
锌多糖的合成方法、结构特征和生物活性研究进展
2022-08-04 07:27袁心田陈华国赵超龚小见周欣
食品与发酵工业 2022年14期
袁心田,陈华国,赵超,龚小见,周欣*
1(贵州中医药大学,贵州 贵阳,550002)2(贵州师范大学, 贵州省山地环境信息系统与生态环境保护重点实验室,贵州 贵阳,550001)3(贵州师范大学, 贵州省药物质量控制及评价技术工程实验室,贵州 贵阳,550001)
锌是人体第二丰富的微量元素,其含量仅次于铁元素,是人体必需微量元素之一,主要存在于骨骼、肌肉、皮肤和男性生殖器官中。锌是人体300多种酶的重要组成部分,例如超氧化物歧化酶[1]、蛋白酪氨酸激酶-1B[2]和碱性磷酸酶[3],所以锌在蛋白质和DNA合成、细胞生长和增殖、代谢调控等生理过程中起着重要作用。人体缺锌会导致味觉障碍、生长发育不良、皮肤损伤、免疫功能损伤和生殖能力减弱[4]。2017年中国营养学会发布了WS/T 578.3—2017《中国居民膳食营养素参考摄入量》,推荐18岁以上男性每日锌的膳食摄入量为12.5 mg,女性每日锌的膳食摄入量为7.5 mg。人体自身不能合成锌,只能从膳食中摄取,但体内锌元素严重缺乏时应通过补锌制剂维持体内锌元素平衡。
锌多糖作为一种新型的补锌制剂,与传统补锌制剂[4](无机锌、有机酸锌和酵母锌)相比,具有一定的优点。首先,多糖可提高锌的生物利用度,如壳聚糖、海藻酸或生马铃薯淀粉能减轻植酸对锌利用率的抑制作用,提高锌的吸收率和股骨锌浓度[5];枸杞叶多糖能调节大鼠体内锌转运蛋白表达,促进大鼠对锌的吸收[6],由此可以推测,多糖与锌螯合形成的锌多糖可有效提高锌的生物利用度。其次,锌多糖维持了多糖的活性结构,且部分锌多糖的活性要优于原多糖。DONG等[7]、董金满[8]对比罗耳阿太菌多糖和罗耳阿太菌锌多糖的红外光谱和核磁共振波谱,发现罗耳阿太菌锌多糖的基本骨架没有改变,只有部分羟基和羰基发生改变,而罗耳阿太菌锌多糖的体内外抗氧化活性都要优于罗耳阿太菌。本文综述了锌多糖的化学合成法、微生物转化法和植物转化法,并分析各自的优缺点;归纳了锌多糖结构特征分析方法,包括原子吸收光谱分析、气相色谱、高效凝胶渗透色谱、扫描电子显微镜、红外光谱分析、核磁共振波谱分析和热重量分析等;最后总结了锌多糖抗氧化、降血糖、抗炎、抗癌、免疫调节、抗菌和保肝等活性,以期为锌多糖的开发利用提供理论依据。
1 合成方法
天然锌多糖在自然界中存在较少,常存在于植物和菌丝体内,例如金针菇菌丝体内有35%左右的锌与多糖结合[9-10]。天然锌多糖来源较少且含量较低,而使用合成方法可得到锌含量和纯度均较高的锌多糖,还能以不同来源的多糖作为原料进行合成,极大地拓宽了锌多糖的研究范围,所以探索锌多糖合成方法很有必要。目前,锌多糖的合成方法主要包括化学合成法、微生物转化法和植物转化法。
1.1 化学合成法
化学合成法是指利用多糖结构中游离的—OH、—COOH或—NH2吸附锌化试剂的Zn2+,以—O—Zn键或—N—Zn键的形式形成网状结构[11-12],常用的锌化试剂包括硫酸锌、氯化锌和醋酸锌。化学合成法几乎没有改变多糖的空间结构,保持了多糖的大部分生物活性,且操作步骤简单、反应可控、成本低、可制备不同种类的锌多糖,是大部分研究人员合成锌多糖的首选方法。
部分锌多糖的合成条件、锌含量和生物活性如表1所示。由表1可知,不同多糖进行锌化修饰后锌含量有所差异,这可能与多糖的单糖组成、分子质量、糖苷键和空间结构等因素有关。
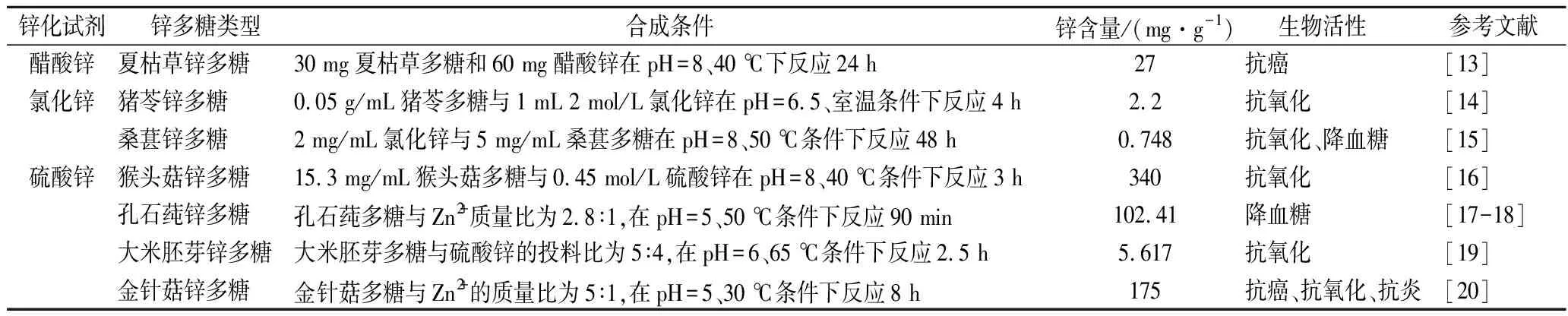
表1 锌多糖的合成条件、锌含量和生物活性Table 1 Synthesis conditions, zinc content amd bioactivities of zinc polysaccharides
1.2 微生物转化法
微生物转化法是指将一定浓度的锌化试剂添加到微生物培养基中,微生物可吸收富锌环境中的锌离子并在体内转化形成锌多糖的方法。微生物转化后的锌多糖可通过传统的多糖提取方法进行直接提取,不同来源的锌多糖提取时间和温度不同。ZHENG等[21]在90 ℃、pH=8条件下提取2 h得到光帽鳞伞SW-02菌丝体锌多糖,ZHANG等[22]在80 ℃提取3 h得到灰树花SH-05胞内锌多糖。此外,可使用碱、酸或酶作为辅助技术提高锌多糖提取率。许诺[23]、XU等[24-25]通过向培养基中加入醋酸锌在白灵菇菌丝体内富集锌元素,以HCl、NaOH和4 %的蜗牛酶溶液作为溶剂提取出白灵菇菌丝体锌多糖,分别得到酸提白灵菇菌丝锌多糖(AcMZPS,由甘露糖、半乳糖和葡萄糖组成,摩尔比为4.0∶5.97∶1.9)、碱提白灵菇菌丝锌多糖(AlMZPS,由半乳糖和葡萄糖组成,摩尔比为6.55∶38)和酶提白灵菇菌丝锌多糖(EnMZPS,由葡萄糖、半乳糖和甘露糖组成,摩尔比1.3∶1.0∶3.8)。
1.3 植物转化法
植物转化法是指植物在酶系统的作用下,通过新陈代谢将土壤或培养基中的无机锌转化为有机锌的方法。已有文献表明土壤中的无机锌可通过植物转化法转化为锌多糖,YANG等[26]用ZnSO4得到高锌含量的糙米,该糙米最佳培养条件为ZnSO4200 mg/L,浸泡时间30.28 min,培养时间3 d,在此条件下糙米的最大锌含量为304.71 μg/g。糙米体内的有机锌主要以锌多糖、锌蛋白、锌核酸、脂质和低分子质量化合物等形式固定和储存在植物体内,其中锌多糖占比最高为30.42 %。FAN等[27]用不同浓度的ZnSO4处理金钗石斛,观察其在不同锌浓度下的生长情况,结果表明低浓度锌(< 400 μmol/L)可增加金钗石斛的光合速率和蒸腾作用,提高金钗石斛叶片中抗氧化酶活性。高浓度锌元素(800 μmol/L)可促进金钗石斛体内的多糖与锌螯合形成锌多糖,减轻锌胁迫对其造成的损害。已有文献采用植物转化法得到硒多糖并进行深入研究[28],而鲜有文献对植物转化法得到的锌多糖进行深入研究,需要研究工作者今后弥补该方面的空白。
综上可以看出每种锌多糖合成方法的优缺点。化学合成法无需培养菌丝体或植物,实验周期短、操作步骤少、成本低且锌元素应用率高,适用于工业生产,但锌多糖的锌含量受到反应条件和多糖结构等因素影响,需要找到最佳的实验条件。微生物转化法的产物活性较稳定、毒性较小、反应绿色环保,但存在反应过程机制尚不明确和产物难分离等问题。植物转化法的研究较少,且存在植物培养周期长的问题,但该方法操作简单、反应过程绿色环保、可应用于农业生产中。制备锌多糖时应考虑多糖种类、操作过程难易程度、成本等问题,选择适当的制备方法制备锌多糖,不同制备方法得到的锌多糖性质可能有差异,需要进行比较以选择最好的制备方法。
2 结构特征分析方法
锌多糖的基本骨架结构仍是多糖链,所以锌多糖的结构可采用表征多糖结构的方法进行分析,包括原子吸收光谱分析、气相色谱、高效凝胶渗透色谱、扫描电子显微镜、红外光谱分析、核磁共振波谱分析和热重量分析,以获取锌多糖的锌含量、单糖组成、分子质量、微观表面结构、结构特征和热稳定性等信息。
2.1 原子吸收光谱(atomic absorption spectroscopy,AAS)分析
AAS是基于物质所产生的原子蒸气对特定谱线的吸收作用来进行定量分析的一种方法,是测定锌多糖中锌含量的一种方法。锌多糖在HNO3-HClO4混合溶液中水解12 h,适当稀释水解液后使用AAS分析其锌含量[21]。LI等[13]通过AAS测定夏枯草锌多糖中锌含量为(27.0±2.4) mg/g,XUE等[16]通过AAS测定猴头菇锌多糖中锌含量为340 mg/g。AAS具有检测限低、准确度高、分析速度快、分析范围广等优点,但容易产生干扰效应,使结果产生误差,需配合其他方法如X射线能谱仪分析法使测量结果准确。
2.2 气相色谱(gas chromatography,GC)
GC采用气体作为流动相,利用物质的沸点、极性及吸附性质的差异来实现混合物的分离,可定量分析锌多糖的单糖组成,有助于判断哪种多糖更适合制备锌多糖。ZHANG等[29]通过GC分析姬菇SS-03锌多糖(intracellular zinc polysaccharides fromPleurotuscornucopiaeSS-03,IZPS)的单糖组成,结果表明IZPS的单糖分别为鼠李糖、木糖、甘露糖、葡萄糖和半乳糖,含量分别为29.25%、0.78%、19.45%、29.84%和20.67%,摩尔比为1.65∶0.05∶1∶1.50∶1.07。ZHENG等[21]测定光帽鳞伞SW-02菌丝体锌多糖的单糖组成,光帽鳞伞SW-02菌丝体锌多糖由葡萄糖、甘露糖、半乳糖和阿拉伯糖组成,摩尔比为33.04∶4.49∶4.28∶1。研究发现,提取方法的不同会影响锌多糖的单糖组成,进而影响其生物活性[23]。
2.3 高效凝胶渗透色谱(high performance gel permeation chromatography,HPGPC)
HPGPC是测定锌多糖分子质量大小和分布的一种常用手段,其分离原理是基于溶液中分子体积的大小差异进行分离。WANG等[30]测定香菇SD-08菌丝体锌多糖(mycelia zinc polysaccharides ofLentinusedodesSD-08,MZPS)的分子质量,其重均分子质量为1.20×105Da,数均分子质量为7.14×102Da。通常采用分子质量分布系数(Mw/Mn)表示聚合物的分散性,分布系数越大说明聚合物分子质量越分散。MZPS的Mw/Mn为168.19,表明MZPS的分子质量分布较宽,长短链混杂。WANG等[15]测定了桑葚锌多糖的分子质量,其平均分子质量为1.561×105Da。硒多糖的分子质量与生物活性具有一定关系[31],而锌多糖分子质量与生物活性之间的研究较为缺乏,这可能是锌多糖构效关系的研究方向之一。
2.4 扫描电子显微镜(scanning electron microscope,SEM)
SEM利用聚焦窄的高能电子束来扫描样品, 通过光束与物质间的相互作用, 反映各种物理信息, 对这些信息收集、放大、再成像以达到对物质微观表面结构表征的目的。SEM可观察锌多糖的微观表面结构,通过对比多糖和锌多糖SEM的差异,结合其他检测方法确定锌是否与多糖螯合。猴头菇多糖(Hericiumerinaceuspolysaccharides,HEP)的微观结构为不规则的海绵状,提供了较大的螯合面积。XUE等[16]观察到HEP-Zn的结构表面有鳞片,表明锌被吸附在HEP表面,形成HEP-Zn。锌多糖与多糖表面结构的差异会导致两者生物活性的差异。ZHANG等[32]发现,锌与平贝母多糖(Fritillariaussuriensispolysaccharides,FUP)螯合后,FUP与锌在生物活性方面的协同作用和FUP-Zn较大的孔径是导致FUP-Zn抗氧化活性增强的原因。SEM可直观地揭示锌多糖的微观表面结构,但无法获得共价键连接方式和内部空间结构等信息,需要结合红外光谱分析和核磁共振分析等方法对锌多糖的结构进行分析。
2.5 红外光谱(infrared absorption spectrometry,IR)分析
IR分析是利用红外光谱对物质分子的分析和鉴定,将一束不同波长的红外射线照射到物质的分子上,某些特定波长的红外射线被吸收,形成这一分子的红外吸收光谱。IR可测定多糖和锌多糖的官能团信息,通过对比多糖与锌多糖的IR,可判断锌元素是否螯合在多糖上以及推测锌元素连接官能团的种类。对比大蒜多糖和大蒜锌多糖的IR,两者红外光谱相似,说明锌化修饰没有改变多糖的基本骨架。大蒜锌多糖在3 368 cm-1吸收峰变弱,并向3 286 cm-1移动,1 600 cm-1和424 cm-1处出现弱吸收峰,说明Zn2+与多糖螯合成功[33]。对比修饰前后的IR图,发现肉苁蓉锌多糖的—OH伸缩振动峰蓝移,—COO—的反对称伸缩振动峰蓝移,说明锌与—COOH和—OH发生螯合[34]。桑葚锌多糖的—OH伸缩振动峰发生红移,并在2 146 cm-1和984 cm-1处出现新的吸收峰,推测产生了—O—Zn[15]。IR操作简单、结果易于分析,但只能分析出锌多糖的官能团相关信息,锌多糖的精确结构需要甲基化分析和核磁共振波谱分析进行佐证。
2.6 核磁共振波谱(nuclear magnetic resonance spectroscopy,NMR)分析
NMR分析是指原子核在外加磁场中吸收从一个自旋能级到另一个自旋能级的电磁波后产生的吸收光谱。NMR作为一种分析化学技术已被广泛应用于研究多糖的分子结构和构象,结合一维谱、同核和异核二维核磁共振谱中的化学位移和偶合常数可以推测糖基的连接和序列。DONG等[7]、董金满[8]收集了罗耳阿太菌锌多糖的核磁共振氢谱和碳谱,发现罗耳阿太菌锌多糖的化学位移主要集中在δ 3.0~5.5,这是多糖的典型特征信号;在δ 5.2和δ 4.7 处的异常质子信号表明,α和β构型都存在;在δ 5.4处没有信号进一步证实了该锌多糖属于吡喃糖。由碳谱可知,在δ 80~90处没有信号,可进一步证实吡喃糖的存在。此外,罗耳阿太菌锌多糖氢谱中的峰强度明显减弱,质子信号的峰型也有一定程度的展宽,说明锌与多糖发生了螯合。NMR是分析多糖结构和构象的常用工具,然而NMR仍存在一些局限性[35]:(1)NMR需要更高的分辨率和灵敏度,才能最大限度地提高结构解析能力;(2)结构复杂或黏度高的多糖仍然是核磁共振分析的巨大障碍;(3)核磁共振分析的规则不统一,包括测试温度和内标。
2.7 热重量分析(thermogravimetric analysis,TGA)
TGA是在程序控制温度下,测量物质的质量与温度或时间的关系的方法。通过分析热重曲线,可知样品及其可能产生的中间产物的组成、热稳定性、热分解情况及生成的产物等与质量相关联的信息。平贝母锌多糖发生热分解的温度高于平贝母多糖,且多糖损失率更低,说明多糖-锌螯合物的热稳定性比未修饰的多糖高[32]。亚侧耳锌多糖(zinc-Hohenbueheliaserotinapolysaccharides,Zn-HSP)在TGA中损失质量主要包括3个阶段:(1)在20~240 ℃,Zn-HSP的质量缓慢下降,主要是以吸附或氢结合的方式结合在锌多糖表面的水蒸发导致的。(2)在240~500 ℃,Zn—HSP热分解,导致其质量迅速下降。(3)随着温度不断上升,Zn—HSP质量趋于恒定。在20~500 ℃,亚侧耳多糖质量损失略大于Zn—HSP,其原因可能是引入Zn—O基团取代了亚侧耳多糖中的—OH,导致结合水的减少,热损失质量减少[11]。
2)退出控制电源、储能电源后,继保人员手动复归,两信号消失。投分一次控制电源后,“告警”信号又出现,且无法复归。保护装置掉电重启后,告警信号可手动复归。再次分投一次控制电源,告警信号再次无法手动复归。
除了上述方法分析锌多糖的结构,高效液相色谱[36]和甲基化[15]也是常用的分析锌多糖结构的方法。目前,锌多糖的结构较为复杂、不同锌多糖之间结构差异较大、结构分析方法较多,需对每一种锌多糖选择合适的分析方法进行分析。
3 生物活性
已有实验表明,与未锌化的多糖相比,锌多糖在许多方面均表现出生物活性的提高,包括抗氧化、降血糖、抗炎、抗癌、免疫调节、抗菌和保肝等活性。
3.1 抗氧化
氧化应激是由于促氧化剂和抗氧化剂之间缺乏平衡造成的。活性氧(reactive oxygen species,ROS)过度增加、抗氧化剂不足或细胞缓冲系统未能维持氧化还原,会导致平衡失调和生物分子变化,最终引发各种疾病,如癌症、肾损伤、各种炎症和糖尿病等[37]。研究表明,ZnSO4和枸杞多糖能协同抑制酒精性肝损伤所带来的氧化应激,表明多糖和锌都具有抗氧化活性[38]。所以,锌多糖的抗氧化活性研究一直是主要研究的方向,在已发表的文中找到许多有关不同来源和制备方法的锌多糖抗氧化活性的研究,包括体内动物实验和体外自由基清除活性实验。表2总结了锌多糖体内外抗氧化活性研究。

表2 锌多糖体内外抗氧化活性研究Table 2 Study on antioxidant activities of zinc polysaccharide in vivo and in vitro
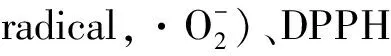
此外,锌多糖具有抗衰老活性,其原因在于锌多糖具有较好的抗氧化活性。衰老是一种以渐进性生理功能损害为特征的自然过程,可导致个体发生多种与衰老相关的退行性疾病。目前,越来越多的证据表明,氧化应激在衰老过程中起着重要作用,最流行的解释衰老过程的理论之一是自由基学说[47]。氧在代谢过程中会产生自由基,但过量的自由基对人体是有害的。常用D-半乳糖建立衰老模型,体内抗氧化酶活性下降可确定造模成功。王丽芹[48]通过D-半乳糖对小鼠进行造模,造模小鼠体内SOD、CAT和GSH-Px活性下降,说明衰老模型造模成功。给予小鼠香菇锌多糖后,小鼠心、肝、肾、血中SOD、CAT和GSH-Px活性显著提高,MDA含量显著降低,说明香菇锌多糖具有抗衰老作用。
3.2 降血糖
糖尿病是一种因胰岛素分泌不足或靶细胞对胰岛素敏感性降低引起的代谢紊乱。锌在人体中起重要作用,补锌可促进血糖水平的适度降低[49]。糖尿病患者体内锌含量不足,缺锌可能会对体内锌依赖激素和酶的功能产生不利影响,从而导致不必要的并发症。多糖能调节磷酸肌醇3激酶(PI3K)/蛋白激酶B(Akt/PKB)信号通路、核因子E2相关因子2(Nrf2)和炎症信号通路,起到降血糖作用[50]。二者都具有降血糖作用,因此锌和多糖结合形成的锌多糖,其降血糖活性受到科技研究人员关注。
3.3 抗炎
炎症是人体对于刺激的一种防御反应,常见于抵御外来病原体侵袭或修复感染造成的组织损伤,然而在某些情况下,炎症反应本身会损害宿主组织并导致器官功能障碍。脂多糖可诱导炎症的产生,促使炎症细胞分泌肿瘤坏死因子(TNF-α)、INF-γ、白细胞介素-6(IL-6)和NO等炎症因子。金针菇锌多糖可降低脂多糖诱导的RAW 264.7细胞中TNF-α、INF-γ、IL-6和NO水平,表现出良好的抗炎活性,且活性与浓度呈正相关[12]。干巴菌菌丝体锌多糖能清除斑马鱼体内自由基,降低脂多糖诱导的ROS水平,抑制中性粒细胞向损伤部位迁移和浸润,起到抗炎作用[53]。浒苔锌多糖可显著抑制仔猪空肠黏膜中NF-κB的激活,并减弱炎症因子的释放,表明浒苔锌多糖可以作为抗生素的替代品添加到断奶仔猪饲料中,增强其免疫调节提高存活率[54]。
3.4 抗癌
癌症是以细胞异常增殖为显著特征的一种疾病,目前治疗癌症主要依靠手术和放射性治疗,对人体有严重损害,因此找出并利用具有抗肿瘤活性且低毒副作用的天然活性物质成为治疗癌症的热点课题。LI等[13]发现,夏枯草锌多糖可以抑制肝癌细胞增殖,有潜在的抗肝癌活性,其表现为细胞形态改变、染色质凝聚和G0/G1期细胞周期阻滞,深入研究发现,夏枯草锌多糖使ROS过度增加,降低线粒体膜电位,激活caspase-3和caspase-9信号通路,诱导HepG2细胞凋亡。张笑然等[55]发现,姬松茸胞内锌多糖能抑制小鼠体内肝癌细胞增殖,对脾脏的生长有一定的促进作用,同时还能提高血清中锌含量,表明姬松茸胞内锌多糖具有抗肝癌和补锌的功能。YAN等[56]对比了茯苓多糖、茯苓锌多糖、茯苓硒多糖和茯苓铁多糖抑制卵巢癌细胞A2780的增殖能力,结果表明茯苓锌多糖的抑制能力最强,在20 μg/mL质量浓度下,A2780细胞的存活率仅为81.087%。LIAO等[57]发现竹荪锌多糖可阻滞细胞周期S期、破坏细胞线粒体功能和诱导细胞内ROS过度生成,同时激活caspase通路,促进人乳腺癌细胞MCF-7的凋亡。
3.5 免疫调节
免疫系统在人体抵御病毒或细菌感染中起着至关重要的作用,但在某些情况下免疫系统会被抑制或失调,因此寻找新的干预措施改善机体的免疫反应是预防或治疗疾病的关键方向之一。锌多糖具有较好的免疫调节活性,可通过多种途径进行免疫调节。阿苯达唑是唯一治疗泡状棘球蚴病的药物,但该药会引起全身性免疫抑制,具有强烈的副作用,而锌葡聚糖螯合物与阿苯达唑联合使用可减少阿苯达唑的副作用。锌葡聚糖螯合物具有免疫刺激活性,可促进小鼠T淋巴细胞增殖,提高IFN-γ水平,激活Th1型细胞免疫应答,减轻阿苯达唑所产生的全身性免疫抑制[58]。蜜蜂采集的枸杞花粉多糖(polysaccharides from bee collected pollen of Chinese wolfberry,WBPPS)是一种天然的锌多糖,ZHOU等[59]从WBPPS中分离出3个组分,并评价其免疫调节活性。结果表明,WBPPS-1、WBPPS-2和WBPPS-3均能提高RAW 264.7细胞NO、TNF-α、IL-6和IL-1β水平,具有良好的免疫刺激作用,其中WBPPS-3的免疫刺激活性最好,推测与WBPPS-3单糖组成中较多的半乳糖和鼠李糖有关。
3.6 其他
除了上述活性之外,锌多糖还具有抗菌和保肝活性,以及存在潜在的调节肠道菌群活性的可能性。ZHANG等[22]在培养基中加入锌化试剂,通过微生物转化法合成灰树花菌株锌多糖(intracellular zinc polysaccharides fromGrifolafrondosaSH-05,IZPS),并测定IZPS和IPS(intracellular polysaccharides fromGrifolafrondosaSH-05,IPS)的抗菌活性。结果表明,IZPS具有较好的抑菌活性,抑菌活性大小顺序为:金黄色葡萄球菌>大肠杆菌>单增李斯特氏菌>巨大芽孢杆菌。此外,与IPS相比,IZPS的抑菌活性更强,说明多糖锌化是提高多糖抑菌能力的一种有效手段。NETANEL等[60]发现紫球藻锌多糖对大肠杆菌和枯草芽孢杆菌的抑菌活性均强于天然多糖,紫球藻锌多糖不仅抑制了细菌的生长,而且减少了活细胞的数量,表现出较强的抗菌活性。ZHANG等[36]研究了红平菇菌丝锌多糖对CCl4所致急性肝损伤的保护作用。红平菇菌丝锌多糖能降低血清中谷丙转氨酶、谷草转氨酶和肝脏指数,显著升高血清中SOD、GSH-Px和CAT活性,具有抗氧化和保肝作用。目前没有相关文献研究锌多糖的调节肠道菌群活性,但锌[61]和多糖[42]均能调节由疾病引起的肠道菌群紊乱,本文猜想锌多糖同样具有调节肠道菌群的活性,这是科技研究工作者今后研究锌多糖生物活性的方向之一。
4 总结与展望
锌多糖是一种新型的补锌制剂,具有高生物利用度、低毒性、易消化吸收等特点。现有研究采用了化学合成法、微生物转化法和植物转化法制备锌多糖,这些方法均能将多糖与无机锌螯合,且稳定性良好。目前锌多糖的结构分析主要采用AAS、GC、HPGPC、SEM、IR、NMR和TGA等方法,获取锌多糖的锌含量、单糖组成、分子质量、微观表面结构、结构特征和热稳定性等结构信息,这有助于研究其构效关系。部分锌多糖表现出比天然多糖更好的生物活性,包括抗氧化、降血糖、抗炎、抗癌、免疫调节、抗菌和保肝等,说明锌多糖在医学领域和保健食品领域具有良好的应用价值和广阔的前景。
随着研究逐渐深入,锌多糖的研究仍然存在诸多问题,需要在今后的研究中加以解决。
(1)不同制备方法得到的锌多糖的锌含量差异较大,然而目前没有证据表明锌含量提高会增强其生物活性,因此不同锌含量锌多糖对生物活性的影响值得深入研究。
(2)多糖是一种天然大分子物质,结构较为复杂,引入锌会导致多糖结构更为复杂。目前锌多糖结构分析方法仍具有一定的局限性,采用更加新颖的方法全面地解析锌多糖结构是该领域研究的方向之一。此外,锌多糖的构-效关系还需要更多的研究。
(3)在锌多糖生物活性相关研究中,大部分研究集中在抗氧化和降血糖方面,相比之下抗炎、抗癌、免疫调节、抗菌、保肝和调节肠道菌群活性的研究较少,应重视这些活性的研究。
(4)许多体内外实验证明了锌多糖有一定的生物活性,但其作用机制尚未研究。
(5)目前锌多糖缺乏临床试验数据,在开发成为新型功能食品之前,必须收集充足的临床试验数据,确保锌多糖对人体安全有效。
因此,需对上述问题进行重点研究,这对锌多糖基础理论的研究、锌多糖作为新一代功能食品或药物辅助治疗剂的研发和生产都具有十分重要的意义。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
世界科学技术-中医药现代化(2021年8期)2021-12-21
载人航天(2021年5期)2021-11-20
中老年保健(2021年9期)2021-08-24
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
中成药(2019年12期)2020-01-04
世界农药(2019年4期)2019-12-30
中央民族大学学报(自然科学版)(2018年1期)2018-06-27
药学研究(2015年11期)2015-12-19
食品工业科技(2014年23期)2014-03-11
