分布梯度上,温带分布型属在海拔4 000 m时出现峰值,含53属,之后随海拔的升高而呈下降的趋势。(3)该区植物区系地理成分的性质和物种丰富度的垂直分布变化格局可能主要受当地半湿润的气候条件与局域环境因子和地形因子综合作用的影响。该研究结果可为该区域的植物多样性保护与资源利用研究提供重要的基础资料。
关键词: 植物区系, 海拔梯度, 植物多样性, 植物地理, 布丹拉山
中图分类号:Q948.15
文献标识码:A
文章编号:1000-3142(2022)03-0384-10
Analysis of elevation pattern of seed flora on the south
slope of Budanla Mountain in southern Tibet, China
WANG Junwei1,2, MING Shengping3, YANG Kun1,2, HE Min1,2, LA Qiong1,2*
( 1. College of Sciences, Tibet University, Lhasa 850000, China; 2. Ecological Research Center of Tibet University, Tibet University,
Lhasa 850000, China; 3. Yunnan Lijiang Forest Ecosystem National Observation and Research Station, Kunming Institute
of Botany, Chinese Academy of Sciences, Lijiang 674100, Yunnan, China )
Abstract:The ecotone of ecological communities is usually rich in species and complex in floristic components, which is regarded as a key zone. Budanla Mountain is located in the transition zone of semi-humid to semi-arid ecological environment in Tibet. Because of its special natural geographical environment, it has rich mountain plant diversity. However, the composition and floristic characteristics of seed plants in this important ecological transition area are still not well understood. In order to clarify the floristic composition and vertical distribution pattern of seed plants on the south slope of Budanla Mountain, the floristic characteristics of seed plants on the south slope of this mountain were systematically studied through field plant community quadrat investigation, laboratory specimen identification and related literature review in this paper, the richness pattern of advantage genera and geographical composition along the altitude gradient was also explored. The results were as follows: (1) There were approximately 316 species of seed plants belonging to 45 families and 156 genera in the south slope of Budanla Mountain, in which there were the most abundant single species and small genera of seed plants, the temperate distribution type was dominant at the genus and species level. (2) In the vertical distribution gradient of geographical elements, the temperate distribution type had a peak value at 4 000 m, including 53 genera, and showed a downward trend with the increase of altitude. (3) The nature of floristic geographical elements and the vertical distribution of species richness in this area might be mainly affected by the local semi-humid climate, local environmental factors and topography. The results of this study can provide important basic data for the study of plant diversity protection and resource utilization in this region.
Key words: flora, altitude gradient, plant diversity, phytogeography, Budanla Mountain
山地生态系统是研究生物和非生物因子在海拔梯度上变化规律的理想天然实验室,它能在有限的地理空间范围内集地形、海拔和温度等多种环境因子和地形因子于一体而生境呈高度异质性(Devadoss et al., 2020; Tito et al., 2020),并孕育着丰富的植物多样性(方精云等,2004a;Yu et al., 2021)。山地生态系统成为研究植物多样性的热点区域,受到生物地理学家、进化生物学家和生态学家的广泛关注(Lomolino, 2001)。另外,由于山地生态系统特殊的生境可以给很多冰期植物提供天然避难所以及提供新分类群的分化条件,所以这一地区往往会有一些成分古老和新兴分化的年轻植物类群,植物区系成分新老兼备(吴征镒,1979)。因此,研究山地生态系统的植物多样性,不仅可以了解山地植物区系构成特点及性质,而且可以理清山地植物区系地理成分的海拔梯度分布规律,从而揭示山地植物区系自身演化发展与环境因子的关系(王荷生,1992;周浙昆等,2017)。
雅鲁藏布江中游谷地作为典型的山地生态系统,有着丰富的山地植物多样性(刘敏杰和李华军,2020)。屈兴乐等(2019)对雅鲁藏布江中游河谷地区灌丛草地的区系组成及特征进行研究,结果表明该区灌丛草地群落结构简单,区系地理成分显示温带性质;Li et al.(2021)对雅鲁藏布江中上游沙生植物的群落结构与多样性分布格局进行抽样调查,研究结果显示该区植物群落类型可划分为7种,水热条件是影响植物分布格局的主要因素;拉琼等(2014)对雅鲁藏布江上游源头区至下游河岸物种丰富度格局的研究结果显示,研究区可划分为5种植被类型,物种丰富度格局受海拔和经度控制呈现浅“U”形。可见,该区是山地植物多样性研究的热点地区。生态过渡带通常因其气候独特、物种丰富和区系成分复杂而被视为关键带。藏南布丹拉山体处在半湿润向半干旱的生态环境过渡带上,是印度洋暖湿气流沿雅鲁藏布江向高原面输送的最后一道天然屏障,形成了南北坡迥然不同的气候环境,尤其南坡植物物种异常丰富,但目前有关这一重要生态过渡区的种子植物组成还缺乏了解,鲜见报道有关这一重要区域的山地植物区系特征特别是其植物区系成分沿海拔梯度的垂直分布格局研究。基于此,我们运用生态学、植物地理学和植物群落生态学的研究方法,通过对布丹拉山南坡海拔梯度上种子植物多样性及区系构成特点的研究,分析了布丹拉山南坡种子植物区系沿海拔梯度的分布格局特征及其可能的环境影响因子。拟探究以下问题:(1) 该区域种子植物多样性特征及区系组成性质如何;(2) 该区域种子植物属的区系地理成分和优势属沿海拔梯度的分布变化规律如何,这些变化规律是否一致。本文丰富了雅鲁藏布江中游山地植物多样性及其区系成分的研究,尤其是对物种丰富的生态群落交错区区系成分的研究和区系成分沿海拔梯度变化规律的研究。
1 研究区自然概况
布丹拉山原称布喳拉山,意为“散落的经书”(边吉,2006),位于雅鲁藏布江中游加查峡谷的山区地带,其山顶海拔有5 127 m,山脚河谷区域海拔为3 700 m,海拔落差大。南坡地理位置为92°22′—92°36′ E、29°02′—29°03′ N。行政区划上隶属于西藏自治区山南市加查县拉绥乡拉索自然村,是加查县县界的最西端,加查县以其为行政界线,与曲松县毗邻(陈芳媛,2009)。省道八邱公路(S306)东西横贯全山,布丹拉山南坡山脚发源有拉绥流域,拉绥河沟谷地是加查县成片分布的农耕地之一(加查县地方志编纂委员会,2010)。
布丹拉山地处冈底斯山—念青唐古拉山与喜马拉雅山东部的缝合地带,是喜马拉雅山区与雅鲁藏布江中游谷地区的过渡带,地貌区划上属于雅鲁藏布江中游深切河谷区,形态以高山峡谷为主(祝嵩,2012)。地势高峻,地形复杂多样,海拔高低悬殊,使这一地区垂直自然带差异变化明显,水热再分配的垂直地带性规律明显,气温在垂直方向上的变化明显高于水平方向上的变化,具有典型山地气候环境的特点。印度洋的暖湿气流被高耸的喜马拉雅山阻挡,雅鲁藏布江的存在为暖湿气流提供了一个巨大的水汽大通道,使其在此区域与高原的寒冷空气交汇,从而形成了特殊的高原温带半湿润气候(祝嵩,2012)。
布丹拉山山体顺依加查峡谷,山系属于东西走向,是这一峡谷地区重要山地的组成部分,雅鲁藏布江加查峡谷以东的中下游地区,气候温暖湿润,91.76%的降雨主要集中在5—9月,山地植被茂盛。气温年变化相对较小、昼夜温差变化大,月平均气温最高月为7月(16.4 ℃),最低月为1月(-1.0 ℃)。布丹拉山南坡海拔落差大,植被垂直差异显著,按气候条件从山脚的拉绥河谷起可以划分为不同的小气候类型,分别为拉绥沟谷温暖半湿润气候(3 800 m以下)、山地温和半湿润气候(3 900~4 200 m)、亚高山温凉半湿润气候(4 200~4 800 m)、高山寒冷半湿润气候(4 800 m以上)(加查縣地方志编纂委员会,2010;薛鸿博,2015)。
2 研究方法
2.1 野外调查
2018年9月至2020年7月分多次对布丹拉山南坡的种子植物进行植物群落样方调查、植物标本采集和植物影像资料收集的研究。调查方法采用样方法和线路法相结合的植物调查方法(线路法是后面的补充调查和采集标本)。选择有代表性的能体现自然生境和人为干扰少的植被类型,从海拔3 800~5 127 m、海拔跨度1 327 m的梯度上,每隔50 m设置一个海拔梯度,在每个海拔梯度上随机设置6个5 m × 5 m的平行样方,共计27个海拔梯度,162个样方。每个样方中详细记录物种数、盖度和生态因子(坡度、海拔和经纬度)等信息。为了能代表整个布丹拉山南坡,我们还调查并记录了样方四周的物种,以及海拔样带之间出现的物种。
2.2 数据分析
首先,通过标本鉴定与查阅相关资料确定以Flora of China学名为准的种子植物物种名录;然后,按照《中国植物志》第一卷、吴征镒等(2003,2006)和陈灵芝等(2015)对属分布类型划分的原理和方法,确定属级分布区类型;最后,结合物种的自然分布区对种进行区系分析。
根据调查所获得的物种海拔分布信息,采用Excel 2019软件统计每一海拔段内的物种类群和所属的区系地理成分的数量分布,之后再统计分析优势属和属区系地理成分的海拔梯度变化趋势。
3 结果与分析
3.1 布丹拉山南坡种子植物多样性
布丹拉山南坡种子植物物种组成,经统计共有种子植物45科156属316种(以Flora of China为准)。其中,裸子植物有1科1属2种,分别占总数的2.20%、0.64%和0.63%;被子植物有44科155属314种,分别占总数的97.70%、99.36%和99.37%;双子叶植物38科129属267种,单子叶植物6科26属47种。布丹拉山南坡种子植物的构成主要以被子植物为主,其中以双子叶植物占据优势。
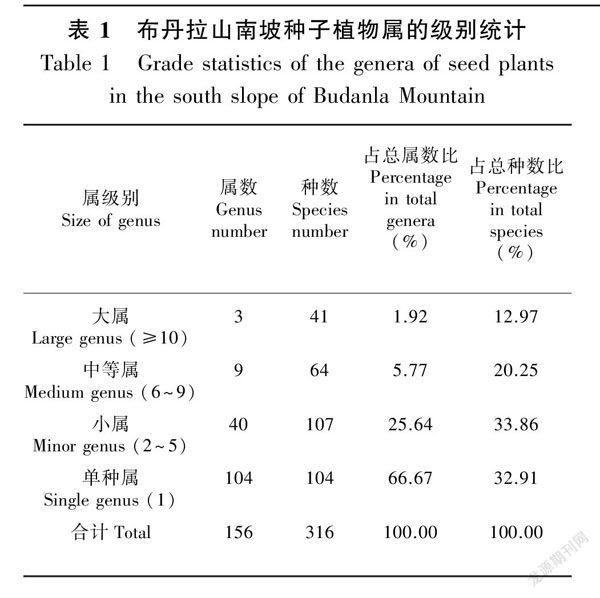
对种子植物属的数量结构分析,按研究区内属所含种的数量将该区的种子植物156个属分为4个等级(张静等,2019),分别为单种属(含1种)、小属(2~5种)、中等属(6~9种)和大属(≥10种)。从表1可以看出,单种属共104属,占总属数的66.67%,所含种数占总种数的32.91%;含2~5种的小属共40属,占总属数的25.64%,所含种数占总种数的33.86%。由此可见,单种属和小属构成了布丹拉山南坡种子植物属的主体,是布丹拉山南坡种子植物属多样性的主要成分。单种属占比大,反映出布丹拉山南坡在属级水平上分化程度较高、多样性丰富。
包含6种以上的属有12个属(表1),分别是虎耳草属(Saxifraga)含18种、马先蒿属(Pedicularis)含12种、龙胆属(Gentiana)含11种、报春花属(Primula)含7种、风毛菊属(Saussurea)含6种、蒿属(Artemisia)含7种、红景天属(Rhodiola)含7种、蓼属(Persicaria)含8种、委陵菜属(Potentilla)含8种、早熟禾属(Poa)含7种、紫堇属(Corydalis)含7种和紫菀属(Aster)含7种。从表1可以看出,布丹拉山南坡种子植物包含10种以上的大属是虎耳草属、马先蒿属和龙胆属,说明布丹拉山南坡草本种子植物属的优势现象非常明显。
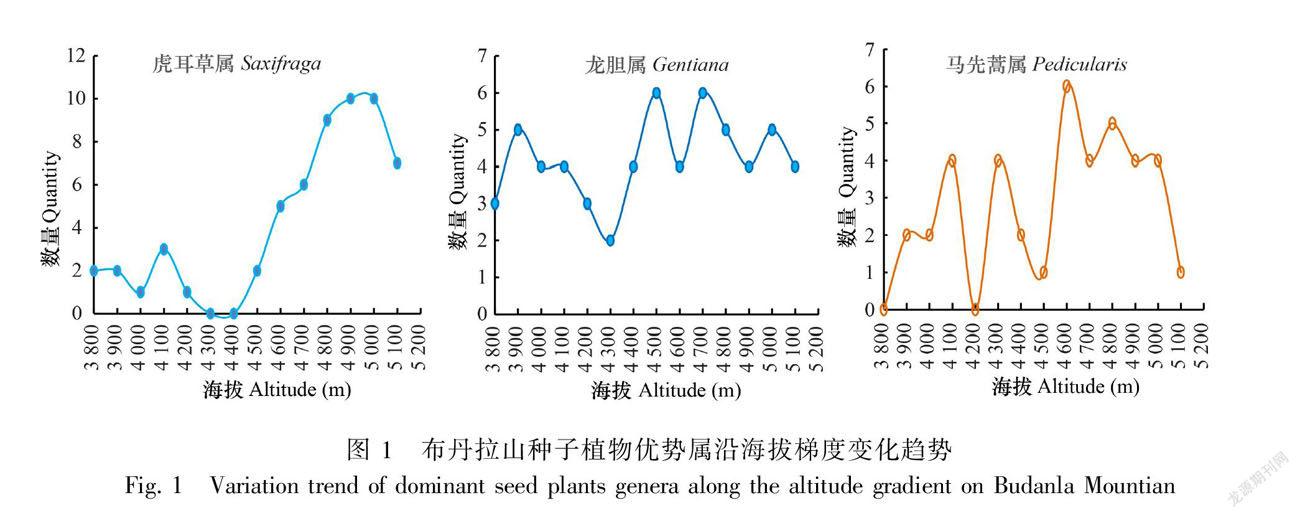
3.2 优势属的垂直海拔分布特征
由图1可知,虎耳草属在海拔4 900 ~5 000 m时物种数分布呈现最高值,含种数10种,而在海拔4 300~4 400 m时没有物种分布,随着海拔的升高呈现先增后减的变化趋势;龙胆属所含物种数在海拔4 300 m时最低(仅含2种),在海拔4 500、4 700 m时出现峰值(含6种);马先蒿属在海拔4 600 m时物种数达到最大值,含种属6种。龙胆属和马先蒿属随着海拔的升高呈现出不规则的变化规律,起伏波动较大,但整体上还是有最高峰值,微弱的有先增后减的变化趋势。
3.3 植物区系地理成分沿海拔梯度的变化
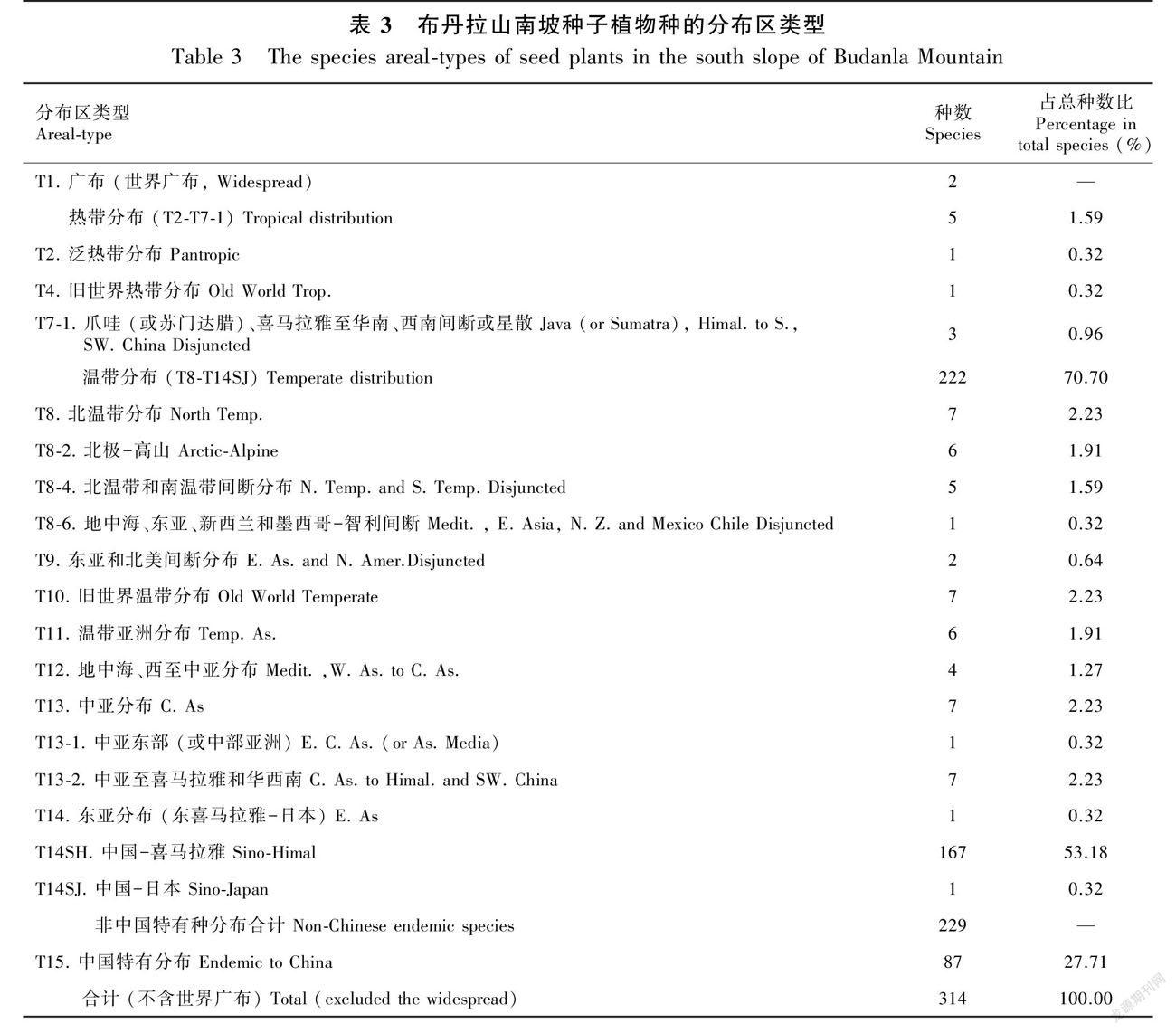
3.3.1种子植物属种地理成分分析 对布丹拉山南坡种子植物区系属的地理成分进行了统计分析,将布丹拉山南坡种子植物区系的156属划为10个分布区类型及10个变型(表2)。其中,世界广布属有23属、热带成分的属有6属(T2-T6型)、温带分布属有123属(T8-T14型)和中国特有分布属有4属,各成分总属数分别占非世界广布总属数的4.51%、92.48%、3.01%。温带分布属包括虎耳草属、马先蒿属和委陵菜属等一些本区优势属,所含属数超过了总属数的2/3,温带分布属中以北温带和南温带间断分布和北温带分布为主,占温带分布总属数的42.28%。由此可见,布丹拉山南坡种子植物的区系在属分布区类型上呈现温带性质,北温带和南温带间断分布和北温带分布型地理成分占绝对优势,符合该区属高原温带半濕润气候的特点,世界广布型和热带分布型其次。由于世界种子植物物种数目庞大,至今没有明确的种分布区类型划分方法,因此本研究参照陈灵芝等(2015)对中国种子植物属的分布区类型划分原则并结合种子植物的自然分布地区,将布丹拉山南坡316种种子植物分布区划分为11个类型及8个变型(表3)。对比属的分布区,种的分布区增加了地中海、西至中亚分布型和中亚分布型,非中国特有种成分有229种,其中温带分布成分222种,占非世界广布种总数的70.70%,占非中国特有种数的96.94%,占非中国特有种成分的绝对主导地位。而温带成分中又以中国-喜马拉雅成分为主,占温带成分总种数的75.23%。热带成分分布的有5种,占比很小,仅占非世界广布种总数的1.59%,世界广布的有2种。布丹拉山南坡316种子植物中,中国特有种成分有87种,占非世界广布种总数的27.71%。从种的地理成分上分析,此区域的种子植物区系也是呈现温带性质,鲜有热带性质的物种出现。
上新世时期( 5.33~2.58 Ma ),雅鲁藏布江南岸喜马拉雅山区中东段的这一地带是暖温带亚热带植被类型(祝嵩,2012),而本研究属种区系地理成分以温带成分占据主导优势,鲜有热带成分渗入,也符合这种植被特征。这种区系地理成分的构成特点及性质与明升平等(2018)在拉萨河谷根培乌孜山的区系研究结果一致,表明在青藏高原面上拉萨河谷和雅鲁藏布江河谷的植物区系有相同的起源及其区系成分交流紧密。
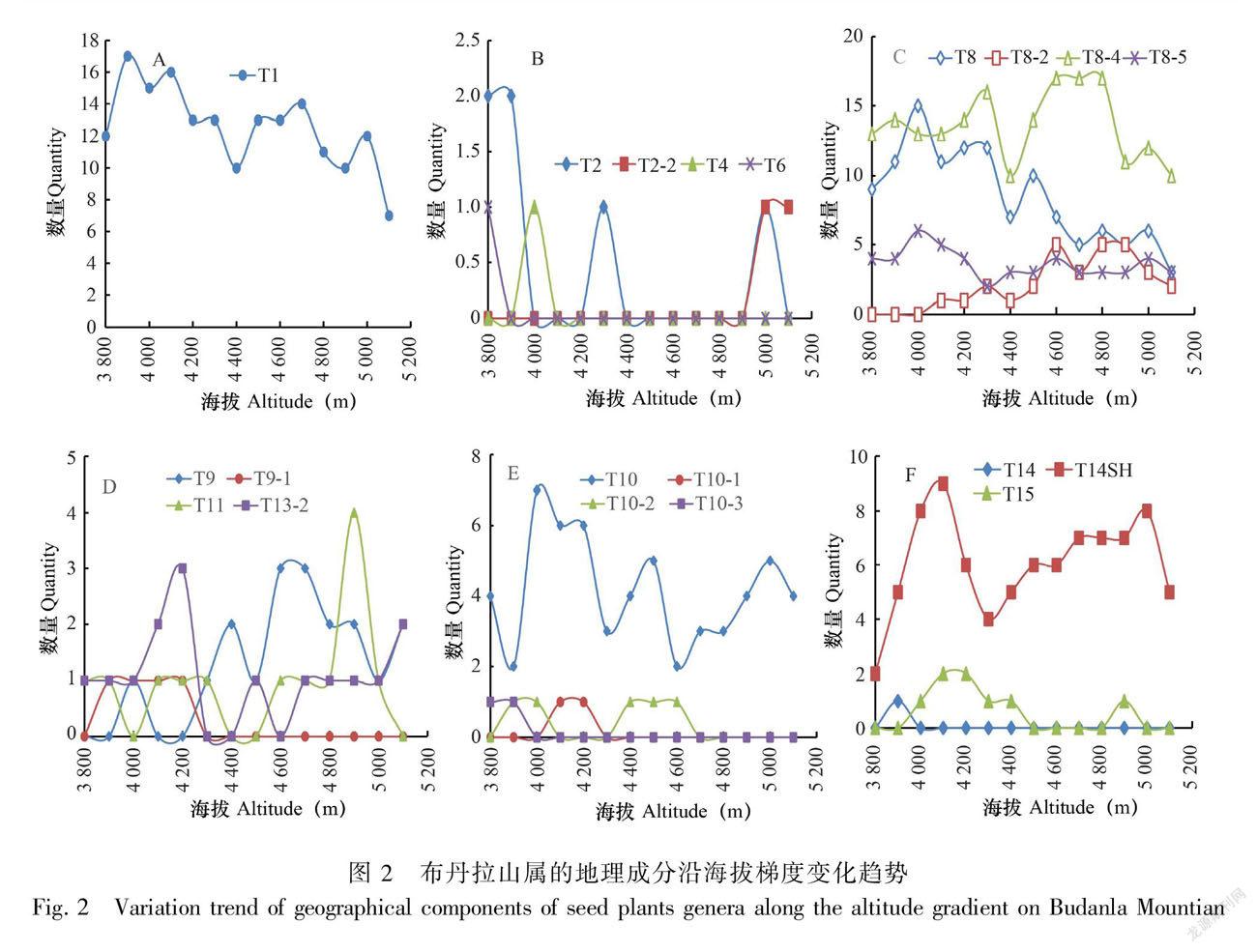
3.3.2 属的区系地理成分沿海拔梯度变化 对属一级的各区系地理成分沿海拔梯度的分布特征进行统计分析,结果如图2所示。
(1)世界广布型(T1):由图2: A可知,世界广布属随海拔的升高,呈现先增后减的趋势,海拔3 900 m时出现最高值,后缓慢下降,变化趋势较平缓,直至海拔最高点5 100 m时出现最低值。
(2)热带分布及其变型(T2、T2-2、T4和T6):由图2:B可知,除了T2(泛热带分布型)属以外,T2-2(热带亚洲、非洲和中美洲至南美洲间断)、T4(旧世界热带分布)和T6(热带亚洲和热带非洲分布)的分布海拔范围都比较狭窄,T2-2型[冷水花属(Pilea)]仅在山顶海拔5 000~5 100 m砾石缝布满苔藓的阴湿地出现,T4型[天门冬属(Asparagus)]仅在海拔4 000 m灌丛中出现和T6型[香茶菜属(Isodon)]仅在3 800 m低海拔谷地灌丛有分布。热带分布属总体趋向低海拔分布格局,在高海拔接近山顶有微弱的上升趋势,这可能与在高海拔极端环境下物种剧烈分化的结果有关。
(3)北温带分布成分及其变型(T8、T8-2、T8-4和T8-5):由图2: C可知,北温带分布成分沿海拔梯度总体呈现先增后减的变化趋势,占比都较稳定,波动起伏不大。T8北温带分布型、变型T8-5欧亚和温带南美洲间断峰值偏向低海拔,在海拔4 000 m时达到最大值,此时T8含15属,T8-5含6属;变型T8-2北极-高山沿海拔的升高缓慢上升至海拔4 600 m时达到最大值,变化趋势缓慢,含5属,后平缓下降;变型T8-4北温带和南温带间断分布在4 600~4 800 m海拔区间段呈现峰值,含17属,在整个海拔梯度始终占据绝对优势。 图2: D中的T11温带亚洲分布型总体上是随海拔先增后减的变化趋势,仅在4 900 m时突然增加且达到最大值,含4属;变型T13-2中亚至喜马拉雅和华西南分布区为中亚东部至喜马拉雅中和中国西南部,其在该研究区峰值出现在海拔4 200 m,含3属,在海拔5 000 m处表现出上升的趋势,可见该地理成分有向高海拔方向发展的趋势。由图2: E可知,温带性质的T10旧世界温带分布型变化的趋势起伏较大,海拔4 000 m时出现最大值;变型T10-1地中海、西亚(或中亚)和东亚间断、T10-2地中海和喜马拉雅间断和T10-3欧亚和南部非洲(有时有大洋洲)间断的分布海拔范围都较狭窄。总体上,T10分布及其变型在该区的分布都集中在低中海拔地段,且随海拔升高逐渐减少。
(4)东亚和北美间断分布及其变型(T9和T9-1):由图2:D可知,T9东亚和北美间断分布型沿海拔的上升呈先增后减趋势,海拔4 600 m时出现峰值,含3属;变型T9-1东亚和墨西哥美洲间断分布海拔上限为4 200 m,仅含1属,此类型的分布较少。
(5)东亚分布及其变型(T14和T14SH):由图2:F可知,T14东亚分布型仅在低海拔3 900 m出现,仅1个属,即野丁香属(Leptodermis),体现布丹拉山处于东亚分布型西缘的特点。东亚地理成分中其变型T14SH中国-喜马拉雅成分占据绝对优势,在该研究区的所有海拔区间段都有分布,在海拔4 100 m时达到峰值,含9属,到海拔4 300 m时迅速下降到4属,后随着海拔上升缓慢增加,到海拔5 000 m时开始下降,含8属,无疑验证了植物区系划分中国-喜马拉雅分布变型的合理性。
(6)中国特有成分(T15):由图2:F可知,T15中国特有属分布型随海拔的升高呈现先增后减的变化趋势,海拔4 100、4 200 m达到峰值,含2属,4 500 m处变为0。布丹拉山在接近山顶的海拔4 900 m处又记录到一中国特有属,中国特有属地理成分的分布格局也显示出了在这一地区物种还在剧烈地演化。
本研究结果表明布丹拉山南坡种子植物优势属和属的区系地理成分沿海拔梯度的分布变化趋势符合普遍的物种丰富度分布格局呈现先增后减的单峰分布格局模式。
4 讨论与结论
布丹拉山处在气候过渡带上且海拔落差大(3 700~5 127 m),山地气候“一山分四季,南北不同天”在这里完美展现,处在生态群落交错带,其南坡是半湿润的气候带,加上海拔和地形的影响,山顶还有流石滩生境植被,此区域山地植物多样性异常丰富、区系成分复杂。本研究区种子植物约有45科156属316种,其中裸子植物1科1属2种,被子植物44科155属314种,说明被子植物占主导地位。属数量结构分析显示,虎耳草属、马先蒿属和龙胆属等生活型全为草本的大属为优势属,可能是草本植物较短的生活史使得它们更快和更好地适应了布丹拉山山区较短暂的生长季条件,并且已有研究也表明这三个属的现代分布和分化中心就在中国-喜马拉雅山区(孙航,2002;Favre et al., 2016)。
从单一的优势属丰富度沿海拔梯度的分布格局分析看,总体上具有先增后减的单峰分布格局,但局部波动起伏变化较大,这是小地形小生境影响下局部水热条件再分配不均的結果(方精云等,2004b),加上优势属所包含的物种数量多,从山脚到山顶的分布范围广泛,但物种的这种垂直分布空间不会很大。在地理成分沿海拔梯度的变化分析中,属级地理成分沿海拔梯度变化也呈现先增后减的变化趋势,个别地理成分分布海拔范围狭窄,所含的属数也非常少,只在某一个海拔区间有分布,说明布丹拉山南坡生境条件对此分布类型的影响很大。因此,变化趋势呈现并不是那么明显的先增后减的变化格局,甚至出现波浪型和双峰型的分布格局,如热带属和中国特有属的成分最明显。总体上热带分布及其变型沿海拔的升高呈逐渐下降的趋势,这也符合热带属性的特点,水热条件对热带成分的影响较大,随着海拔的升高,温度降低,热带成分必然减少。有学者也对喜马拉雅山东部不丹的种子植物丰富度海拔格局进行研究,结果显示物种丰富度峰值在中海拔,温带类群的海拔丰富度峰值集中在中高海拔近似于与海拔对称的单峰格局,热带类群的海拔丰富度高峰集中在低海拔,温度对物种丰富度分布格局影响最大(Jürgen et al., 2017; Li & Feng, 2017)。关于种子植物区系的垂直分布格局研究中,如曾敏等(2021)对卧龙国家级自然保护区的种子植物区系垂直分布格局的研究表明,物种丰富度随海拔升高呈单峰分布格局,热带成分占比下降,温带成分占比上升;布买丽娅木·吐如汗等(2017)对库车种子植物区系垂直分布格局研究结果显示,该区植物科属种丰富度随着海拔的升高呈现先增后减的单峰格局,属的地理成分垂直分布格局也呈现先增后减的变化趋势;刘彬等(2018)对新疆天山南坡中段种子植物区系垂直分布格局进行了系统研究,结果表明植物区系成分沿海拔梯度的变化总体上都是先增后减的变化趋势,个别较小的地理成分波动起伏较大,以上大量温带地区的山地植物多样性区系研究也表明物种丰富度在中海拔区域最高。本研究也得到了相似的结论,即植物区系成分沿海拔梯度有先增后减的变化趋势,但局部的小环境作用对物种丰富度格局的影响很大,进而影响植物区系分布类型的垂直变化。因此,植物区系的垂直分布变化格局可能主要受到当地气候条件与海拔梯度引起的水热条件变化的共同影响。
植物区系地理成分的分析表明,属水平上布丹拉山南坡种子植物的区系呈现温带性质,温带分布型地理成分占绝对优势,其中以北温带分布型及变型为主;种水平上布丹拉山南坡种子植物的区系也是呈现温带性质,温带成分中又以中国-喜马拉雅成分为主,可见该区与喜马拉雅山脈有着密切联系。另外,属种的区系分析结果也符合该区属高原温带半湿润气候的特点。沈泽昊等(2017)对中国14座高山带的植物区系研究显示,高山带有极丰富的种子植物区系,成分主要由北温带分布及其亚型、世界分布、旧世界温带分布和东亚分布及其中国-喜马拉雅分布亚型等成分构成,缺少中国-日本分布变型,有极高的中国特有属比例,本文对布丹拉山南坡种子植物区系的研究结果与其一致。极端生境和物种迁入率低的高海拔被认为有较高的物种特有率,特有属沿海拔会出现多种分布变化格局(Lomolino, 2001; Kessler, 2002; Vetaas & Grytnes, 2002; Jürgen et al., 2017)。布丹拉山在接近山顶的海拔4 900 m处又记录到一中国特有属,中国特有属成分的分布格局也显示出了在这一地区物种的分化还在剧烈地演绎,也为高海拔物种特有率高的观点补充了新资料。
综上所述,布丹拉山作为印度洋暖湿气流沿雅鲁藏布江向高原面输送的最后一道天然屏障之一,形成了天然的生态群落交错区,造成了南北坡迥然不同的气候类型,其南坡气候温暖湿润,因此山地植物多样性异常丰富,植被茂盛。由布丹拉山南坡种子植物多样性结构分析结果可知,此区单种属和小属占据主导地位。由区系分析结果可知,属种区系成分复杂多样,以温带分布型为优势,尤其以中国-喜马拉雅分布亚型为主,这也完全符合此区域的植物地理位置。从生活型上看,较短生活史的草本植物占据主导地位。区系地理成分和优势属丰富度沿海拔的分布格局呈现先增后减的单峰变化趋势,基本保持一致,植物区系成分的海拔格局变化也符合物种丰富度分布格局研究中的中间膨胀效应假说,即认为物种丰富度在中海拔地区最高(Colwell & Lees, 2000;王志恒等,2004)。有研究也认为低海拔地区温度高且蒸发量大导致水分不足,高海拔地区水分充足而热量不足,中低海拔是水热组合最适的生境,因此物种组成丰富(王国宏,2002;王志恒等,2004)。本研究得出结论是,布丹拉山南坡种子植物区系的成分在中低海拔区域最为复杂多样,且区系成分占据主导地位的温带分布型属也在中低海拔4 000 m时达到峰值,同时可能受小生境和小地形等大量复杂环境因子影响而局部波动起伏较大,但总体上呈现偏峰分布格局的变化趋势。本文研究的不足之处,由于缺少每一个调查样方的水热条件具体的环境指标,如土壤含水量、温度、湿度和土壤氮磷钾等具体环境因子指标,所以无法深入讨论这种植物区系海拔格局的形成机制,只能根据目前关于雅鲁藏布江流域植物区系的研究来推测判断。
参考文献:
BIAN J, 2006. The beautiful of Gyaca[J]. China’s Tibet, (2): 56-61. [边吉, 2006. 秀美加查[J]. 中国西藏, (2): 56-61.]
CHEN FY, 2009. Thousand year old walnut woods at Gyaca[J]. Tibet Tour,(8): 98-101. [陈芳媛, 2009. 加查千年核桃林[J]. 西藏旅游, (8): 98-101.]
CHEN LZ, SUN H, GUO K, 2015. China flora and vegetation geography[M]. Beijing: Science Press: 30-163. [陈灵芝, 孙航, 郭柯, 2015. 中国植物区系与植被地理[M]. 北京: 科学出版社: 30-163.]
COLWELL RK, LEES DC, 2000. The mid-domain effect: geometric constraints on the geography of species richness[J]. Trend Ecol Evolut, 15(2): 70-76.
DEVADOSS J, FALCO N, DAFFLON B, et al., 2020. Remote sensing-informed zonation for understanding snow, plant and soil moisture dynamics within a mountain ecosystem[J]. Remote Sens, 12(17): 2733.
Delectis Florae Reipublicae Popularis Sinicae Agendae Academiae Sinicae Edita, 1959-2004. Flora Reipublicae Popularis Sinicae (vol. 1-80)[M]. Beijing: Science Press. [中国科学院中国植物志编辑委员会, 1959-2004. 中国植物志(1-80卷)[M]. 北京: 科学出版社.]
FANG JY, SHEN ZH, CUI HT, 2004a. Ecological characteristics of mountains and research issues of mountain ecology[J]. Biodivers Sci, (1): 10-19. [方精云, 沈泽昊, 崔海亭, 2004a. 试论山地的生态特征及山地生态学的研究内容[J]. 生物多样性, (1): 10-19. ]
FANG JY,KANZAKI MR, WANG XP, et al., 2004b, Community structure of alpine sparse vegetation and effects of microtopography in Pushila, Everest-Choyu region, Tibet, China[J]. Biodivers Sci, (1): 190-199. [方精云, 神崎护, 王襄平, 等, 2004b. 西藏珠峰-卓奥友峰普士拉地区高山稀疏植被的群落特征及小地形的影响[J]. 生物多样性, (1): 190-199.]
FAVRE A, MICHALAK I, CHEN CH, et al., 2016. Out-of-Tibet: the spatio-temporal evolution of Gentiana (Gentianaceae)[J]. J Biogeogr, 43(10): 1967-1978.
Gyaca Local Records Compilation Committee, 2010. Gyaca counties annals[M]. Beijing: China Tibetology Press: 1-976. [加查县地方志编纂委员会, 2010. 加查县志[M]. 北京: 中国藏学出版社: 1-976]
JRGEN K, SEBASTIAN W, SIMON L, et al., 2017. Elevational seed plants richness patterns in Bhutan, Eastern Himalaya[J]. J Biogeogr, 44(8): 1711-1722.
KESSLERM, 2002. The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels[J]. J Biogeogr, 29(9): 1159-1165.
LIU MJ, LI HJ, 2020. Rare wildlife resources and protection in national nature reserve for black-necked cranes in the middle reaches of the brahmaputra river valley in Tibet[J]. Cent S For Invent Plan, 39(2): 57-61. [刘敏杰, 李华军, 2020. 西藏雅鲁藏布江中游河谷黑颈鹤国家级自然保护区珍稀野生动植物资源与保护[J]. 中南林业调查规划, 39(2): 57-61.]
LI C, XU WL, LI QK, et al., 2021. Community structure and diversity distribution pattern of sandy plants in the middle and upper reaches of the Yarlung Zangbo River[J]. J Resour Ecol, 12(1): 11-21.
LA Q, ZHA XCR, ZHU WD, et al., 2014. Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet, China[J]. Biodivers Sci, 22(3): 337-347. [拉瓊, 扎西次仁, 朱卫东, 等, 2014. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释[J]. 生物多样性, 22(3): 337-347.]
LI M, FENG JM, 2017. Biogeographical interpretation of elevational patterns of genus diversity of seed plants in Nepal[J]. PLoS ONE, 10(10): e0140992.
LIU B,TURGAN B, KEREMU A, et al., 2018. Vertical distribution patterns of the seed plant flora in the middle section on the southern slopes of the Tianshan Mountains in Xinjiang, China[J]. Plant Sci J, 36(2): 191-202. [刘彬, 布买丽娅木·吐如汗, 艾比拜姆·克热木, 等, 2018. 新疆天山南坡中段种子植物区系垂直分布格局分析[J]. 植物科学学报, 36(2): 191-202.]
LOMOLINO MV, 2001. Elevation gradients of species-density: historical and prospective views[J]. Global Ecol Biogeogr, 10(1): 3-13.
MING SP, ZHONG Y, XU M, et al., 2018. Comparison of floristic composition and characteristics of vascular plants on the northern and southern slopes of Gambo Utse Mountain, Lhasa[J]. Plateau Sci Res, 2(1): 61-72. [明升平, 钟杨, 许敏, 等, 2018. 拉萨根培乌孜山阴阳坡维管束植物区系组成及特征比较[J]. 高原科学研究, 2(1): 61-72.]
QU XL, GUO WW, LUO DQ, et al., 2019. Composition and characteristics of subalpine shrub-grass communities in the Yarlung Zangbo River valley[J]. J SW Univ (Nat Sci Ed), 41(3): 1-8. [屈兴乐, 郭文文, 罗大庆, 等, 2019. 雅鲁藏布江河谷山地灌丛草地区系组成及特征[J]. 西南大学学报(自然科学版), 41(3): 1-8.]
SHEN ZH, YANG MZ, FENG JM, et al., 2017. Geographic patterns of alpine flora in China in relation to environmental and spatial factors[J]. Biodivers Sci, 25(2): 182-194. [沈泽昊, 杨明正, 冯建孟, 等, 2017. 中国高山植物区系地理格局与环境和空间因素的关系[J]. 生物多样性, 25(2): 182-194. ]
SUN H, 2002.Tethys retreat and Himalayas-Hengduanshan mountains uplift and their significance on the origin and development of the Sino-Himalayan elements and alpine Flora[J]. Acta Bot Yunnan, 24(3): 273-288. [孙航, 2002. 古地中海退却与喜马拉雅-横断山的隆起在中国喜马拉雅成分及高山植物区系的形成与发展上的意义[J]. 云南植物研究, 24(3): 273-288. ]
TITO R, VASCONCELOS HL, FEELEY KJ, 2020. Mountain ecosystems as natural laboratories for climate change experiments[J]. Front For Glob Chang, 3(38): 1-8.
TURGAN B, LIU B, YIMIT AKBR, 2017.Analysis on vertical distribution pattern of flora of seed plants in Kuche[J]. Guihaia, 37(4): 453-460. [布買丽娅木·吐如汗, 刘彬, 艾克拜尔·依米提, 2017. 库车种子植物区系垂直分布格局分析[J]. 广西植物, 37(4): 453-460.]
VETAASOR, GRYTNES JA, 2002. Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal[J]. Global Ecol Biogeogr, 11(4): 291-301.
WANG GH, 2002. Species diversity of plant communities along an altitudinal gradient in the middle section of northern slopes of Qilian Mountains Zhangye Gansu China[J]. Biodivers Sci, 10 (1): 7-14. [王国宏, 2002. 祁连山北坡中段植物群落多样性的垂直分布格局[J]. 生物多样性, 10(1): 7-14.]
WANG HS, 1992. Floristic geography[M]. Beijing: Science Press: 1-176. [王荷生, 1992. 植物区系地理[M]. 北京: 科学出版社: 1-176.]
WANG ZH, CHEN AP, PIAO SL, et al., 2004.Pattern of species richness along an altitudinal gradient on Gaoligong Mountains , Southwest China[J]. Biodivers Sci, 12 (1): 82-88. [王志恒, 陈安平, 朴世龙, 等, 2004. 高黎贡山种子植物物种丰富度沿海拔梯度的变化[J]. 生物多样性, 12(1): 82-88.]
WU ZY, 1979. On China flora partition problem[J]. Acta Bot Yunnan, 1(1): 1-20. [吴征镒, 1979. 论中国植物区系的分区问题[J]. 云南植物研究, 1(1): 1-20.]
WU ZY, ZHOU ZK, SUN H, et al., 2006. The areal-types of seed plants and their origin and differentiation[M]. Kunming: Yunnan Science and Technology Press: 1-566. [吴征镒, 周浙昆, 孙航, 等, 2006. 种子植物分布区类型及其起源和分化[M]. 昆明: 云南科技出版社: 1-566.]
WU ZY, ZHOU ZK, LI DZ, et al., 2003. The area-types of the world families of seed plants[J]. Acta Bot Yunnan, 25 (3): 245-257. [吴征镒, 周浙昆, 李德铢, 等, 2003. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 25 (3): 245-257.]
XUE HB, 2015. Research on the development of towns in Shannan prefecture of Tibet[D]. Beijing: Beijing University of Civil Engineering and Architecture: 5-81. [薛鸿博, 2015. 西藏山南地区城镇发展研究[D]. 北京: 北京建筑大学: 5-81.]
YU YY, LI J, ZHOU ZX, et al., 2021. Response of multiple mountain ecosystem services on environmental gradients: How to respond, and where should be priority conservation? [J]. J Clean Prod, (278): 123264.
ZENG M, MA YH, SHEN WT, et al., 2021. Vertical distribution pattern of seed plants flora in Wolong national nature reserve[J]. J Chin West Norm Univ (Nat Sci Ed), 42(2): 110-115. [曾敏, 馬永红, 沈文涛, 等, 2021. 卧龙国家级自然保护区种子植物区系垂直分布格局[J]. 西华师范大学学报(自然科学版), 42(2): 110-115.]
ZHANG J, CAIWEN DJ, XIE YP, et al., 2019. Characteristics on the flora of seed plants in Sanjiangyuan national park[J]. Acta Bot Boreal-Occident Sci,39(5): 935-947. [张静, 才文代吉, 谢永萍, 等, 2019. 三江源国家公园种子植物区系特征分析[J]. 西北植物学报, 39(5): 935-947.]
ZHOU ZK, HUANG J, DING WN, 2017. The impact of major geological events on Chineses flora[J]. Biodivers Sci, 25(2): 123-135. [周浙昆, 黄健, 丁文娜, 2017. 若干重要地质事件对中国植物区系形成演变的影响[J]. 生物多样性, 25(2): 123-135.]
ZHU S, 2012. River landform and geology environment evolution in the Yarlung Zangbo River valley[D]. Beijing: Chinese Academy of Geological Science: 1-153. [祝嵩, 2012. 雅鲁藏布江河谷地貌与地质环境演化[D]. 北京: 中国地质科学院: 1-153.]
(责任编辑 蒋巧媛)
收稿日期:2021-08-28
基金项目:国家自然科学基金(31760127) [Supported by National Natural Science Foundation of China (31760127)]。
第一作者: 王俊伟(1994-),硕士研究生,主要研究方向为植物多样性,(E-mail)jwyx12240315@126.com。
通信作者:拉琼,博士,教授,博士研究生导师,主要研究方向为植物多样性与进化生态,(E-mail)lhagchong@163.com。
猜你喜欢
科技创新导报(2017年10期)2017-06-19
绿色科技(2017年9期)2017-06-01
科学与财富(2017年13期)2017-05-31
安徽农学通报(2016年15期)2017-05-12
农业工程技术·综合版(2016年8期)2017-04-22
绿色科技(2017年5期)2017-03-31
农业与技术(2016年22期)2017-03-07
中国高新技术企业(2016年32期)2016-12-27
湖北农业科学(2016年18期)2016-12-08
