
西伯利亚杏不同树冠光合特性及产量差异
2022-04-18 04:49陈建华孙永强丁士富郑永涛董胜君
河南农业科学 2022年1期
张 剑,陈建华,孙永强,丁士富,郑永涛,董胜君
(1. 沈阳农业大学林学院,辽宁 沈阳 110866;2. 北票市林木良种繁育中心,辽宁 北票 122102)
西伯利亚杏[Armeniaca sibirica(L.)Lam.]是蔷薇科杏属植物,是亚洲特有的“三效合一”树种,具有广泛的应用前景,极具开发潜力[1]。西伯利亚杏生态适应性强,是干旱半干旱地区的重要造林树种;同时,其杏仁因含有苦杏仁甙、杏仁蛋白、维生素及微量元素硒等物质可作为药物、保健食品的原材料[2]。研究发现,西伯利亚杏主枝基角是影响产量的主导因子[3],并且可直接影响树形和冠层内空间结构。不同树形、冠层内的枝叶数量、比例和空间分布各不相同,从而导致树冠内光照、温度、水分的二次分配不同,最终形成一个复杂的微气候环境,最终直接或间接地影响树木生长发育和果实产量及品质[4-6]。CASTILLORUIZ等[7]研究发现,橄榄果实[Canarium album(Lour.)Rauesch.]总产量的60%来自树冠中、外和上部。 库尔勒香梨(Pyrus sinkiangensisYü)疏散分层形平均单株果实产量高达1 116 个,是所有试验树形平均值的2.88 倍[8],并且疏散分层形和圆柱形外围的净光合速率均高于内膛[9]。在枣(Ziziphus jujubMill.)不同树形中,其冠层中上层的果实品质要优于下层[10]。TRAD 等[11]对无花果(Ficus caricaL.)研究发现,果实在冠层中的位置对果实的硬度、可滴定酸度有显著影响,并且冠层外部的果实比树冠下部果实甜度、成熟度更高,单果质量也更大。随着杏属植物多年开发利用,植株年龄不断增大且缺乏管理,树冠逐渐郁闭,枝叶相互遮掩,导致光能利用率降低、树冠有效结果区减小,造成果实产量与品质下降,其树冠结构的研究需要愈发迫切,但关于杏属植物不同树冠结构的研究鲜见报道。因此,以西伯利亚杏无性系作为试验材料,通过主枝基角开张角度对其树形进行分级,基于此测定其不同生长时期、不同树形、不同冠层部位光合生理特性指标及产量,进而明确西伯利亚杏高产树形及结果部位,最终为培育其高产树冠结构提供理论依据和技术指导。
1 材料和方法
1.1 试验地概况
试验地设在沈阳农业大学山杏种质资源圃,位于辽宁省北票市泉巨永乡,东经120°18′,北纬42°25′,为温带大陆性季风气候;冬季长而干冷,夏季短而多雨,年均气温8.6 ℃,无霜期约153 d,年均蒸发量2 000 mm,年均降水量509 mm。该地属于丘陵山区,平均海拔200 m。土层厚度50~100 cm,土壤为褐土、砂壤,肥力中等,pH值7.0~7.2。
1.2 试验材料
供试材料为资源圃内自然生长的74 个西伯利亚杏无性系,树龄14~15 a,株行距2 m×3 m。每个无性系选择3株生长一致且长势良好的样木进行各项指标测定。
1.3 试验方法
于2018年4月,测定每个样木主枝基角α,并采用KS 检验对其主枝基角进行正态性检验。对符合正态分布的主枝基角参照刘孟军[12]的方法进行3 级概率分级。于同年7月下旬测定结实量并采集不少于10个果实测定平均单果质量(精准至0.1 g),计算单株产量(单果质量×结实量,精准至0.1 g)。
基于概率分级结果按主枝基角大小对西伯利亚杏进行树形分类,每类选取1个无性系(每个无性系3 株)作为代表,参照魏钦平等[13]的方法,将冠层由内到外均分为内部、中部和外部,于7月中旬分树形、冠层统计单株产量(精准至0.1 g);于2018 年6月下旬、7 月下旬、9 月中旬,每天6:00—18:00 每2 h 对各冠层部位内当年生枝条自上而下第5~7 片叶使用Licor-6400XT 便携式光合仪进行测定,每个叶片重复3次,测定指标包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和光照强度(PAR)。
1.4 数据处理
使用Excel 2019对测定数据进行整理、作图、分析,利用SPSS 22.0进行多重比较。
2 结果与分析
2.1 西伯利亚杏主枝基角概率分级
由表1 可知,西伯利亚杏主枝基角Sig.值大于0.05,表明其符合正态分布,可用于概率分级。根据概率分级3 级分级法,将西伯利亚杏按主枝基角开张角度分为3级,结果如表2所示。其中3级的产量极显著高于2 级和1 级,同时2 级产量极显著高于1级。因此,可根据主枝基角开张角度将西伯利亚杏分为紧密形(1 级)、均匀形(2 级)、开阔形(3 级)3种树形。

表1 西伯利亚杏主枝基角正态性检验Tab.1 Test of normality for main branch angle of Armeniaca sibirica
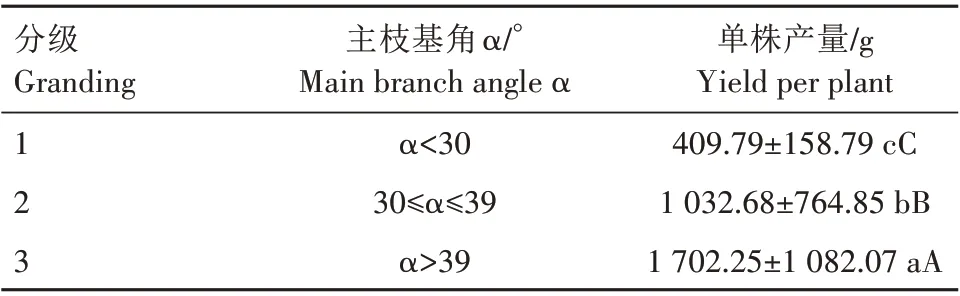
表2 西伯利亚杏主枝基角对单株产量的影响Tab.2 Effects of main branch angle on yield per plant of Armeniaca sibirica
2.2 西伯利亚杏不同树形、不同冠层部位光合日变化分析
2.2.1 光照强度 由图1 可知,西伯利亚杏紧密形与均匀形各冠层部位在不同时期的光照强度日变化均为单峰曲线,而开阔形各冠层部位在6、7 月为单峰曲线(图1-Ⅰ、Ⅱ),但在9月呈双峰曲线(图1-Ⅲ);各树形各冠层光照强度平均值在各时期总体趋势均为均匀形[1 012.29 μmol/(m2·s)]>开阔形[1 006.69 μmol/(m2·s)] >紧密形[971.11 μmol/(m2·s)],外部[1 059.26 μmol/(m2·s)]>中部[1 009.85 μmol/(m2·s)] >内部[920.99 μmol/(m2·s)],6 月[1 252.41 μmol/(m2·s)]>9 月[900.90 μmol/(m2·s)]>7月[836.78 μmol/(m2·s)]。

图1 西伯利亚杏不同树形、不同冠层部位各时期的光照强度日变化Fig.1 Diurnal variation of PAR in different tree shapes and different canopies of Armeniaca sibirica
2.2.2 净光合速率 6 月,开阔形、紧密形、均匀形西伯利亚杏各冠层净光合速率日变化均为单峰曲线(图2-Ⅰ),呈倒“V”形。其中,开阔形、均匀形西伯利亚杏的净光合速率变化一致,自6:00 起逐步上升,12:00 到达峰值,后逐渐下降;而紧密形各冠层净光合速率峰值出现在14:00。
7 月,紧密形、均匀形各冠层及开阔形内部净光合速率为双峰曲线(图2-Ⅱ),呈“M”形。其中,均匀形、紧密形各冠层在10:00 出现第1 个且最大峰值,12:00 发生“光合午休”,14:00 出现第2 个峰值;开阔形内部在10:00 出现第1 个峰值,随后进行“光合午休”,不同的是,其第2个峰值出现在16:00。而开阔形外部、中部净光合速率呈单峰曲线,在10:00达到峰值。
9 月,紧密形各冠层净光合速率日变化呈双峰曲线(图2-Ⅲ),2 个峰值分别出现在10:00、14:00,12:00 发生“光合午休”。均匀形、开阔形各冠层均呈单峰曲线,在12:00达到峰值。

图2 西伯利亚杏不同树形、不同冠层部位各时期的净光合速率日变化Fig.2 Diurnal variation of Pn in different tree shapes and different canopies of Armeniaca sibirica
不同树形西伯利亚杏各冠层在各时期的净光合速率总体趋势均为开阔形[10.44 μmol/(m2·s)]>均匀形[10.14 μmol/(m2·s)]>紧密形[9.45 μmol/(m2·s)],外部[11.20 μmol/(m2·s)]>中部[10.06 μmol/(m2·s)]>内部[8.78 μmol/(m2·s)],6 月[12.09 μmol/(m2·s)]>7 月[9.98 μmol/(m2·s)]>9 月[7.97 μmol/(m2·s)]。此外,6、7、9 月的最大净光合速率均在开阔形外部,分别是19.78、15.01、13.65 μmol/(m2·s),而每月12:00 最小净光合速率均在紧密形内部,分别是13.24、9.46、9.09 μmol/(m2·s)。
2.2.3 蒸腾速率 6 月,均匀形、紧密形西伯利亚杏各冠层蒸腾速率日变化均呈单峰曲线(图3-Ⅰ),其中均匀形各冠层部位的峰值出现在12:00,紧密形各冠层蒸腾速率峰值出现在14:00;而开阔形各冠层蒸腾速率日变化呈双峰曲线,第1 个峰值出现在10:00,第2个峰值较大,出现在14:00。
7 月,西伯利亚杏最大蒸腾速率出现在开阔形外部,为6.75 mmol/(m2·s),并且各树形各冠层部位的蒸腾速率日变化均为单峰曲线(图3-Ⅱ),其中开阔形各冠层的峰值出现在12:00,而紧密形和均匀形各冠层的峰值稍晚,出现在14:00。
9 月,开阔形外部蒸腾速率最大,为4.74 mmol/(m2·s)。紧密形各冠层蒸腾速率日变化呈双峰曲线,10:00 达到第1 个峰值,12:00 发生“光合午休”,14:00出现第2个更大的峰值(图3-Ⅲ);均匀形及开阔形各冠层部位的蒸腾速率日变化均呈单峰曲线,且均在14:00达到峰值。
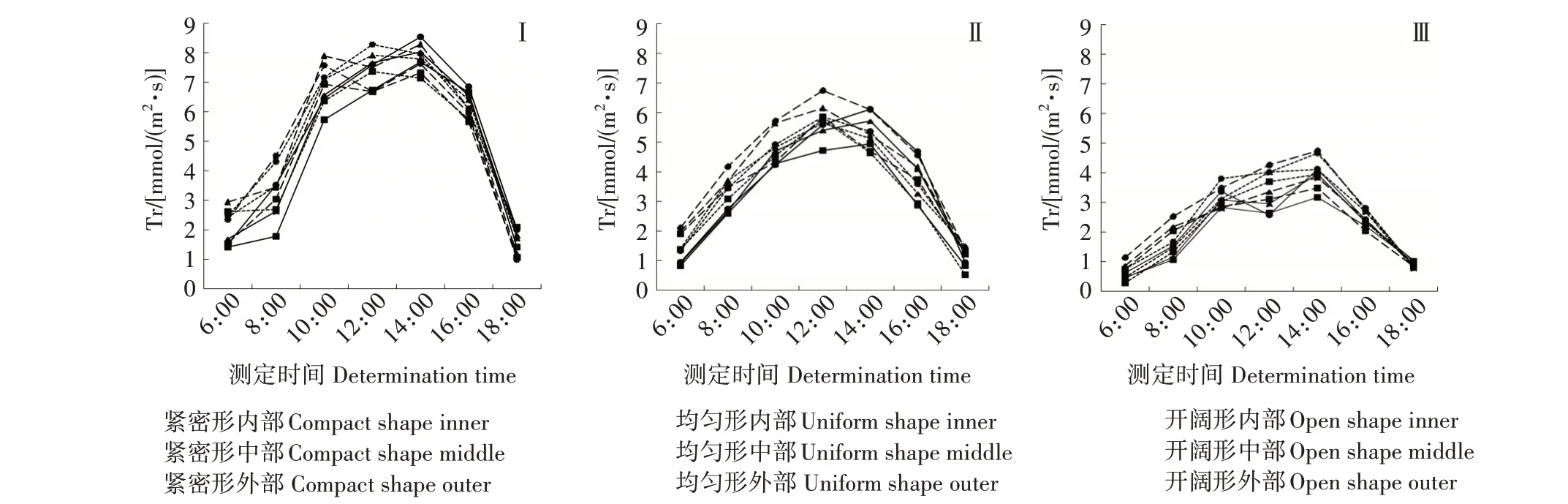
图3 西伯利亚杏不同树形、不同冠层部位各时期的蒸腾速率日变化Fig.3 Diurnal variation of Tr in different tree shapes and different canopies of Armeniaca sibirica
不同树形西伯利亚杏各冠层部位在不同时期的蒸腾速率总体表现均为开阔形[3.82 mmol/(m2·s)]>均匀形[3.71 mmol/(m2·s)]>紧密形[3.46 mmol/(m2·s)],外部[3.91 mmol/(m2·s)]>中部[3.74 mmol/(m2·s)]>内部[3.35 mmol/(m2·s)],6 月[5.03 mmol/(m2·s)] >7 月[3.64 mmol/(m2·s)] >9 月[2.31 mmol/(m2·s)],且每个月14:00 之后急速下降,18:00各树形各冠层部位的蒸腾速率趋于一致,达到最低。7月和9月最大蒸腾速率均出现在开阔形外部,且12:00 最低蒸腾速率均为紧密形内部,与净光合速率表现一致。
2.2.4 气孔导度 6 月,西伯利亚杏各树形各冠层气孔导度日变化均呈单峰曲线。其中,均匀形和紧密形各冠层气孔导度峰值出现在12:00,而开阔形各冠层峰值出现在14:00(图4-Ⅰ)。
7 月,西伯利亚杏各树形各冠层部位的气孔导度日变化均为双峰曲线,其中,各树形各冠层部位气孔导度的第1 个且最大峰值均出现在10:00,最大值为开阔形外部[0.22 mol/(m2·s)]。紧密形和均匀形各冠层部位气孔导度的第2 个峰值出现在14:00,而开阔形各冠层部位气孔导度的第2个峰值出现在16:00(图4-Ⅱ)。
9 月,西伯利亚杏各树形各冠层部位的气孔导度均为双峰曲线,其中,各树形各冠层部位气孔导度的第1 个且最大峰值均出现在8:00,最大值为开阔形外部[0.19 mol/(m2·s)]。紧密形和均匀形各冠层部位气孔导度的第2 个峰值出现在14:00,而开阔形各冠层部位气孔导度的第2 个峰值稍早,出现在12:00(图4-Ⅲ)。

图4 西伯利亚杏不同树形、不同冠层部位各时期的气孔导度日变化Fig.4 Diurnal variation of Gs in different tree shapes and different canopies of Armeniaca sibirica
西伯利亚杏各树形各冠层部位各时期的气孔导度总体趋势为开阔形[0.15 mol/(m2·s)]=紧密形[0.15 mol/(m2·s)]>均匀形[0.14 mol/(m2·s)],外部[0.16 mol/(m2·s)]>中部[0.15 mol/(m2·s)]>内部[0.13 mol/(m2·s)],6 月[0.22 mol/(m2·s)] >7 月[0.13 mol/(m2·s)]>9 月[0.09 mol/(m2·s)]。7、9 月,气孔导度的第1 个峰值和第2 个峰值均为开阔形外部最大,且12:00 紧密形内部气孔导度最低,与净光合速率、蒸腾速率表现一致。
2.3 西伯利亚杏不同树形、不同冠层部位最大净光合速率分析
如图5 所示,不同树形西伯利亚杏不同冠层在各时期的最大净光合速率均表现为6 月>7 月>9 月,开阔形>均匀形>紧密形,冠层外部>冠层中部>冠层内部。6 月(图5-Ⅰ),开阔形外部最大净光合速率最大,为19.78 μmol/(m2·s)。树形方面,开阔形中部极显著高于紧密形中部、均匀形中部,同时,均匀形中部显著高于紧密形中部;开阔形内部极显著高于均匀形内部,显著高于紧密形内部,但紧密形内部与均匀形内部间无显著性差异;开阔形外部极显著高于紧密形外部,显著高于均匀形外部,而紧密形外部与均匀形外部差异不显著。冠层方面,均匀形外部最大净光合速率极显著高于其内部,显著高于中部,同时其中部显著高于内部;紧密形外部极显著高于其内部、中部,而内部和中部间无显著差异;开阔形外部极显著高于其中部、内部,并且其中部极显著高于内部。
7 月(图5-Ⅱ),开阔形外部最大净光合速率最大,为15.01 μmol/(m2·s)。树形方面,开阔形中部最大净光合速率显著高于紧密形中部、均匀形中部;各树形内部无显著差异;开阔形外部极显著高于紧密形外部,显著高于均匀形外部。冠层方面,均匀形外部最大净光合速率极显著高于其中部、内部,而中部和内部之间差异不显著;紧密形各冠层部位无显著差异;开阔形中部、外部均极显著高于其内部,同时,开阔形外部显著高于其中部。
9 月(图5-Ⅲ),开阔形外部最大净光合速率最大,为13.65 μmol/(m2·s)。树形方面,均匀形内部极显著高于紧密形内部、开阔形内部,而开阔形内部与紧密形内部间差异不显著;在冠层外部,各树形间无显著差异,同时在冠层中部,各种树形间差异也不显著。冠层方面,均匀形外部最大净光合速率显著高于其内部,而其中部和外部之间、中部和内部之间无显著差异;开阔形外部极显著高于其内部,显著高于其中部,而中部和内部间差异不显著;紧密形外部极显著高于其内部,但与其中部差异不显著,并且其中部显著高于内部。
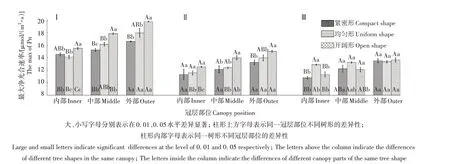
图5 西伯利亚杏不同树形、不同冠层部位各时期的最大净光合速率Fig.5 The max of Pn of Armeniaca sibirica in different tree shapes and different canopies
2.4 西伯利亚杏不同树形、不同冠层部位间产量差异分析
由表3 可知,开阔形西伯利亚杏单株产量(1 303.00 g)极显著高于均匀形(424.31 g)、紧密形(150.73 g),同时均匀形极显著高于紧密形。西伯利亚杏产量主要来自于冠层外部,占比53.61%,可达1 006.88 g,极显著高于冠层内部、中部,同时,冠层中部产量极显著高于冠层内部。
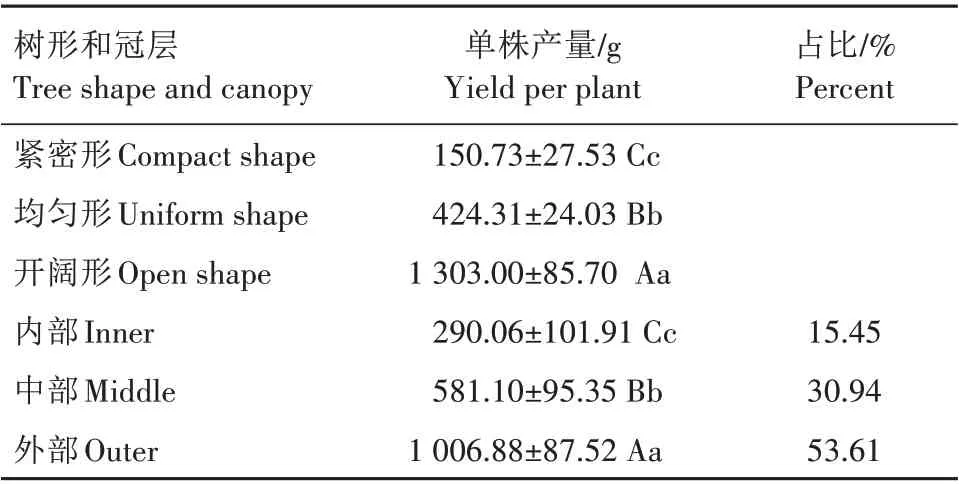
表3 西伯利亚杏不同树形、不同冠层部位间产量差异Tab.3 Yield differences of Armeniaca sibirica with different tree shapes and canopies
西伯利亚杏不同树形、不用冠层部位对产量的交互作用具体表现为(表4):树形方面,在冠层外部和中部,开阔形产量均极显著高于均匀形、紧密形;在冠层内部,开阔形产量显著高于均匀形,且极显著高于紧密形;均匀形外部产量显著高于紧密形外部,但中部、内部产量与紧密形中部、内部均无显著性差异。冠层方面,开阔形外部极显著高于其内部,显著高于其中部,同时其中部产量显著高于其内部;均匀形各冠层部位产量均存在极显著差异,且外部>中部>内部;紧密形外部产量极显著高于其中部、内部。开阔形外部、均匀形外部、紧密形外部产量分别占整株产量的52.43%、56.08%、56.88%,远高于自身中部、内部,其中开阔形外部产量最高,可达683.20 g。

表4 西伯利亚杏不同树形、不同冠层部位交互作用的产量差异Tab.4 Yield difference of different tree shapes and canopies of Armeniaca sibirica
3 结论与讨论
传统树形分类方法均为基于生产经验的目视分类法,虽然简单易行,但各树形的概率分布不一,且存在主观因素,难以形成统一的标准。而概率分级法的分级结果具有较强的客观性、可比性和通用性[14]。本研究按主枝基角开张角度将西伯利亚杏分为3种树形,使树形分级变得有据可依。
光合作用的日变化是植物对各种生理生态因子综合影响的反映,是分析环境因子与植物生长代谢之间关系的重要手段[15]。冠层内的光照强度是光在冠层内分布状况的最直接体现。本试验中,西伯利亚杏均匀型冠层外部光照强度最佳。净光合速率是光合作用的直接体现,可以反映植物光合作用积累有机物的速度[16]。大量研究表明,在一定范围内,树冠外围的净光合速率高于内部,主枝开张角度大的净光合速率更大,如南方红豆杉(Taxus chinensisvar.mairei)不同冠层净光合速率均表现为外部>内部[17],富士苹果(Malus domesticaBorkh. cv.‘Fuji’)110°拉枝处理的净光合速率极显著高于55°、70°、90°、120°处理[18],本研究中也发现类似现象,即西伯利亚杏净光合速率峰值表现为冠层外部>中部>内部、开阔形>均匀形>紧密形。蒸腾速率是植物体内降温增湿能力和物质运输能力的直接体现[19]。本研究中,西伯利亚杏冠层外部的蒸腾速率大于中部、内部,表明冠层外部具有更强的物质运输能力,这与长白落叶松(Larix olgensisHenry)树冠上层的蒸腾速率表现一致[20]。气孔导度可以反映植物气孔传导二氧化碳和水汽的能力,是影响植物蒸腾速率和净光合速率的重要因素[21-22]。本研究中,西伯利亚杏各树形各冠层部位各时期的气孔导度在6 月均为单峰曲线,峰值出现在12:00 和14:00;在7月,除均匀形内部为单峰曲线外,其余均呈双峰曲线,且均在10:00 达到第1 个峰值,但第2 个峰值出现的时间有所差别,紧密形和均匀形各冠层部位气孔导度第2 个峰值出现在14:00,而开阔形第2 个峰值出现于16:00。在格木[23]与灰枣[24]中也存在类似现象。
不同树形冠层结构的特点和微环境不同,产量也因此各不相同。适宜的树形可以改善树冠内光照条件,促进光合作用,最终提高果实产量[25-26]。因此,培育合理树形结构、改善树冠内通风透光是至关重要的。本研究发现西伯利亚杏冠层外部的产量极显著高于冠层中部、内部,占比可达整株树产量的53.61%,并且其开阔形产量极显著高于均匀形、紧密形,表明冠层越外部产量越高,树冠越开阔产量越高。 WEN 等[27]也发现板栗(Castanea mollissimaBlume)冠层外部和上部的产量更高。综合分析树形和冠层交互作用对产量的影响发现,开阔形外部单株产量最高,可达683.20 g,显著高于其他处理,这可能是由于冠层外部光照、通风条件好,物质运输、转化、积累更快,并且树冠越开阔,其外表面积越大,参与高效光合的枝组越多,产量越高。同时,当外层光照充足时,会形成更多的结果枝,从而阻挡光照达到冠层中部和内部,导致中部、内部光照不足,影响树木坐果与果实发育,甚至导致落果,最终严重影响产量[28-29]。本研究中开阔形外部产量比例较均匀形、紧密形偏小,并且开阔形中部和内部占比分别高于均匀形、紧密形的中部和内部,分析认为这可能是开阔树形的中部、内部光照增加,光合作用加强,从而中部、内部产量比例上升,最终促进果实发育和整株增产。
综上所述,西伯利亚杏各树形各冠层部位各时期的净光合速率、蒸腾速率、气孔导度及产量总体表现为6 月高于7、9 月,外部>中部>内部,开阔形>均匀形>紧密形。其中,开阔形西伯利亚杏冠层外部最大净光合速率在各月均极显著高于其内部。6月,开阔形外部最大净光合速率极显著高于紧密形外部,显著高于均匀形外部。开阔形西伯利亚杏产量极显著高于紧密形、均匀形,冠层外部产量极显著高于内部、中部。因此,在西伯利亚杏树体管理中应以开阔形为主,可以通过主枝拉枝,改善树形,增加树冠内通风、透光条件及受光枝组比例,以优化树冠结构,促进光合作用,最终达到增产、稳产的效果。
猜你喜欢
中国畜牧杂志(2022年10期)2022-10-12
红领巾·成长(2019年3期)2019-04-16
三联生活周刊(2017年43期)2017-10-20
价值工程(2017年11期)2017-04-18
热带农业科学(2016年12期)2017-03-31
科技与创新(2016年13期)2016-07-22
江苏农业科学(2015年2期)2015-03-12
