
鼠茅草套种对不同林龄油茶根际土真菌群落的影响
2022-05-18 22:59王晓艳
福建农业科技 2022年3期
王晓艳

摘 要:為探究林下套种鼠茅草对不同林龄油茶根际土真菌群落的影响,在覆盖鼠茅草条件和无鼠茅草覆盖条件下分别取3~5年生、10~15年生、50年生及以上的油茶根际土进行高通量测序。结果发现:覆草条件下的根际土微生物多样性更丰富,且50年生及以上林龄的根际土真菌群落多样性指数更高。在科水平上优势菌三形菌Trimorphomycetaceae在10~15年林龄的相对丰度最高;在属水平上优势菌三形菌属Saitozyma在10~15年林龄的相对丰度最高。在油茶根际土真菌群落及林龄相关性分析中陆生壶菌科Terramycetaceae相对丰度与3~5年林龄阶段正相关性最强;锥梗菌科Dissoconiaceae相对丰度与
10~15年林龄阶段正相关性最强、黑球腔菌科Melanommataceae相对丰度与50年及以上林龄阶段正相关性最强。腐生菌的功能作用在覆草条件下更强,表明种植鼠茅草能更好地促进土壤养分的转化,从而为油茶的生长提供营养。总而言之,本研究证明了鼠茅草的种植能增加油茶根际土真菌微生物多样性、改善真菌群落的功能、促进油茶的生长发育,且在10~15年生阶段油茶效益最高,为油茶产业的发展提供参考。
关键词:油茶;鼠茅草;林龄;真菌;高通量测序
中图分类号:S 154.3 文献标志码:A 文章编号:0253-2301(2022)03-0032-08
DOI: 10.13651/j.cnki.fjnykj.2022.03.006
Effects of Interplanting with Vulpia myuros on the Fungal Community in Rhizosphere Soilof Camellia oleifera at Different Stand Ages
WANG Xiao-yan
(Fujian Sanming Bureau of Forestry, Sanming, Fujian 365000, China)
Abstract: In order to investigate the effects of interplanting with Vulpia myuros on the fungal community in rhizosphere soil of Camellia oleifera at different stand ages, the rhizosphere soils of Camellia oleifera aged 3-5 years, 10-15 years and 50 years or more were sequenced by using the high-throughput sequencing under or without the condition of covering with Vulpia myuros. The results showed that the microbial diversity of rhizosphere soil was more abundant under the condition of straw mulching, and the diversity index of the fungal community in rhizosphere soil with the stand age of 50 years or more was higher. At the family level, the relative abundance of dominant bacteria Trimorphomycetaceae was the highest at the stand age of 10 to 15 years, while at the genus level, the relative abundance of dominant bacteria Saitozyma was the highest at the stand age of 10 to 15 years. In the correlation analysis of the fungi community in rhizosphere soil of Camellia oleifera and the stand age, the relative abundance of Terramycetaceae had the strongest positive correlation with the stand age of 3 to 5 years. The relative abundance of Dissoconiaceae had the strongest positive correlation with the stand age of 10 to 15 years, while the relative abundance of Melanommataceae had the strongest positive correlation with the stand age of 50 years or more. The functional effect of saprophytic bacteria was stronger under the condition of straw mulching, which indicated that the cultivation of Vulpia myuros could better promote the transformation of soil nutrients, thus to provide the nutrition for the growth of Camellia oleifera. In conclusion, this study proved that the cultivation of Vulpia myuros could increase the microbial diversity of fungi in the rhizosphere soil of Camellia oleifera, improve the function of fungal community, and promote the growth and development of Camellia oleifera, and the benefit of Camellia oleifera was the highest when planted for 10 to 15 years, which would provide reference for the development of Camellia oleifera industry.
Key words: Camellia oleifera; Vulpia myuros; Stand age; Fungi; High-throughput sequencing
油茶Camellia oleifera Abel.,俗称大果油茶,山茶科山茶属。在中国主要分布于黄河以南,海拔150~2 050 m的山坡灌丛、常绿林区。油茶其种子可榨油,其叶可作肥料,此外还具有一定的药用价值和工业用途[1]。为了追求高产,油茶林的施肥过量导致土壤板结等问题;有研究证明生态栽培可减少水土流失,保持土壤养分和生物多样性[2-4],因此生态栽培成为目前解决油茶林地土壤问题的主要方法。鼠茅草Vulpia myuros为禾本科植物,具有易种植、耐阴性强、分檗能力强等优点[5],有研究表明套种鼠茅草可以有效抑制杂草、提高土壤孔隙度、降低土壤容重、减轻水土流失等[6-7],因此可选用油茶林套种鼠茅草来实现生态栽培。
根际微生物群落在植物的生长发育方面起着关键作用,有益微生物可通过各种直接和间接的方式促进生长,包括增加养分供应、预防疾病、诱导抗性等作用[8-9];反之,有害微生物中的致病菌累积到一定程度会致使植物死亡[10]。如何合理调节根际微生物菌落,增加有益微生物的多样性,达到高产和稳产的效果,是高效生态栽培的核心。
真菌群落可促进植物对氮、磷、钾等元素的吸收,提高铁元素的有效性[11]。其群落的组成与功能受植物年龄的影响较大,有研究发现,在粗枝云杉人工林土壤中,真菌的相对丰度随着林龄的增加而增加
[12];不同林龄条件下,银杏内生真菌群落的结构有显著性差异[13];肖龙敏等[14]发现宁夏枸杞根际土中真菌群落的多样性与林龄呈负相关。高通量测序微生物技术已在其他植物研究中有效成熟的运用:有研究通过高通量测序结果发现不同的耕作方式对水稻根际土真菌群落的多样性和真菌群落的结构组成有影响[15];高通量测序结果表明患病的三七中,球囊菌门的丰度高于健康三七[16];在青稞中,不同种植模式下真菌的优势种群具有显著差异[17];而在油茶的研究中,测序主要集中在对不同油茶品种[18]或病毒的转录组测序[19-20]方面,对不同林龄油茶根际土真菌群落的探究尚不明确,因此本研究利用高通量测序分析方法来探究油茶根际土真菌群落与覆草条件和年龄条件之间的关系及差异变化,以期填补油茶根际土真菌群落研究的空白,为油茶的种植及产业发展提供相关参考数据。
1 材料与方法
1.1 样地概况
试验样地位于福建省三明市清流县赖坊镇山珍源油茶开发有限公司山场(25°56′13″N,116°53′23″E),气候温暖湿润,四季分明,平均温度15.6~19.6℃;年平均降水量1 510~1 840 mm,年均降雨日174 d,年日照时数1 878 h,无霜期303 d;样地土壤为山地红壤,其中种植有鼠茅草,覆盖度为83%。
1.2 采集油茶根际土样品
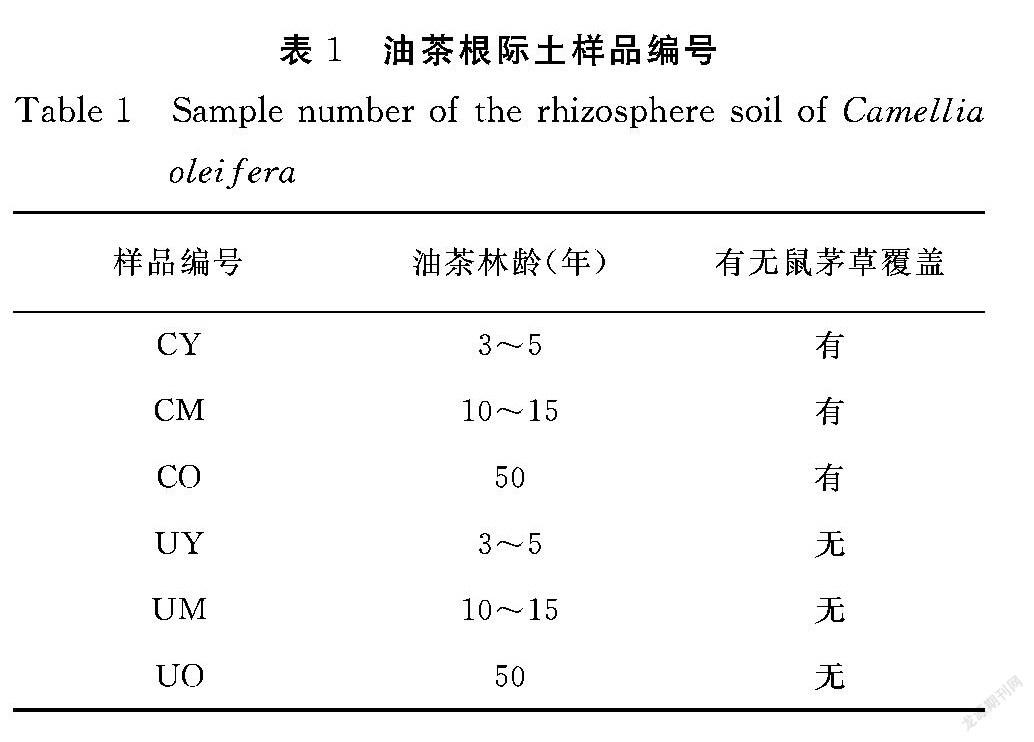
于试验样地分别采集3~5年生、10~15年生、50年及以上3个年龄段有鼠茅草覆盖和无鼠茅草覆盖条件下的油茶根际土,每个样品3个重复;样品编号见表1。采样时取油茶30 cm深处的根际土壤于50 mL无菌离心管,立即放入干冰储存,运输至实验室后放置于-80℃冰箱保存。
1.3 处理油茶根际土样品
在超净台内抖落附着在根系的非根际土,镊子置于酒精灯上消毒,并用无菌水冷却镊子至常温后,用镊子将根系样品转移至新的50 mL离心管并加入20 mL 1×PBS,于28℃摇床振荡20 min。用无菌镊子去除离心管中的根系,离心(4 000 r·min-1、 4℃、 20 min), 弃上清, 储存于-80℃。
1.4 油茶根际土ITS区测序及数据处理
1.4.1 基因组提取 用DNA Kit(Omega Bio-tek,Norcross,GA,U.S.)试剂盒提取基因组DNA,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA。
1.4.2 樣品质量检测与扩增 Nanodrop检测样品质量,取30 ng进行PCR扩增。按MiSeqPE300测序区域,合成带有barcode的特异引物,F′引物:CTTGGTCATTTAGAGGAAGTAA,R′引物:TGCGTTCTTCATCGATGC。PCR反应体系为:2×Taq Plus Master Mix 12.5 μL,F′引物1 μL,R′引物1 μL,DNA 1 μL,BSA(2 ng·μL-1) 3 μL,ddH2O 6.5 μL,共25 μL。PCR扩增程序:预热94℃ 5 min,94℃变性30 s,55℃退火30 s,72℃延伸1 min,变性-退火-延伸设置34个循环,72°保持7 min,4℃保存。PCR 产物使用 1%琼脂糖凝胶电泳检测扩增目的条带大小,并用AgencourtAMPure XP 核酸纯化试剂盒[贝克曼库尔特商贸(中国)有限公司]纯化。
1.4.3 文库构建与测序 构建Miseq文库并上机测序,为使分析结果更加准确、可靠,测序原始数据下机后,首先进行数据质控,通过序列拼接、过滤和去嵌合体后得到优化序列,然后进行OTU聚类。为了得到每个OTU对应的物种分类信息,采用blast或RDP Classifier、uclust consensus taxonomy assigner等方法对OTU代表序列进行比对分析,并在各个水平(门、纲、目、科属、种)注释其群落的物种信息。用uparse算法进行OTUs聚类分析,R语言工具统计做veen图以分析样品相似性、物种稀释曲线及物种组成分析。利用qiime软件运算Chao1、observed_species、PD_whole_tree、Shannon等alpha多样性指数,用SPSS软件进行显著性差异分析(P<0.05)。功能基因使用Blast 在NCBI NT 数据库(https://ftp.ncbi.nih.gov/blast/db)进行比对。本试验样品送至奥维森基因科技有限公司测序分析。2 结果与分析
2.1 不同种植条件下油茶根际土壤真菌OTUs聚类分析
图1a显示,随着样品数量的增加,物种稀释曲线趋向平坦,说明本次测序数据量合理,可以反映样本中绝大多数的微生物信息。Miseq测序得到Pair-End(PE)双端序列数据在去除barcode和primer并拼接后得到raw_tags,raw_tags经进一步去除嵌合体、短序列后得到优质序列clean_tags。将clean tags用uparse聚类法进行聚类,共产生4 699个OTUs,经过抽平处理后剩余4 621个OTUs。其中覆草条件的样品有1 386个特有的OUTs,无草条件的样品有1 210个特有的OTUs,两类样品共有OTUs数量为2 025个(图1b)。
2.2 不同种植条件下油茶根际土真菌多样性分析
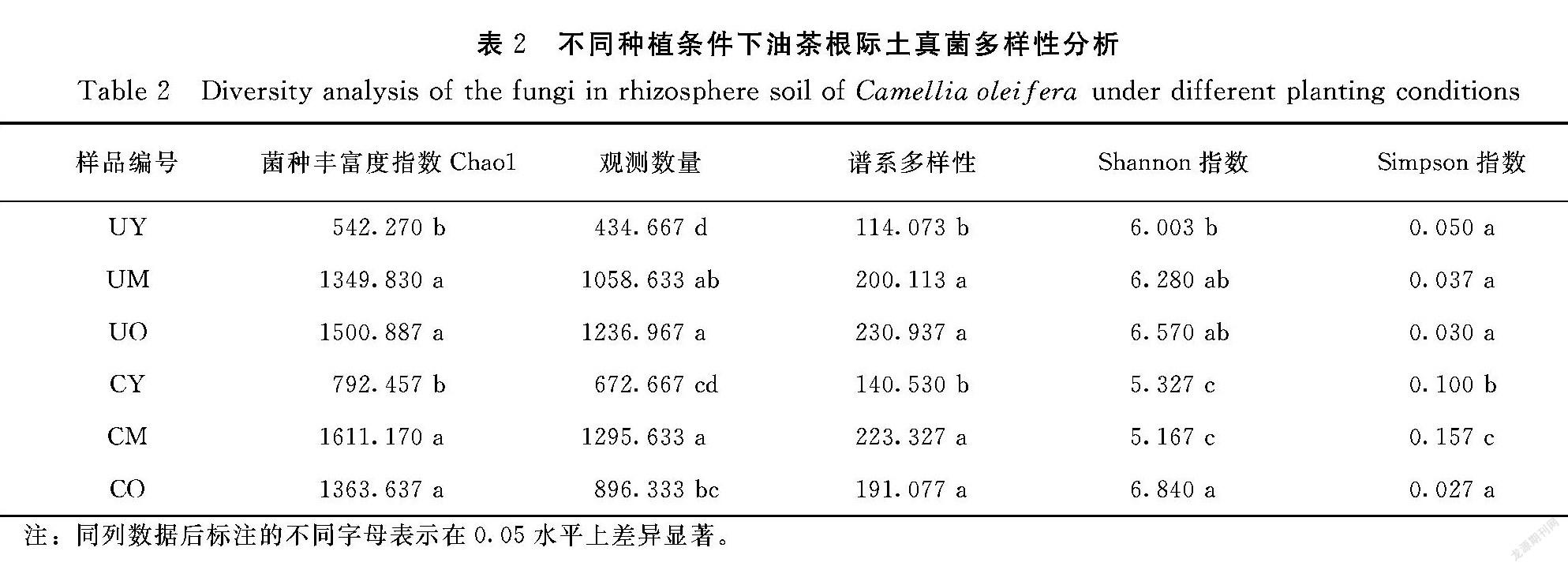
将样品多样性指数的重复值取平均值后进行显著性差异比较(表2),Chao1指数在无草条件的3组样品中由高到低分别为UO(1500.887)、UM(1349.830)、UY(542.270),在覆草条件的3组样品中由高到低分别为CM(1611.170)、CO(1363.637)、CY(792.457)。UY、CY与其他4组样品有显著性差异(P<0.05)。observed_species指数中UY与其他4组样品有显著性差异(P<0.05),CM为最高值1295.633,UY为最低值434.667。UY、CY的PD_whole_tree指数与UM、UO、CM、CO有显著性差异(P<0.05),无草条件的样品中UO(230.937)>UM(200.113)>UY(114.073),覆草条件的样品中CM(223.327)>CO(191.077)>CY(140.530)。整体来看无草条件组的Shannon指数高于覆草条件的Shannon指数,其中最高值为CO(6.840),最低值为CM(5.167),CM和CY与其他4组样品有显著差异(P<0.05)。CM的Simpson指数为最低值(0.843)显著低于其他样品;其次为CY(0.900)显著低于UY、UM、UO和CO,CO为最高值(0.973)。
2.3 不同种植条件下油茶根际土真菌物种组成分析
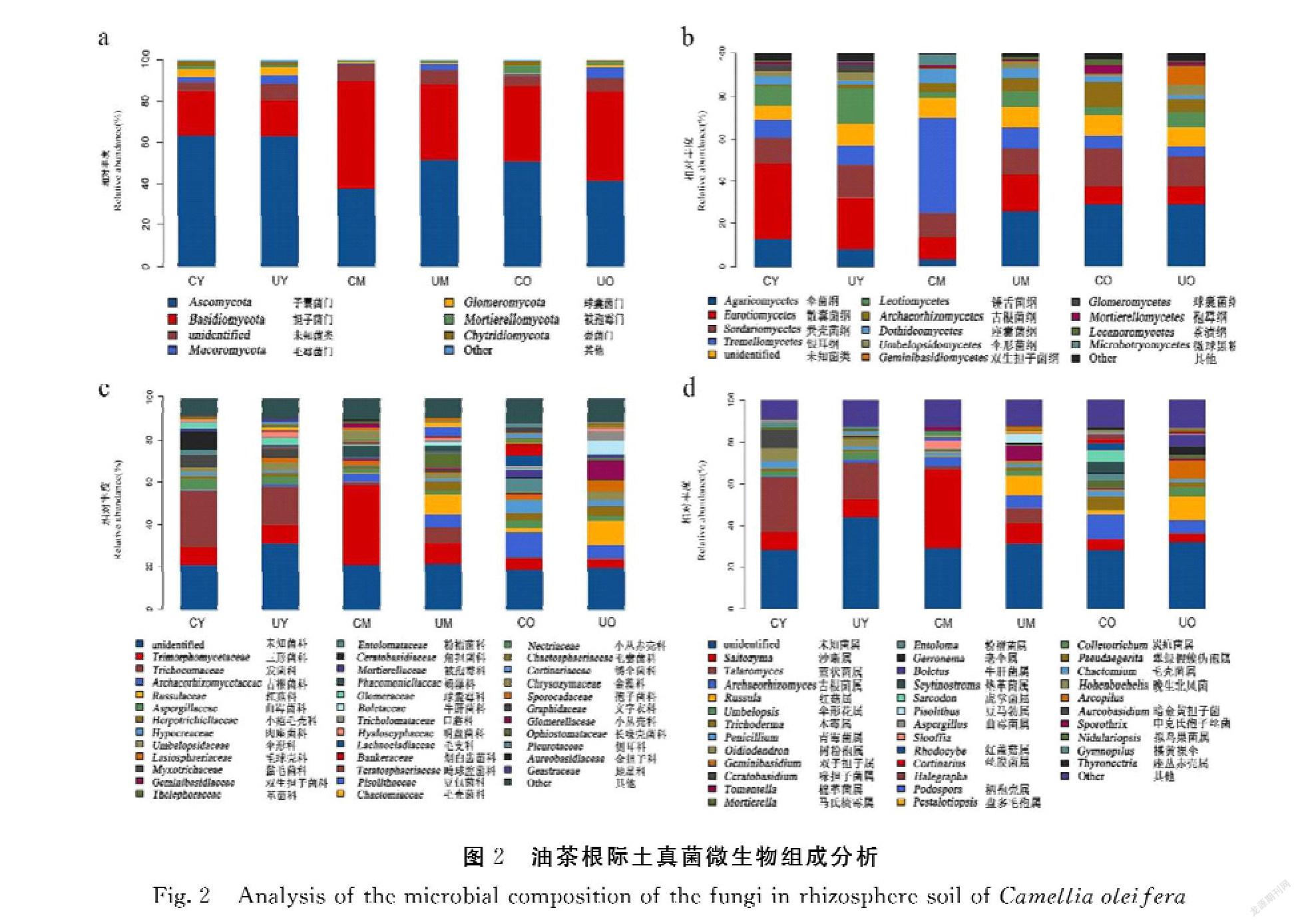
在门水平上(图2a),所有样品主要由子囊菌门Ascomycota和担子菌门Basidiomycota组成(图2a),覆草条件的3组样品中子囊菌门Ascomycota的相对丰度CY>CO>CM,无草条件的3组样品中子囊菌门Ascomycota的相对丰度UY>UM>UO;覆草条件的样品担子菌门Basidiomycota的相对丰度由高到低分别为CM、CO、CY,无草条件的样品中担子菌门Basidiomycota的相对丰度UO>UM>UY。在纲水平上(图2b),除CM外其他样品主要由伞菌纲Agaricomycetes、散囊菌纲Eurotiomycetes、粪壳菌纲Sordariomycetes组成,而CM的优势菌纲是银耳纲Tremellomycetes相对丰度最高。在覆草条件的样品中伞菌纲Agaricomycetes相对丰度由高到低为CO、CY、CM,在无草条件的样品中伞菌纲Agaricomycetes相对丰度随着年龄的增加而增加;散囊菌纲Eurotiomycetes在覆草条件的样品中相对丰度由高到低为CY、CM、CO,在无草条件的样品中相对丰度随着年龄的增加而降低;粪壳菌纲Sordariomycetes在覆草条件的样品中相对丰度由高到低为CO、CY、CM,在无草条件的样品中相对丰度由高到低为UY、UO、UM。在科水平上(图2c),覆草条件的样品中三形菌科Trimorphomycetaceae相对丰度由高到低为CM、CY、CO,无草条件的样品中由高到低为UM、UY、UO; CO样品中古根菌科Archaeorhizomycetaceae相对丰度最高,其次为肉座菌科Hypocreaceae;UO主要由红菇科Russulaceae、双生担子科Geminibasidiaceae组成;CY、UY样品中相对丰度最高的为毛刷囊菌科Trichocomaceae,为优势菌科。在属水平上(图2d),三形菌属Saitozyma在覆草条件的样品中相对丰度CM>CY>CO,在无草条件的样品中相对丰度UM>UY>UO;CO样品中相对丰度最高的为古根菌属Archaeorhizomyces;UO样品中红菇属Russula的相对丰度最高,其次为双生担子属Geminibasidium;CY、UY样品中相对丰度最高的为篮状菌属Talaromyces,为优势菌属。
2.4 林龄与不同种植条件下油茶根际土真菌的相关性
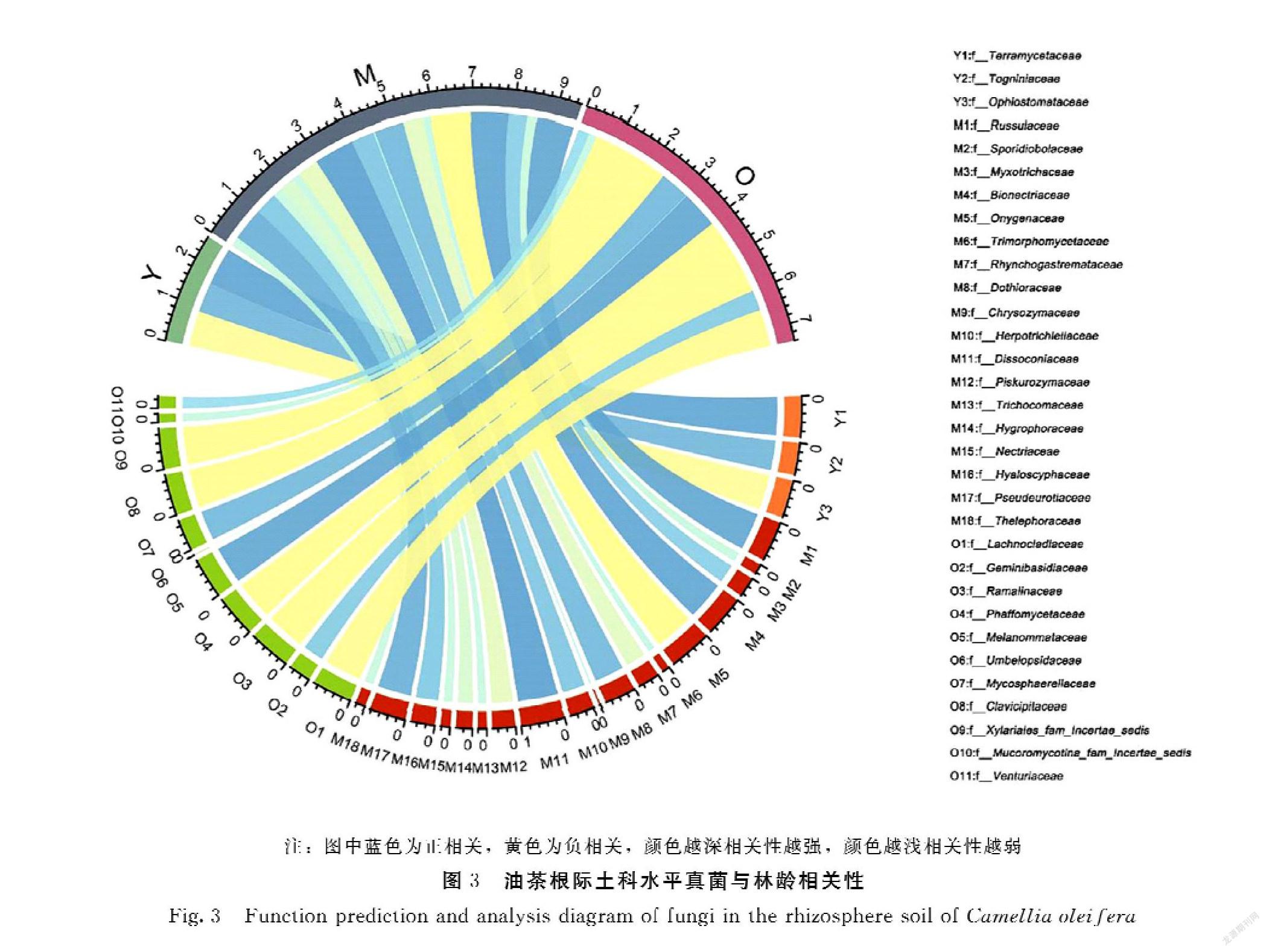
每个林龄段油茶根际土真菌在科水平上的物种不尽相同,因此将不同林龄与其下科水平真菌相对丰度做差异性分析和相关性分析。数据分析过程中,将3~5年林龄定义为Y,10~15年林龄定义为M,50年以上林龄定义为O。以上不同林龄为变量,科水平真菌群落的相对丰度为因变量,选取P<0.01水平下的相关性系数r做弦图。结果图3显示:在3~5年林齡阶段,陆生壶菌科Terramycetaceae、Togniniaceae(Y2)、长喙壳菌科Ophiostomataceae(Y3)相对丰度均与之呈极显著相关;其中,陆生壶菌科Terramycetaceae(Y1)相对丰度正相关程度最大,长喙壳菌科Ophiostomataceae(Y3)相对丰度负相关程度最大。在10~15年林龄阶段,红菇科Russulaceae(M1)、Sporidiobolaceae(M2)、黏毛菌科Myxotrichaceae(M3)、Bionectriaceae(M4)、Onygenaceae(M5)、三形菌科Trimorphomycetaceae(M6)、Rhynchogastremataceae(M7)、Dothioraceae(M8)、金藻科Chrysozymaceae(M9)、小疱毛壳科Herpotrichiellaceae(M10)、Dissoconiaceae(M11)、Piskurozymaceae(M12)、发菌科Trichocomaceae(M13)、Hygrophoraceae(M14)、小丛赤壳科Nectriaceae(M15)、明盘菌科Hyaloscyphaceae(M16)、Pseudeurotiaceae(M17)、革菌科Thelephoraceae(M18)相对丰度均与该林龄段呈极显著相关;其中锥梗菌科Dissoconiaceae(M11)相对丰度正相关程度最强,爪甲团囊菌科Onygenaceae(M5)相对丰度负相关程度最强。在50年及以上林龄阶段,Lachnocladiceae(O1)、双生担子菌科Geminibasidiaceae(O2)、树花科Ramalinaceae(O3)、发夫酵母菌科Phaffomycetaceae(O4)、黑球腔菌科Melanommataceae(O5)、伞形科Umbelopsidaceae(O6)、Mycosphaerellaceae(O7)、Clavicipitaceae(O8)、Xylariales_fam_Incertae_sedis(O9)、Mucoromycotina_fam_Incertae_sedis(O10)、Venturiaceae(O11)相对丰度均与之呈极显著相关;其中黑球腔菌科Melanommataceae(O5)相对丰度正相关程度最大,发夫酵母菌科Phaffomycetaceae(O4)、树花科Ramalinaceae(O3)相对丰度负相关程度最大。
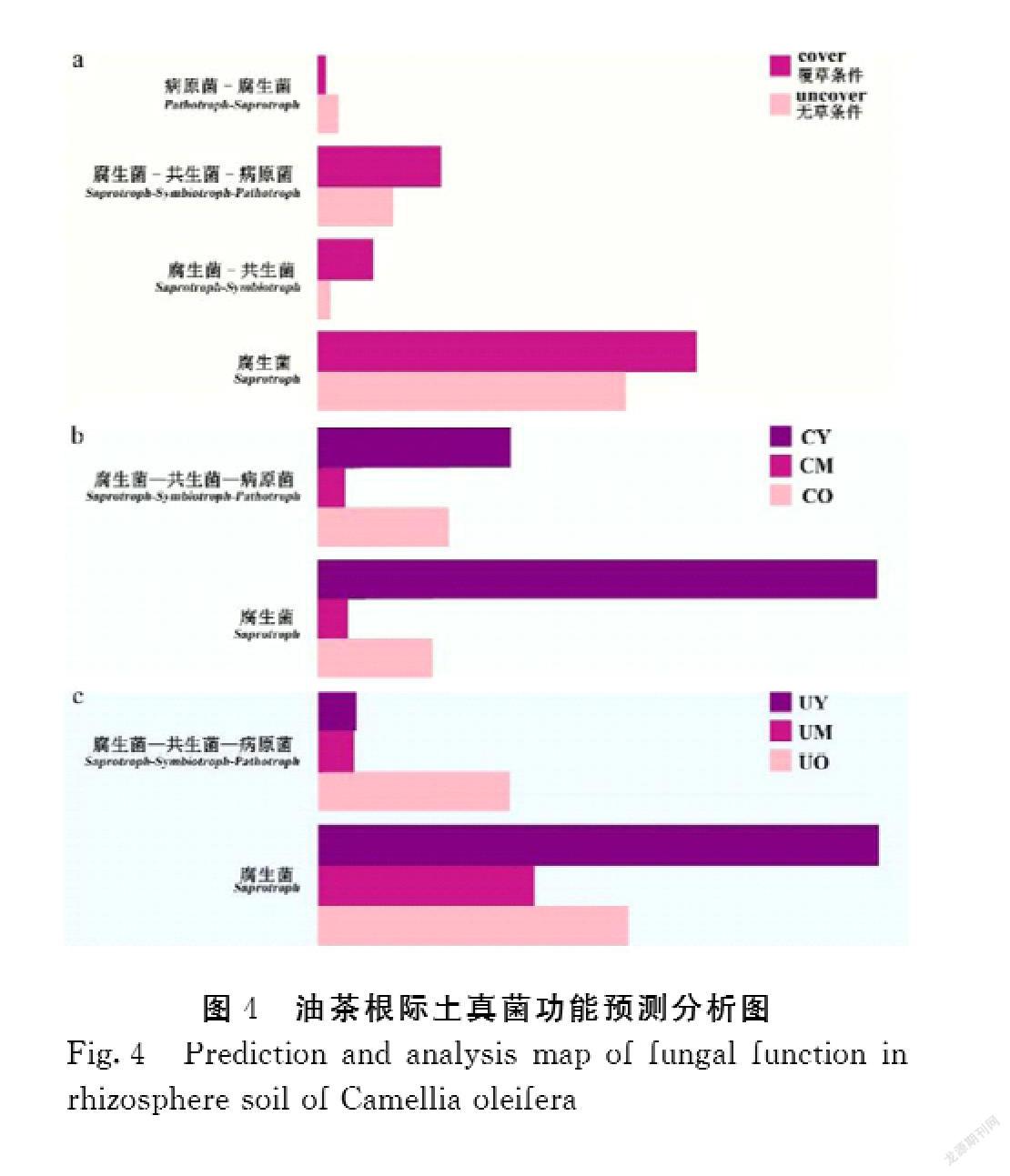
2.5不同种植条件下油茶根际土真菌功能预测分析
本试验中,在有鼠茅草覆盖的样品中腐生菌saprotroph、腐生菌-共生菌saprotroph-symbiotroph和腐生菌-共生菌-病原菌saprotroph-symbiotroph-pathotroph的功能作用高于無草条件的样品(图4a),病原菌-腐生菌pathotroph-saprotroph的功能低于无草覆盖的样品;4种功能中,腐生菌saprotroph的作用显著强于其他3种作用。从年龄层面来看,覆草条件下(图4b),3~5年生组(CY)腐生菌saprotroph、腐生菌-共生菌-病原菌saprotroph-symbiotroph-pathotroph相对丰度显著高于50年生及以上的样品(CO),10~15年生(CM)丰度最低。在无草条件下(图4c)腐生菌saprotroph在UY样品中的相对丰度最高,其次是UO,UM最低,而腐生菌-共生菌-病原菌saprotroph-symbiotroph-pathotroph在UO中的功能作用显著高于UY、UM。
3 结论与讨论
覆草条件的根际土样品内检测出的真菌OTUs数量和特有的OTUs数量均大于无草条件的根际土样品(图1b),表明在地表非裸露的情况下,根际土中的真菌群落更丰富。覆草条件样品的Alpha多样性指数高于无草条件的根际土样品(表1),说明覆草条件的土壤中由于生物多样性相较于裸露的地表更丰富,使得根际土内真菌多样性更高。从年龄层面来看,无草条件的样品中,50年以上林龄(UO)的油茶根际土内真菌的多样性指数最高,说明在土壤裸露的条件下,随着时间的增长,根际土内微生物的多样性随着年龄的增长而增长,这可能与林龄越长,土壤分解了更多的凋落物有关[21];而覆草条件的样品中,10~15年生(CM)的油茶根际土内真菌的丰富度显著高于其他两个年龄组,但50年以上林龄(CO)根际土壤内真菌的多样性显著高于另外两组,这表示植物年龄是根际土微生物生长、发育过程中的重要影响因子。
本研究表明油茶根际土真菌群落在门水平上主要由子囊菌门Ascomycota和担子菌门Basidiomycota组成(图2a),其中3~5年生样品(CY、UY)的相对丰度高于另外两个年龄组。担子菌门可以与植物形成共生关系,促进菌根的生长,油茶生长早期需要大量的根来吸收养分,所以担子菌门相对丰度较高。在科水平上(图2c),优势菌三形菌科Trimorphomycetaceae在3个年龄段中呈现先增长后减少的趋势;在属水平上(图2d),优势菌属三形菌属Saitozyma也呈现先增长后下降的趋势。有研究证明三形菌属Saitozyma是一种耐重金属的真菌[22],此类菌可通过减少重金属对植物生长的抑制作用,从而促进植物修复。属水平上,3~5年生样品的优势菌为篮状菌属Talaromyces;有研究表明篮状菌属Talaromyces具有抑菌、抗虫等作用[23-24]。在50年生及以上样品中,古根菌属Archaeorhizomyces为有鼠茅草覆盖条件下的优势菌,它会响应空气中CO2的变化[25];无草覆盖的条件下,红菇属Russula为优势菌属,对植物养分的吸收有积极作用[26]。
不同林龄阶段油茶根际土真菌微生物的种类及其相关性均有不同,分析发现,陆生壶菌科Terramycetaceae相对丰度与3~5年林龄正相关性最强,陆生壶菌科微生物主要为根瘤菌,在油茶生长初期根瘤菌通过促进根系的生长来帮助油茶吸收养分;锥梗菌科Dissoconiaceae相对丰度与10~15年林龄正相关强度最大,该菌会引发煤污病,降低树木果实的品质[27-28];有研究表明爪甲团囊菌科Onygenaceae相对丰度与植物根系的生长呈正相关[29],而本研究中爪甲团囊菌科Onygenaceae相对丰度与10~15年林龄负相关性最强,这可能是由于这个生长阶段植物发育成熟,根系生长速度开始减缓;黑球腔菌科Melanommataceae相对丰度和50年及以上林龄正相关程度最大,黑球腔菌科Melanommataceae属于格孢腔菌目Pleosporales,该目水平下的菌通常会引起植物叶子变黑、果实腐烂[30-31];相对丰度呈显著负相关的法夫酵母菌科Phaffomycetaceae、树花科Ramalinaceae目前在土壤中的作用尚未明确。
腐生菌saprotroph从宿主中通过降解土壤中的有机物来获取能量。功能预测结果显示,腐生菌saprotroph的功能作用最为显著,且覆草条件下功能作用强于无草条件。在年龄变化条件中,3~5年林龄的油茶林土壤中腐生菌saprotroph功能作用最强,其次为50年生及以上林龄,最后为10~15年林龄,说明在初期腐生菌saprotroph通过大量降解周围生物为油茶根提供营养物质,在生长后期通过降解凋落物来提供养分。
本研究探讨了覆草、无草条件下3个不同林龄的油茶根际土中真菌的变化,覆草条件的根际土真菌相对丰度大于裸露地表的根际土,在油茶种植中可考虑覆草、埋草种植模式,提高微生物种类,丰富微生物群落功能,促进土壤元素的转化,改善土壤肥力[32],促进油茶长势。与张雅倩[33]的研究结果有所不同,本试验中10~15年生油茶根际土的真菌相对丰度最高,但多样性低于50年生及以上林龄的根际土,这可能是由于50年及以上林龄的油茶林分密度大,人为干扰较低,使得微生物群落多样性回升[34];此外生长后期凋落物增多,真菌微生物在转化土壤元素的过程中多样性有所增加[35]。在科水平和属水平上,年龄间的变化呈现先增长后下降的趋势,优势菌在有草的样品中占比更多,且腐生菌的功能显著高于无草条件的样品,说明有鼠茅草覆盖的条件下真菌群落能更好地促进油茶林的生长。综上所述,在种植鼠茅草的条件下,10~15年生油茶根际土真菌微生物丰富度最高,且优势菌在该年龄段占主导地位,在一定程度上促进油茶的生长发育,为油茶林的高收益提供一定的支持。
参考文献:
[1]肖凯英,刘娟.普通油茶育种研究进展[J].生物灾害科学,2021,44(2):114-118.
[2]傅聿青,李江,李建安,等.生草栽培对油茶林地土壤养分特征和微生物及酶活性的影响[J].经济林研究,2018,36(3):82-88.
[3]牛清清,张琳,陈云峰,等.生态栽培对果园水土及养分流失影响的Meta分析[J].生态学杂志,2021,40(10):3175-3183.
[4]郑梅霞,陈宏,苏海兰,等.多花黄精林下仿生态栽培关键技术[J].福建农业科技,2021,52(6):36-39.
[5]黄梅玲.火龙果套种鼠茅草优质高效栽培技术[J].青海农林科技,2019(3):108-109.
[6]李元雪,张广娜,苗悦,等.鼠茅草生草对苹果园土壤理化性质的影响[J].中国果菜,2019,39(10):86-89.
[7]陈汝顶,黄梅玲,刘春冬,黄战友,王志明.鼠茅草的特征特性及高产栽培技术[J].青海农林科技,2019(4):97-99.
[8]BAKKER PETER A H M,DOORNBOS ROGIER F,ZAMIOUDIS CHRISTOS,et al.Induced systemic resistance and the rhizosphere microbiome[J].The plant pathology journal,2013,29(2):DOI:10.5423/PPJ.SI.07.2012.0111.
[9]SANTHANAM RAKESH,LUU VAN THI,WEINHOLD ARNE,et al.Native root-associated bacteria rescue a plant from a sudden-wilt disease that emerged during continuous cropping[J].Proceedings of the National Academy of Sciences of the United States of America,2015,112(36):DOI:10.1073/pnas.1505765112.
[10]周文杰,吕德国,秦嗣军.植物与根际微生物相互作用关系研究进展[J].吉林农业大学学报,2016,38(3):253-260.
[11]沈仁芳,赵学强.土壤微生物在植物获得养分中的作用[J].生态学报,2015,35(20):6584-6591.
[12]罗明霞,胡宗达,刘兴良,等.川西亚高山不同林龄粗枝云杉人工林土壤微生物生物量及酶活性[J].生态学报,2021,41(14):5632-5642.
[13]杨琴,邹凯,陈爱佳,等.不同树龄银杏叶内生真菌的多样性及群落结构分析[J].湖南农业大学学报(自然科学版),2019,45(1):42-49.
[14]肖龙敏,唐明,张好强.不同种植年限宁夏枸杞根际微生物的群落多样性[J].西北林学院学报,2018,33(6):31-39+163.
[15]劳承英,申章佑,李艳英,等.基于高通量测序技术分析不同耕作方式下水稻根际土壤真菌多样性[J].热带作物学报,2021,42(9):2717-2726.
[16]文永均,黄璜,马中刚,等.Illumina高通量测序分析健康三七与患根腐病三七根际土和根内生真菌多样性[J].食品与发酵科技,2020,56(6):22-30.
[17]姚有华,王玉林,姚晓华,等.不同种植模式对青稞根际土壤微生物群落结构的影响[J].西北农业学报,2021,30(04):545-554.
[18]陈凯.小果油茶与普通油茶系统发育关系及果实大小分化的转录组分析[D].南昌:南昌大学,2020.
[19]李河.油茶炭疽病菌群体遗传及MAPK基因CfPMK1功能研究[D].长沙:中南林业科技大学,2018.
[20]彭梁.油茶中两种新病毒的基因克隆及分子特性分析[D].南昌:南昌大学,2019.
[21]宋影,辜夕容,严海元,等.中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J].环境科学,2014,35(3):1151-1158.
[22]TERRY J T C,CEDAR H,CHERYL R K,et al.Presence and distribution of heavy metal tolerant fungi in surface soils of a temperate pine forest[J].Applied Soil Ecology,2018,131:66-74.
[23]柴晓蕾,宋希强,朱婕.华石斛内生真菌组织分布特点及其抑菌活性[J].热带作物学报,2018,39(1):137-144.
[24]何军伟,高昊,刘杏忠,等.湿地真菌Talaromyces flavus中2个新的聚酯类化合物[J].中国中药杂志,2015,40(17):3347-3351.
[25]贾夏,KHADKHUREL L,赵永华,等.连续4个生长季大气CO2升高与土壤铅(Pb)污染耦合下刺槐幼苗根际土壤微生物特征[J].环境科学,2021,42(6):3046-3055.
[26]李国杰,李赛飞,赵东,等.红菇属研究进展[J].菌物学报,2015,34(5):821-848.
[27]李焕宇.煤污病相关真菌属分类及中国种类多样性研究[D].咸阳:西北农林科技大学,2012.
[28]杨迪,杜婵娟,叶云峰,等.广西香蕉煤污病病原菌的分离鉴定及其生物学特性[J].植物病理学报,2021,51(4):507-514.
[29]李发虎,李明,刘金泉,等.生物炭对温室黄瓜根际土壤真菌丰度和根系生长的影响[J].农业机械学报,2017,48(4):265-270,341.
[30]王艳,晋玲,申培增,等.基于形态学及多基因位点的枸杞褐斑病病原鉴定[J].植物保護,2020,46(5):122-127.
[31]冯雅蓉.预冷方式对红玛瑙樱桃果实冷藏期间真菌多样性的影响[J].食品科学,2020,41(11):214-221.
[32]魏倩倩,杨文权,韩明玉,等.白三叶返园对苹果园土壤微生物群落的影响[J].草业科学,2016,33(3):385-392.
[33]张雅倩,黄蕊,左林芝,等.海南岛不同林龄木麻黄凋落物内外细菌多样性及其化感潜力[J].应用生态学报,2020,31(7):2185-2194.
[34]王超群,焦如珍,董玉红,等.不同林龄杉木人工林土壤微生物群落代谢功能差异[J].林业科学,2019,55(5):36-45.
[35]赵辉,周运超,任启飞.不同林龄马尾松人工林土壤微生物群落结构和功能多样性演变[J].土壤学报,2020,57(1):227-238.
(责任编辑:柯文辉)
猜你喜欢
农村百事通(2020年14期)2020-11-23
故事作文·低年级(2020年7期)2020-07-28
农业与技术(2020年5期)2020-03-25
大自然探索(2019年1期)2019-01-24
知识就是力量(2017年12期)2018-01-15
中国中药杂志(2016年24期)2017-04-18
大自然探索(2017年3期)2017-04-07
大自然探索(2017年3期)2017-04-07
中国中药杂志(2017年4期)2017-03-28
中国中药杂志(2017年3期)2017-03-20
