
基于ASFV EP402R 编码蛋白CD2v 的生物信息学分析
2022-04-26 07:40顾文源赵云环左玉柱范京惠
河北农业大学学报 2022年2期
张 帅,顾文源,赵云环,刘 莹,左玉柱,范京惠
(1. 河北农业大学 动物医学院,河北 保定 071001;2. 河北省动物疫病预防控制中心,河北 石家庄 050035;3. 河北省兽医生物技术创新中心,河北 保定 071001)
非洲猪瘟(African swine fever,ASF)是由非洲猪瘟病毒(African swine fever virus,ASFV)感染引起的家猪和野猪的最具破坏性的出血性传染病之一。早在撒哈拉以南非洲国家流行,2007 年传入格鲁吉亚地区,近年来,又传播到西欧和东南亚各国,2018 年传入我国后,迅速在各地猪场蔓延,给我国养猪业造成了毁灭性的打击。根据毒力大小,ASFV可分为高毒力、中毒力、低毒力和无症状感染毒株。在非洲猪瘟的临床病例中,高致病性病毒感染最为常见,发病率和死亡率可高达100%。因此,世界动物卫生组织(OIE)将ASF 列为必须报告的动物疫病,在我国也将ASF 列为一级动物疫病[1]。
ASFV 是一种结构复杂的大型、双链、线性DNA 病毒,基因组庞大,大小约为170 ~190 kb,编码超过200 种蛋白质。由于其结构的复杂性,尽管国内外学者对其进行了大量研究,大多数蛋白质的功能仍不清楚,阻碍了疫苗的研发和致病机制及免疫逃避机制的探明,导致目前尚无商品化疫苗及特效的药物用于ASF 的有效防控[2]。在编码的200多种蛋白质中,超过50 种是构成病毒结构的主要蛋白,其中由EP402R 基因编码的CD2v 蛋白是ASFV晚期表达蛋白[3],可介导红细胞对ASFV 感染细胞的吸附作用,抑制有丝分裂原诱导的淋巴细胞增殖,延长免疫系统消灭病毒的时间,同时还是ASFV 血清型分型的关键蛋白[4],且在机体免疫调节、免疫逃逸和ASFV 病毒入侵宿主和传播扩散中发挥重要作用。目前该蛋白的结构尚未完全解析,因此深入了解CD2v 蛋白的结构及功能,将有助于ASFV 的有效防控[5]。本研究以中国首次分离且在国内流行的非洲猪瘟病毒Pig/HLJ/2018 株为研究对象,对ASFV CD2v 蛋白进行了遗传进化分析、理化性质分析、蛋白结构和亚细胞定位预测、蛋白跨膜区和信号肽分析以及B 细胞和T 细胞抗原表位分析等生物信息学分析[6],以期为ASFV 的进化分析和CD2v蛋白的功能研究、ASFV 与宿主细胞的作用机制研究以及疫苗研发提供一些基础性数据,为ASF 的有效防控奠定基础。
1 材料与方法
1.1 CD2v 蛋白遗传进化分析
通过登录NCBI 查询中国流行株Pig/HLJ/2018株(GenBank:QBH90580)ASFV EP402R 基因并选取其他国内、外具代表性的毒株20 条,获取基因组和基因型等信息(见表1),用MEGA 7.0 软件进行遗传进化树构建,描述20 种不同基因组数据之间的关系;用Megalign 软件对20 条氨基酸序列进行比较和分析。
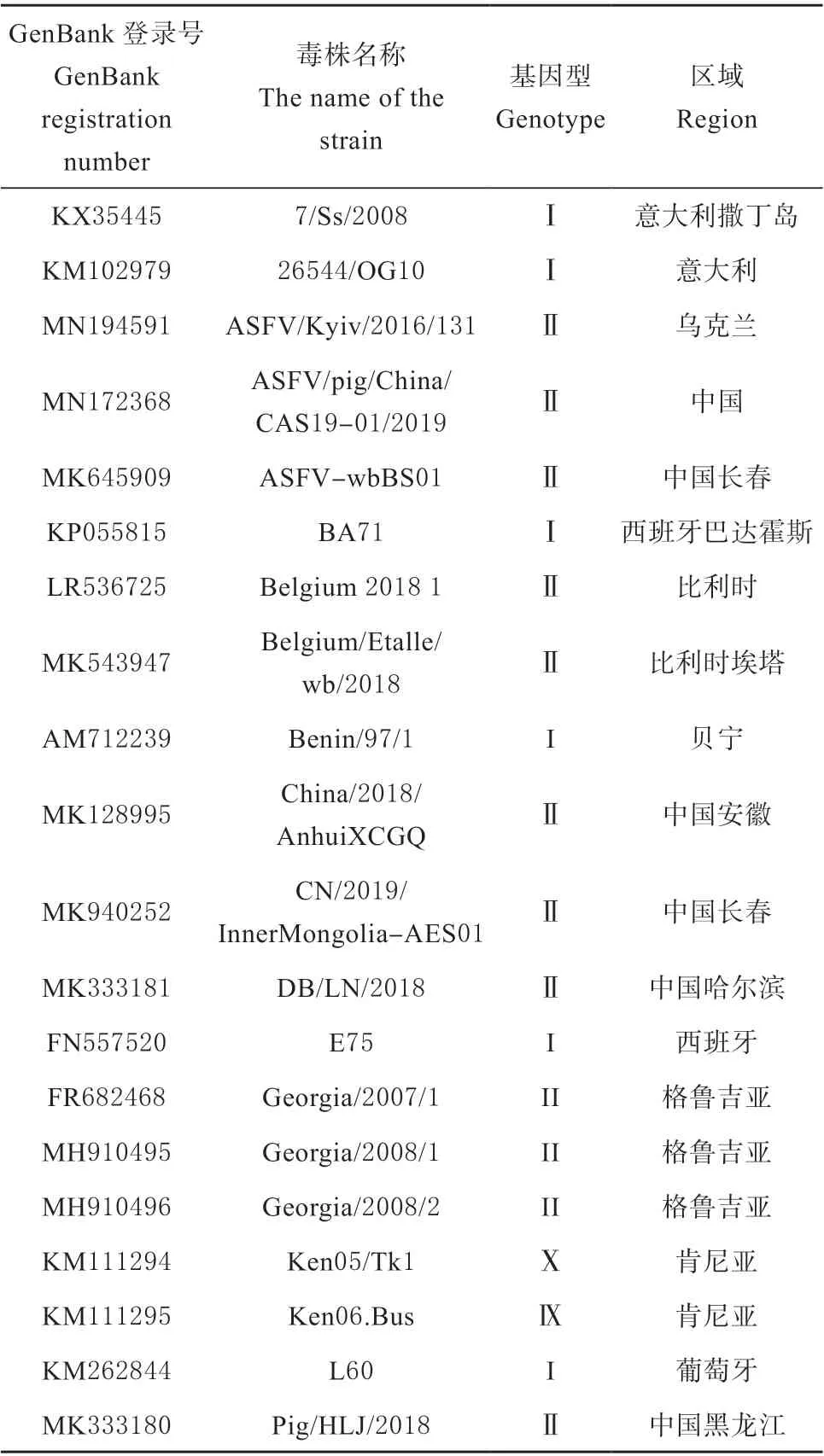
表1 20 株ASFV EP402R 基因统计信息Table 1 Statistical information of EP402R gene of 20 ASFV strains
1.2 CD2v 蛋白理化性质分析
利用ExPASy 在线分析工具ProtParam 对CD2v蛋白的分子量、等电点、氨基酸组成、原子数、疏水性等理化性质进行分析。
1.3 CD2v 蛋白结构预测
利 用 在 线 工 具GOR4、Swiss-Model、cNLS Mapper、Psort 等分析软件对CD2v 蛋白NLS 序列和二、三级结构和亚细胞定位情况进行预测[7]。
1.4 CD2v 蛋白跨膜区和信号肽预测
通过TMHMM-2.0 在线工具分析CD2v 蛋白跨膜区[8],通过SignalP-5.0 在线工具分析CD2v 蛋白信号肽[9]。
1.5 CD2v 蛋白B 细胞和T 细胞抗原表位分析
采用在线分析工具BepiPred 1.0、IEDB 分析序列B 细胞抗原表位区域[10],采用NetMHC 4.0、NetMHCpan 4.1 在线服务器分析序列T 细胞抗原表位区域[11],并结合蛋白二级结构预测中的α-螺旋、β-折叠、β-转角等不易形成抗原表位的区域,筛选出优势表位区域。
2 结果与分析
2.1 基于Pig/HLJ/2018 株ASFV CD2v 蛋白序列遗传进化分析结果
2.1.1 基于ASFV CD2v 蛋白基因EP402R 的进化树 通 过 对Pig/HLJ/2018 株ASFV 和 从NCBI 查询筛选出的ASFV CD2v 蛋白基因EP402R 序列组成20 条序列构建进化树(图1)结果显示,Pig/HLJ/2018 株 与 格 鲁 吉 亚Georgia 2007、Georgia 2008 以及其他中国毒株处于同一分支,基因组分型同为Ⅱ型,同时显示当前格鲁吉亚地区、中国、东南亚地区流行毒株均为基因Ⅱ型;西欧地区流行毒株主要为基因Ⅰ型;非洲地区当前流行毒株多为Ⅸ型和Ⅹ型,从进化关系来看,Ⅸ型和Ⅹ型具有很高的相似性[12]。

图1 20 株ASFV EP402R 基因进化树Fig.1 20 ASFV EP402R gene evolutionary tree
2.1.2 ASFV CD2v 蛋白氨基酸序列分析 通过Megalign 软件对20 条氨基酸序列进行比较和分析后结果(见图2)显示,不同基因型之间氨基酸序列存在着不同程度的突变、插入与缺失,同一基因型间氨基酸序列的一致性较高,Pig/HLJ/2018 株与格鲁吉亚Georgia 2007、Georgia 2008 以及中国AnhuiXCGQ 等基因Ⅱ型毒株CD2v 蛋白氨基酸序列一致性高达100%,与其他基因型的氨基酸一致性在76.8%~81.7%之间。同时还可以看出,CD2v 蛋白氨基酸序列末端(C 端)出现了KPCPPP 的重复序列。
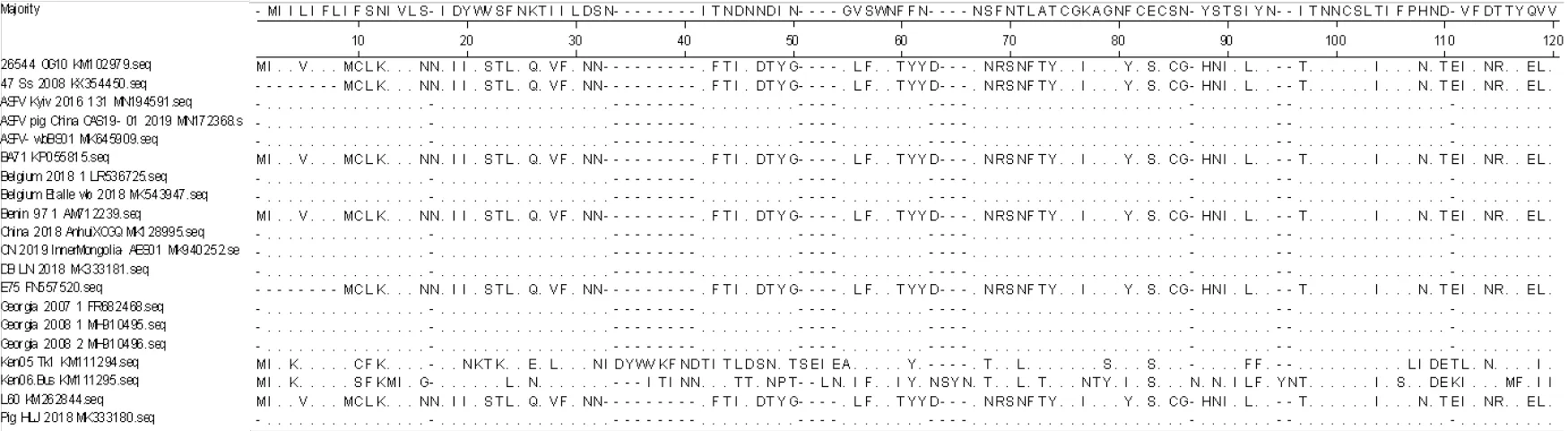
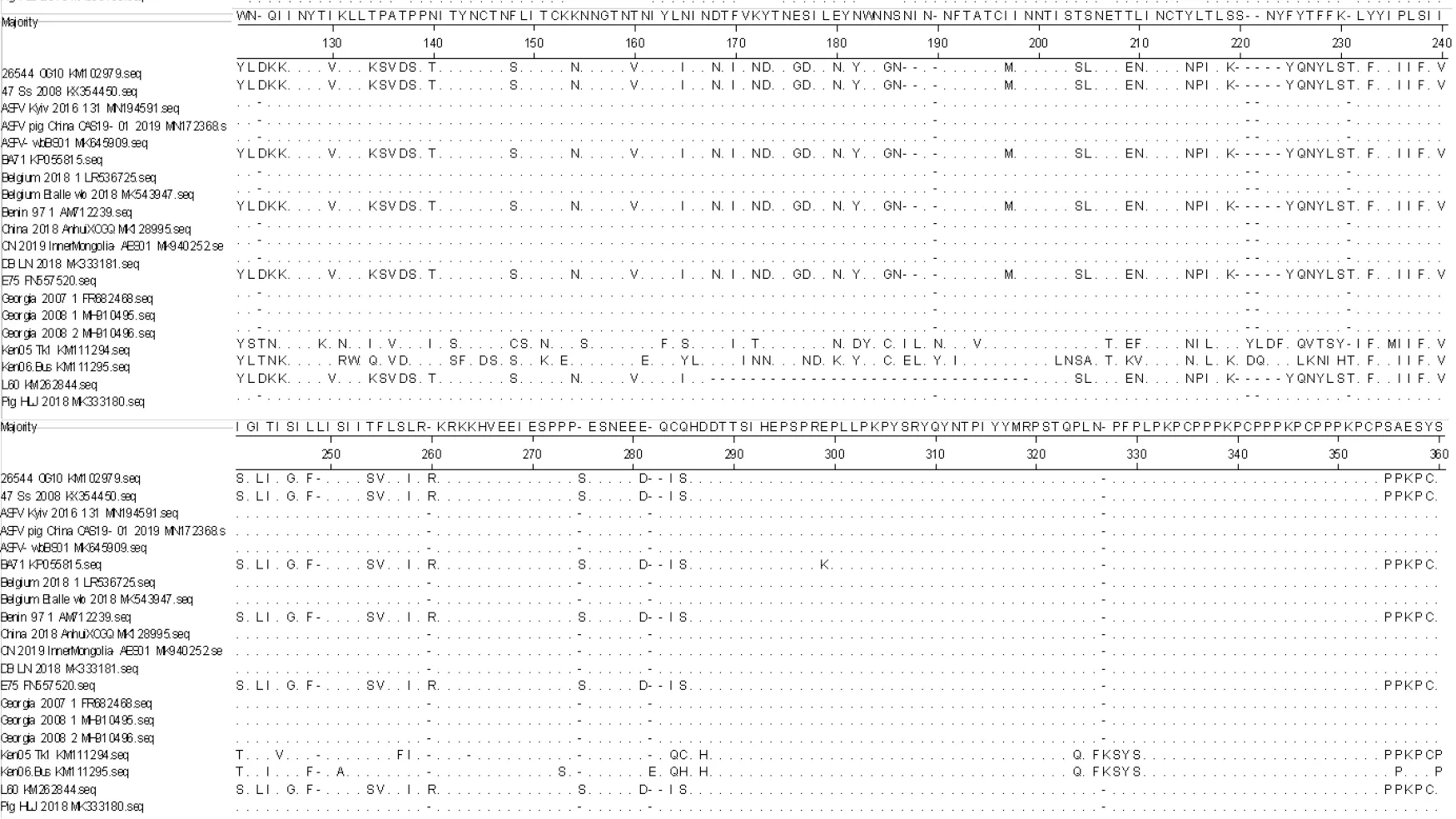
图2 ASFV CD2v 氨基酸序列分析结果Fig.2 ASFV CD2v amino acid sequence analysis results
2.2 CD2v 蛋白理化性质分析结果
根据ExPASy 在线工具中的ProtParam 分析结果显示:CD2v 蛋白共360aa,分子量为41.008 89 kD,其中天冬氨酸(12.5%)、异亮氨酸(12.5%)、脯氨酸(11.4%)和苏氨酸(9.7%)占比最高,其他氨基酸占比从0.6% ~8.9% 不等;理论等电点为6.21,带负电荷的氨基酸总数为24,带正电荷的氨基酸总数为22;原子总数为5 749,分子式为C1868H2858N460O548S15;预测280 nm 消光系数为51 800 M-1cm-1~52 550 M-1cm-1;在哺乳动物体外网织红细胞中的半衰期为30 h,在酵母体内半衰期大于20 h,在大肠杆菌体内半衰期大于10 h;不稳定指数为47.78,脂肪指数为88.81,亲水性总平均值为-0.209,说明该蛋白属于亲水性蛋白。
2.3 CD2v 蛋白结构和亚细胞定位预测
Pig/HLJ/2018 株ASFV CD2v 蛋白一级结构表明该基因全长1 080 bp,G+C 含量为27.88%,A+T含量为72.12%;表达的蛋白为41.008 89 kD,通过在线工具GOR4 预测的CD2v 蛋白二级结构结果(图3)显示,α-螺旋占5.56%,扩展链占31.11%、无规卷曲占63.33%;将CD2v 蛋白序列提交在线工具Swiss-Model 预测该蛋白的三级结构模型(见图4),证明了该蛋白主要以无规卷曲为主。
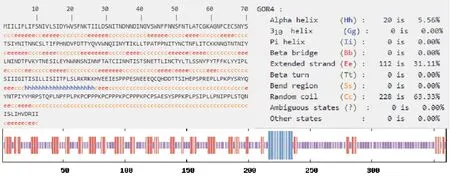
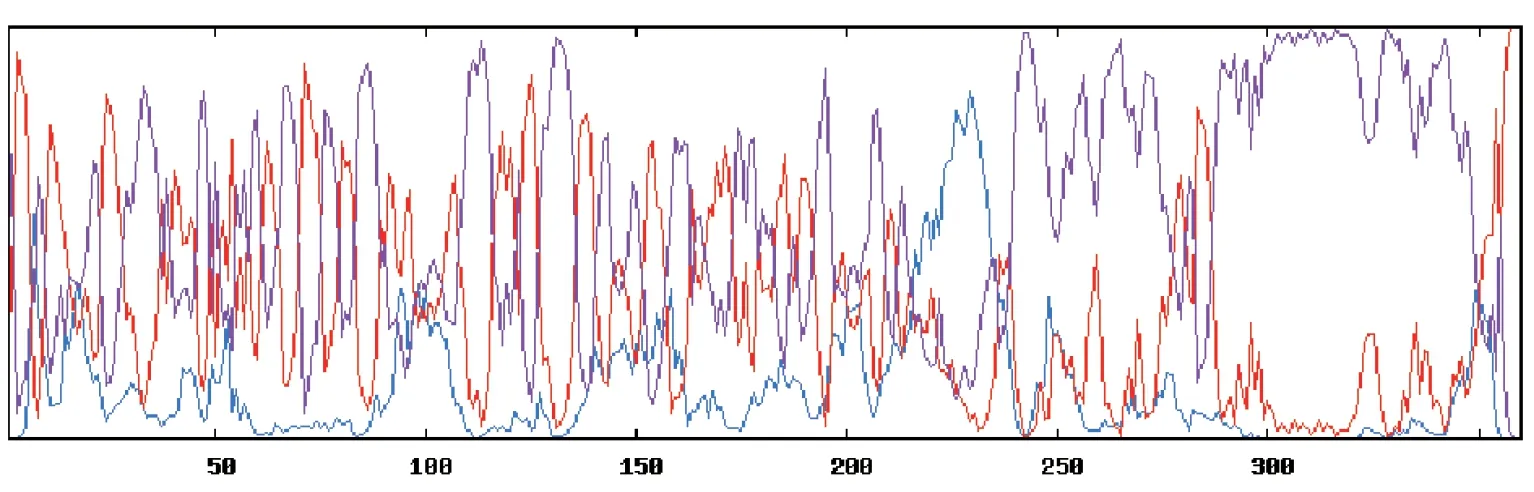
图3 CD2v 蛋白二级结构预测结果Fig.3 Rediction results of secondary structure of CD2v protein
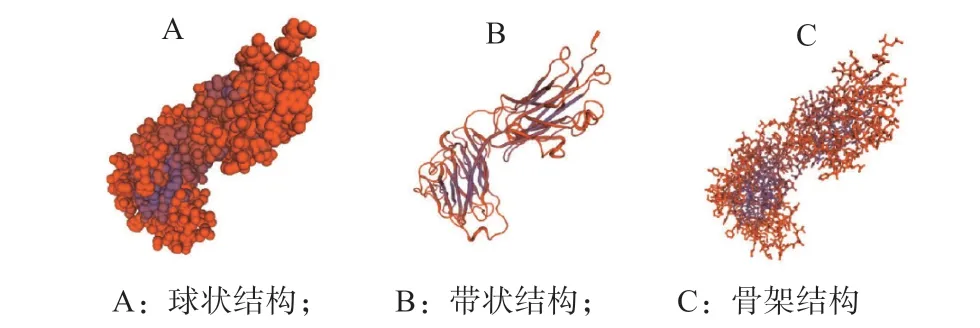
图4 CD2v 蛋白三级结构预测结果Fig.4 Prediction of tertiary structure of CD2v protein
根据在线工具cNLS Mapper 和Psort 预测结果显示,该蛋白可能包括一段NLS 单分型序列(表2),无双分型序列,其cut-off 分数为7.5;CD2v 蛋白可能定位于高尔基体的概率为22.2%,定位于内质网和内膜的概率均为11.1%。

表2 ASFV CD2v 蛋白序列单分型NLS 预测Table 2 Single NLS prediction of ASFV CD2v protein sequence
2.4 CD2v 蛋白跨膜区和信号肽分析结果
根据TMHMM-2.0 在线工具分析结果显示,CD2v 蛋白存在一个跨膜区(见图5-A),信号肽分析结果的可能性为0.925,说明存在信号肽(见图5-B)。
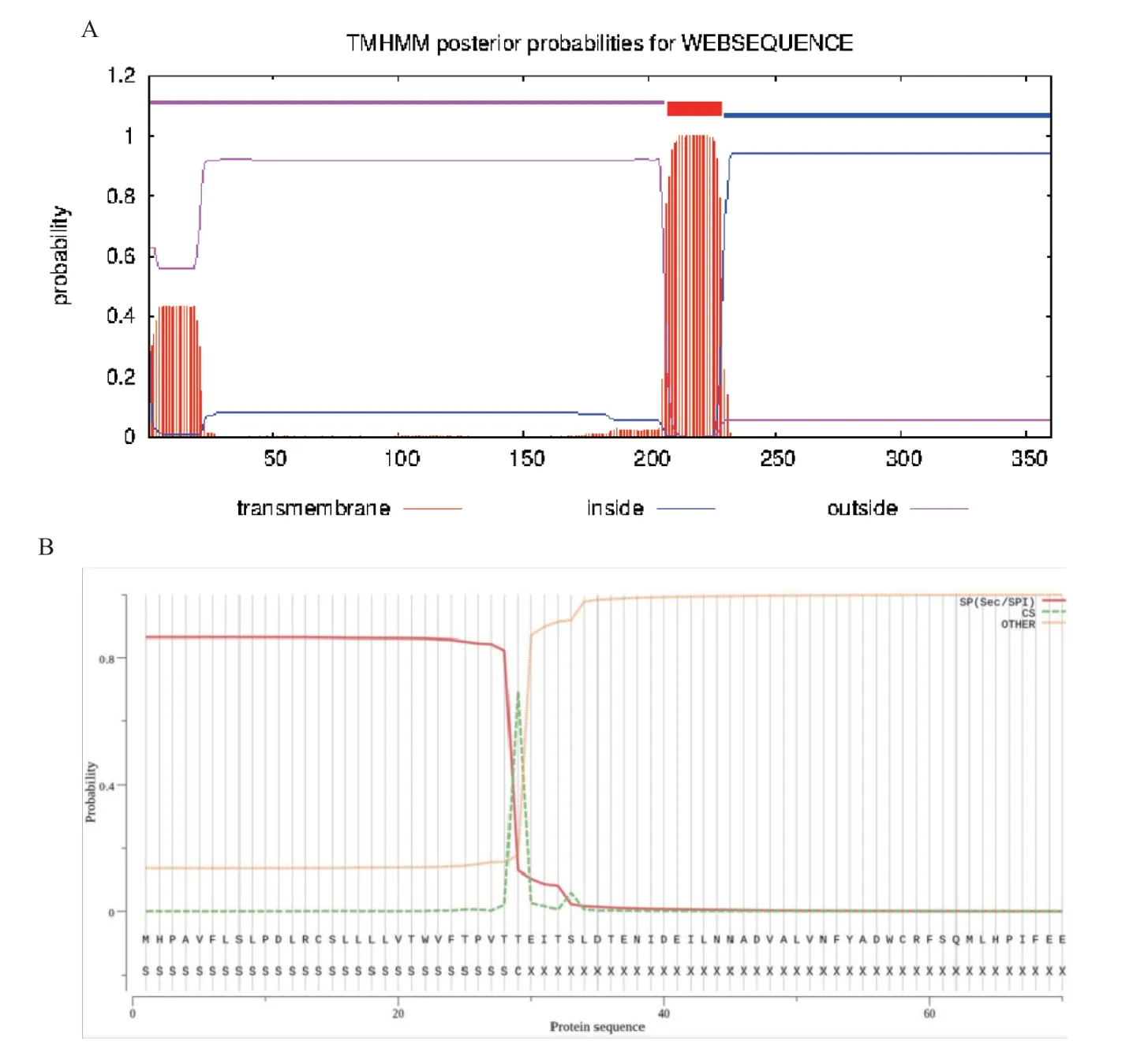
图5 CD2v 蛋白跨膜区和信号肽预测结果Fig.5 Prediction results of CD2v protein transmembrane region and signal peptide
2.5 CD2v 蛋白B 细胞和T 细胞抗原表位分析结果
根据在线分析工具BepiPred 1.0、IEDB 分析序列B 细胞抗原表位区域结果显示,存在9 个B 细胞抗原表位;根据NetMHC 4.0、NetMHCpan 4.1 在线服务器分析序列T 细胞抗原表位区域结果显示,CD2v 蛋白存在14 个T 细胞抗原表位;结合2.3 中预测的二级结构,去掉其中的α-螺旋等筛选出4 个优势B 细胞抗原表位和5 个优势T 细胞抗原表位(见表3)。
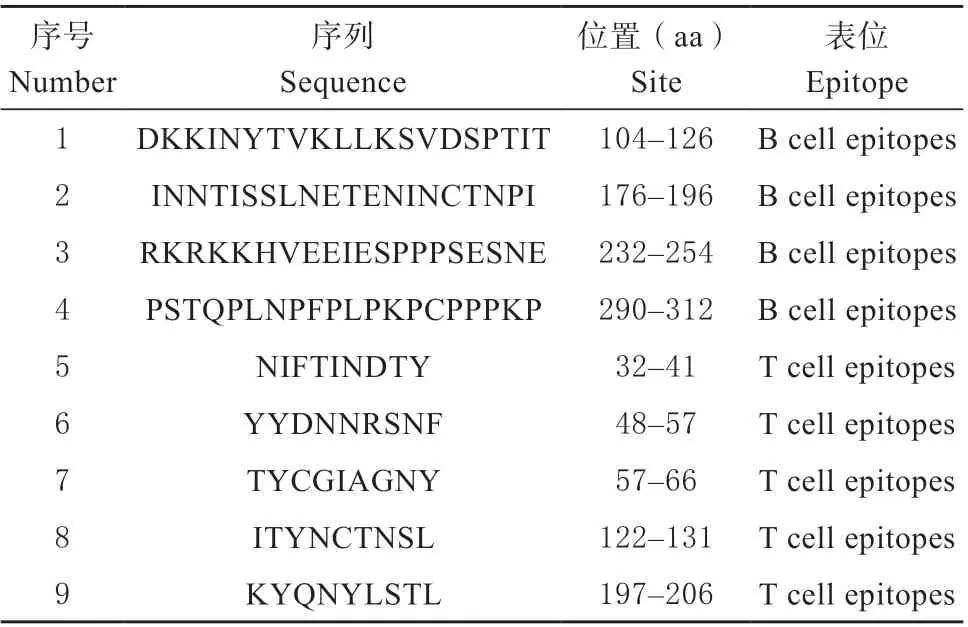
表3 CD2v 蛋白优势B、T 细胞抗原表位分析统计结果Table 3 Statistical results of CD2v protein dominant B cell and T cell epitope analysis
3 讨论与结论
ASFV 是一种基因组庞大而复杂的DNA 病毒,可导致猪的急性死亡,给全球养猪业造成了巨大的经济损失。我国于2018 年在辽宁首次爆发ASF[13],之后迅速蔓延至多个省市和地区,并呈区域性流行,严重威胁着我国养猪业的健康发展。尽管政府部门按照一类传染病的防控措施,对疫区及受威胁地区进行了严格的管控[14],但防控形势依然严峻,急需安全有效的疫苗用于本病的防控。然而,由于对ASFV 与宿主之间的作用机制、病毒在宿主体内的复制调控机制尚不清楚,对与病毒免疫逃避相关蛋白的研究相对较少,目前尚无商品化的非洲猪瘟疫苗。因此,深入研究病毒蛋白的结构和功能,将有助于病毒致病机制的阐明以及非洲猪瘟疫苗的研发。
3.1 ASFV CD2v 蛋白的遗传变异分析
CD2v 蛋白位于ASFV 的外层囊膜,可诱导机体产生中和抗体,是病毒毒力及免疫逃避的相关蛋白之一[15-16]。本研究对ASFV CD2v 蛋白氨基酸序列分析结果显示,不同基因型之间氨基酸序列存在较大差异,以中国流行株Pig/HLJ/2018 株ASFV CD2v 蛋白氨基酸序列为例,在C 末端出现了连续的重复序列(KPCPPP),研究表明,该重复序列是一种遗传标记或抗原表位[17-18],Yang 等[18]确认了该重复序列(KPCPPP)为细胞穿透肽(CPP),CPP 是多种多肽家族,通常由5 ~30 个氨基酸组成,可通过能量依赖或能量独立机制通过组织和细胞膜,不与特定受体相互作用[19],它可以携带EGFP 通过网格蛋白和微胞饮介导的内吞进入CHO 细胞,这为ASFV 颗粒进入细胞的方式提供了新的见解,同时也提示ASFV研究可以发展成为药物传递的工具。另外,连续的脯氨酸序列与宿主SH3 结构域蛋白SH3P7/mAbp1 结合,并在囊泡运输中发挥作用[20]。对ASFV CD2v 蛋白遗传进化与氨基酸序列分析表明该蛋白可作为ASF 治疗药物和预防疫苗的研究靶点,为ASF 的有效防控奠定基础。
3.2 ASFV CD2v 蛋白结构和抗原表位
对ASFV CD2v 蛋白的结构分析结果显示,CD2v 是一种跨膜蛋白,蛋白二级结构显示α-螺旋结构占5.56%,存在跨膜区和信号肽,具有由2 个免疫球蛋白样结构域组成的细胞外N 端区域和胞质C 端区域,与Rodríguez 等[21]的研究结果相一致。Sereda 等[22]研究表明CD2v 蛋白能介导红细胞对ASFV 感染细胞的吸附作用,并指出ASF 保护性免疫可能是血清型特异性的表现,血液吸附(HAI)抑制同一血清型内的病毒互相交叉保护,而同一血清型外的病毒则没有交叉保护;同时Malogolovkin 等[23]根据CD2v 蛋白HAI 特性进行了血清学分型,发现同一基因型下的毒株表现出不同的血清型分型,这可能更好地解释ASFV 分离株之间的同源交叉保护问题和影响疾病出现的病毒决定因素,表明ASFV CD2v 蛋白的血清分型对疫苗的设计和开发非常重要。T 细胞介导的细胞免疫和B 细胞介导的体液免疫,在机体抵抗病原微生物感染的过程中,发挥着重要作用,B、T 细胞的优势抗原表位常用于靶向药物或表位疫苗的研制,Burmakina 等[24]在免疫动物攻毒后7 ~10 d,观察到T 细胞对病毒的强大反应,并鉴定出了在ASF 保护性免疫环境中诱导T 细胞应答的T 细胞表位区域,这表明ASFV 的T 细胞表位对保护性宿主反应具有重要意义。同时,Burmakina等[24]提示T 细胞表位对给定的病毒血清型也有特异性,T 细胞宿主反应可能与观察到的血清型特异性保护有关。本研究通过在线工具筛选出4 个优势B 细胞抗原表位和5 个优势T 细胞抗原表位,可作为靶向药物、表位疫苗筛选的对象[6],有助于ASFV 亚单位疫苗的设计和开发。
通过对ASFV 的CD2v 蛋白及其基因进行生物信息学分析,深入解析了中国流行毒株Pig/HLJ/2018株CD2v 蛋白基因的遗传进化情况以及CD2v 蛋白的结构与功能关系,为进一步阐明病毒-宿主相互作用机制以及ASFV 疫苗的研发奠定了基础。
猜你喜欢
传染病信息(2022年4期)2022-09-17
中国高原医学与生物学杂志(2022年3期)2022-06-22
福建农业学报(2022年3期)2022-05-24
智慧健康(2021年2期)2021-03-15
昆明医科大学学报(2021年1期)2021-02-07
温州医科大学学报(2019年4期)2019-04-28
数码世界(2018年1期)2018-12-23
生物学教学(2018年10期)2018-11-29
江苏农业科学(2017年23期)2018-01-29
今日健康(2016年7期)2017-04-12
