
性早熟儿童肠道菌群结构特征分析
2022-06-02 00:35田佳梅王锁英金胜利王信超边高瑞
医学研究杂志 2022年5期
田佳梅 王锁英 金胜利 刘 欢 王信超 边高瑞
性早熟指女童8岁前,男童9岁前出现第二性特征的发育,或女童10岁前出现月经初潮,女孩的发生率是男孩的5~10倍[1]。随着社会经济发展及国民生活水平的提高,家庭对儿童身心健康及生长发育的关注度在逐年增加,其中对于儿童性早熟的关注度增加较为明显,性早熟的危害不仅在于影响儿童成年身高,且身体心理的不平衡发展容易引起心理障碍,使得青少年危险行为的发生率增加。近年来,儿童性早熟的发生率呈逐年上升趋势,同时发现肥胖儿童中性早熟的发生率较体重指数(body mass index, BMI)正常儿童高,其体重每增加1kg,初潮年龄提前13天,这提示肥胖与性早熟的发生可能存在联系,肥胖可能是性早熟发生的重要因素之一[2,3]。
自全球人类元基因组启动计划至今,随着对人体肠道菌群功能探索的不断深入研究发现,人类肠道菌群具有强大的物质能量代谢功能与机体健康密切相关,被称为人体的第九大系统。已有研究发现,肠道菌群结构的改变可能直接或间接地导致肥胖、糖尿病等内分泌疾病。本研究应用16S rRNA高通量测序的方法,分析了解性早熟儿童肠道菌群的结构特征,对比分析性早熟儿童、肥胖儿童及BMI正常儿童肠道菌群异同,分析性早熟儿童临床指标与肠道菌群之间的相关性,进而探寻肠道菌群与性早熟发生之间的关系,并探讨性早熟与肥胖之间可能的相互关系。
对象与方法
1.研究对象及分组:选择2018年9月~2019年10月因发现乳房增大就诊于江苏大学附属医院儿科的性早熟儿童19例,并按照临床路径完成性激素激发试验等相关检验,按照中枢性性早熟诊断与治疗共识确诊为中枢性性早熟,患儿年龄7.50±0.95岁,选择已于笔者医院确诊为中枢性性早熟14例,患儿年龄8.47±0.63岁,接受曲普瑞林治疗1.59±0.60年,作为性早熟治疗后组。同期于笔者医院门诊体检的单纯性肥胖儿童15例,患儿年龄7.20±1.35岁,BMI正常健康儿童19例,年龄7.24±0.34岁。性早熟组儿童排除垂体、下丘脑及肾上腺等器官疾病导致的继发性性早熟,符合特发性中枢性性早熟特征,所有儿童排除其他遗传、代谢、内分泌系统及中枢神经系统疾病,要求近1个月内无抗生素使用史,无肠道益生菌服用史[1]。研究经笔者医院医学伦理学委员会批准,患儿监护人均知情同意。
2.粪便的采集:采集研究对象新鲜粪便于5ml灭菌干燥离心管中,封口后即放入-80℃冰箱保存。
3.粪便16S rRNA 测序:利用QIAamp®DNA Stool Mini Kit试剂盒(德国QIAGEN公司)提取样本的基因组DNA,用Onedrop仪器和琼脂糖凝胶电泳检测DNA的纯度和浓度。取2.5ng稀释后的基因组DNA作为模板,使用带有Barcode的16S V4通用引物(515F-806R),用GoTaq®Hot Start Colorless Master Mix高效高保真酶(美国Promega公司)进行PCR。
使用Pico Green荧光定量检测PCR产物浓度,根据产物浓度进行等浓度混样,充分混匀后用QIAquick®PCR净化装置试剂盒(美国Qiagen公司)纯化回收产物并开始第2轮扩增,引入Illumina桥式PCR兼容引物,扩增产物用Pico Green荧光定量以及Agilent 2200 TapeStation电泳工作平台检测,合格后上机测序。
采用Illumina MiSeq测序平台(美国Illumina公司)得到下机数据,并对该数据进行预处理,根据Barcode的信息,将单个样品原始数据拆分,取出引物序列,进行测序序列质量控制,通过质量检查的序列,应用核糖体数据库项目(Ribosomal Database Project,RDP) Classifier 2.3,使用Silva数据库进行序列比对,确定每条序列的分类等级;应用Mothur以序列相似性97%为标准进行分类操作单元(operational taxonomic units,OTUs)划分,并按序列数量生成OTUs丰富度谱。
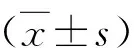
结 果
1.各组儿童肠道菌群Alpha多样性结果分析:Alpha多样性是反映特定区域内生物群落丰富度和均匀度的综合指标。Ace指数及Chao1指数代表微生物群落丰富度,这两者数值越高说明菌群丰富度越高;Shannon指数及Simpson指数代表一定区域内微生物群落的多样性,Shannon指数值越高,表明群落的多样性越高,Simpson指数值越大,说明群落多样性越低。性早熟治疗前组Ace指数及Chao1指数较性早熟治疗后及对照组明显下降(P<0.05),而与肥胖比较,差异无统计学意义(P>0.05);而Shannon指数及Simpson指数在各组间比较,差异无统计学意义(P>0.05),详见表1。

2.各组儿童肠道菌群门水平差异性分析:在肠道菌群门水平,所有样本以拟杆菌门、厚壁菌门、变形菌门、放线菌门及疣微菌门为主要菌门,包含所有样本丰富度99.08%的菌。门水平分布结果显示性早熟治疗前、后组及肥胖组拟杆菌门丰富度较对照组明显升高(P<0.05),而厚壁杆菌门及放线菌门丰富度明显降低(P<0.05),而此3组间拟杆菌门、厚壁杆菌门及放线菌门丰富度比较,差异无统计学意义(P>0.05);与对照组相比较,疣微菌门丰富度在性早熟治疗前、后组明显下降,而性早熟治疗后组较治疗前组明显上升(P<0.05);变形菌门在各组之间变化比较,差异无统计学意义(P>0.05),详见图1,表2。

图1 各组儿童肠道菌群门水平分布

表2 各组肠道菌群门水平比较[M(Q1,Q3)]
3.各组儿童肠道菌群属水平差异性分析:肠道菌群属水平检测出前50个优势菌属,丰富度占所有菌属的97.06%,所有样本的菌群均由这50个菌属及少量其他菌属构成。分析发现性早熟治疗前、后组及肥胖组拟杆菌属、粪杆菌属、普氏菌属、另枝菌属等菌属丰富度均较对照组儿童高(P<0.05),而双歧杆菌属、乳杆菌属、布劳特菌属及黄曲霉菌属明显下降(P<0.05);与性早熟治疗后比较,性早熟治疗前儿童肠道内布劳特菌属丰富度较低,而帕拉杆菌属丰富度增高(P<0.05),详见图2、表3。

表3 各组儿童肠道菌群属水平优势菌比较[M(Q1,Q3)]

图2 各组儿童肠道菌群属水平分布
4.各组儿童肠道菌属PCA分析:各组儿童肠道菌群PCA分析显示性早熟治疗前、后组及肥胖组各样本点之间距离较小,提示此3组儿童肠道菌群差异相对小,而此3组与对照组各样本点之间距离较大,提示其与对照组儿童肠道菌群比较,差异有统计学意义,详见图3。

图3 主成分分析横坐标为第一主成分,28.63%为第一主成分对样品差异的贡献值;纵坐标为第二主成分,10.88%为第二主成分对样品差异的贡献值;图中的每个点表示1个样品
5.各组儿童饮食结构特征:制订统一调查问卷,对纳入研究的儿童日常饮食习惯情况进行统计分析,结果显示,性早熟组儿童饮食偏肉食、服用滋补品、儿童饮料食用概率均较对照组儿童高(P<0.05),与性早熟组类似(P>0.05);性早熟组在反季水果方面概率明显高于肥胖组及对照组(P<0.05);在喜食油炸食品的概率方面,3组间比较,差异无统计学意义(P>0.05),详见表4。

表4 各组儿童饮食结构比较[n(%)]
讨 论
下丘脑-垂体-性腺轴(hypothalamus-pituitary-gonadal,HPG)的启动为青春期开始最直接的因素。研究发现,HPG在青春期启动之前,因下丘脑及垂体对性激素水平较为敏感而处于静止或者抑制状态,随着机体的发育,这种敏感度逐渐下降,HPG内分泌活动逐渐增强,使性腺器官的发育逐渐趋于成熟[4]。性早熟的发生则为HPG的提前启动,研究显示,脂肪组织的囤积可能是青春期启动的因素之一,同时性早熟儿童中肥胖儿童占多数,且本研究中性早熟组肥胖或超重者占54.55%,与此前结果一致[1,3]。研究发现,肥胖人群肠道菌群中拟杆菌门丰富度明显增高,而双歧杆菌等肠道益生菌明显减少,肥胖的发生可能与肠道菌群结构紊乱有关[5,6]。处于青春发育阶段的儿童各方面均处于快速生长阶段,此时期儿童肠道菌群的结构特征值得研究,其作用不可被忽视。
本实验利用16S rRNA高通量测序的方式,研究分析了性早熟、肥胖及BMI正常的健康儿童肠道菌群的结构后发现,未经治疗性早熟儿童肠道菌群的多样性明显低于正常儿童,而与肥胖儿童对比差异无统计学意义,经治疗后的性早熟儿童肠道菌群丰富性及多样性明显升高。据文献报道,肠道菌群丰富度较低的个体体内脂肪含量相对较多,且多数出现胰岛素抵抗,且出现糖尿病等肥胖相关疾病的风险更高[7]。结合本研究,未经治疗性早熟及肥胖儿童肠道菌群丰富度的降低可能促进其体内脂肪囤积,据此亦可解释为何脂肪累积可能是青春期启动的重要信号之一。
分析各组儿童肠道菌群门水平分布发现,性早熟治疗前、后组及肥胖组较正常组儿童肠道具有更为丰富的拟杆菌门,而厚壁菌门及放线菌门丰富度相对较低,此3组间这三大菌门丰富度变化比较,差异无统计学意义,这是否进一步提示性早熟的发生可能与儿童肥胖有关?且肥胖组拟杆菌门及厚壁菌门变化趋势与先前研究一致[5,8]。目前对肥胖个体肠道菌群的研究多集中于动物模型,研究发现肥胖小鼠粪便中拟杆菌门的数量减少而厚壁菌门数量增多,结果与人粪便研究结果存在差异,这还需大量研究来解释肠道菌群与肥胖及相关内分泌疾病的关系[9]。本研究中性早熟治疗前、后组疣微菌门丰富度明显下降,且治疗前组下降更明显,疣微菌门丰富度可否用来预测性早熟的发生?性激素水平是否会影响疣微菌门丰富度?通过何种途径影响?还需要大量研究来探究其中关系,为研究肠道菌群与性早熟发生之间的关系提供一个研究方向。
生理条件下,肠上皮细胞与细菌壁的脂磷壁酸之间形成一层生物屏障来维护肠道内环境的稳态。研究发现,肠道局部及全身长期处于低度炎性状态为肥胖及相关代谢性疾病发生的一个重要因素,肠道菌群结构的改变使肠生物屏障受损,肠道细菌及内毒素等移位后激活免疫系统产生炎性反应。双歧杆菌及乳杆菌等肠道有益菌在维持肠道微生态及酶的平衡方面具有重要作用。本研究进一步分析各组儿童肠道菌群属水平结构后发现,性早熟组双歧杆菌及乳杆菌属等肠道有益菌属丰富度明显降低,拟杆菌属及粪杆菌属丰富度明显升高,提示性早熟儿童肠道菌膜群的构成可能发生改变,肠生物屏障可能受损。此外,普氏菌属、普雷沃菌属及帕拉杆菌属丰富度于性早熟组明显增高,而毛螺菌属及布劳特菌属丰富度下降明显。Hjorth等[10]研究发现,普雷沃菌属及帕拉杆菌属与肠道物质代谢密切相关,给予普雷沃菌属与拟杆菌属比值(prevotella/bacteroides,P/B)不同的个体相同高纤维饮食24周后发现,P/B比值高的个体体重减轻更多,而本研究中性早熟及肥胖儿童P/B比值明显低于对照组,提示性早熟及肥胖儿童能量代谢可能较BMI正常儿童慢。
针对毛螺菌属的研究发现,毛螺菌属可增加肠道对病原体的抵抗性,本研究中性早熟及肥胖组儿童此菌属丰富度明显下降,提示性早熟及肥胖儿童肠道生物屏障的抗性下降,毛螺菌属丰富度的下降可能是肠道慢性炎症的产生的因素之一[11]。布劳特菌属具有抗炎效应,且与粪便中的炎性细胞因子水平相关,在肥胖儿童及有胰岛素抵抗的儿童的粪便中表现更明显[12]。脂肪组织的囤积可增加胰岛素抵抗,肥胖个体胰岛素抵抗更明显,高水平的胰岛素通过增加芳香化酶活性促进睾酮转化为雌激素,使雌激素水平增高,促进青春期启动及发育,本研究中性早熟组及肥胖组布劳特菌属丰富度均明显下降,可能与其体内脂肪组织含量有关[13,14]。那么,毛螺菌属及布劳特菌属可否作为一种生物学标志物来监测儿童肠道炎症程度,从而达到预防性早熟的目的。另枝杆菌属丰富度在性早熟组明显高于肥胖组及正常儿童,且与雌激素水平呈正相关,是否可以通过监测儿童肠道另枝杆菌丰富度来达到预防性早熟发生的可能?
通过对各组儿童饮食习惯的调查研究发现,性早熟及肥胖儿童在饮食偏肉类、常服用滋补品及儿童饮料食用方面概率明显高于对照组,而此两组间比较,差异无统计学意义。Jansen等[15]针对456例女童的饮食情况及月经初潮年龄相关的一项前瞻性研究发现,每周食用红肉4次的女孩月经初潮提前的概率是每周食用2次女孩的1.64倍。研究发现,长期高膳食纤维饮食可明显增加有益菌丰富度[16]。一项针对我国成都市9~15岁的调查研究发现,每天摄入较高膳食纤维的儿童及青少年,其青春期发育相对较晚[17]。这些研究均提示饮食习惯影响健康并可能影响肠道菌群结构,那么是否可通过增加富含膳食纤维及维生素的蔬菜等植物类食物的摄入量,来达到控制体重的目的,远期的达到预防及控制性早熟等相关内分泌疾病的目的。
随着肠道菌群研究的深入,微生态制剂也逐渐丰富起来,益生菌、益生元、合生元及后生素已经广泛应用于医疗、保健等行业。其中益生菌在儿科的应用较为广泛,如双歧杆菌、布拉酵母菌等,但较多为短期的小剂量应用于腹泻及便秘等消化系统疾病的学龄前儿童,调节肠道菌群的作用有限。本实验通过对性早熟及肥胖儿童肠道菌群的研究发现其肠道菌群结构出现明显改变,双歧杆菌及乳杆菌等肠道益生菌丰富度明显下降,那么是否可以利用益生菌的占位效应,通过长期小剂量补充肠道益生菌,来调节菌群结构,修补肠道生物屏障,形成一定的定植抗力,来阻止致病菌的侵入并减少肠黏膜受损。疣微菌门及另枝杆菌属等菌属是否可以作为生物学标志物来监测肠道炎症程度及预测性早熟发生可能性。这为将来探讨肠道菌群紊乱导致儿童性早熟及肥胖提供研究思路,同时也为早期干预治疗儿童性早熟及肥胖提供思路。
猜你喜欢
今日农业(2022年14期)2022-09-15
中老年保健(2022年2期)2022-08-24
传染病信息(2022年2期)2022-07-15
中国饲料(2022年5期)2022-04-26
浙江农业学报(2022年1期)2022-02-18
军事文摘·科学少年(2021年5期)2021-05-27
华声文萃(2020年4期)2020-05-19
大自然探索(2020年1期)2020-02-16
上海预防医学(2017年8期)2017-09-06
百科知识(2017年10期)2017-05-19
