
大豆蛋白源性肽调节糖脂代谢机制研究进展
2022-11-30 07:04李岩异吕娜陈金利李晓张卫婷张红霞
生物技术进展 2022年6期
李岩异,吕娜,陈金利,李晓,张卫婷,张红霞
华北制药金坦生物技术股份有限公司,石家庄 050035
大豆蛋白源性肽(soybean protein derived peptides)是大豆蛋白酶的降解产物,是植物源性肽的重要来源,它能平衡人体内必需氨基酸含量。已有研究发现,除大豆异黄酮和低聚糖等功能成分外,许多从大豆蛋白中提取的功能肽具有多种功能,如改善脂代谢、糖代谢等[1]。Cabanos等[2]研究发现,与摄入动物蛋白(如酪蛋白)相比,摄入植物蛋白(如大豆蛋白)后,机体血液中胆固醇的浓度增加较少。人们的生活方式和饮食习惯与很多疾病密切相关,尤其是动脉硬化、糖代谢等相关疾病,通过改变饮食结构、改善胆固醇代谢和糖代谢是当前预防和治疗高胆固醇血症、高脂血症和糖尿病等疾病的重要策略。本文综述了大豆蛋白源性肽在体内外改善脂质和糖代谢方面的生理功能,并对大豆源性肽的前景进行了展望,以期为大豆蛋白源性肽的研究与应用提供参考。
1 大豆蛋白源性肽
大豆蛋白源性肽是由大豆蛋白经过酶水解或者微生物发酵产生的,包含多种且分子链长度不等的肽类混合物,通常情况下,分子量在1 000以下。除了含有氨基酸外,还含有少量的游离氨基酸、糖类和无机盐等。早在20世纪40年代,欧美科学家就对大豆蛋白源性肽进行了初步研究,但直到六七十年代,随着生物技术和生命科学的快速发展,大豆蛋白源性肽的生物学活性才被人们重视。欧美国家从20世纪70年代就开始了大豆蛋白源性的规模化生产和研究。我国是20世纪80年代末开始进行该领域的研究,经过近几十年的发展,在大豆蛋白源性肽的生产、开发、应用方面均取得了显著的进展。大豆蛋白源性肽除了具有易吸收的特点外,还具有降低胆固醇、降血脂和降血糖的作用,因此大豆蛋白源性肽被广泛应用于食品和保健品中[3]。
2 大豆蛋白源性肽在脂代谢中的调节作用
脂类是三大营养成分之一,是机体能量最重要的储存形式,可分为胆固醇(total cholesterol,TC)、甘油三酯(triglyceride,TG)和磷脂(phospholipid,PL),其中TC是细胞浆膜、维生素和胆汁酸的重要组成成分,TG主要参与人体的能量代谢,PL是构成磷脂双分子层的主要组成成分。脂质代谢异常会引起慢性疾病,例如高胆固醇血症、高脂血症、动脉硬化和肥胖等。长期摄入大豆蛋白源性肽,可以有效调节胆固醇、甘油三酯代谢,维持机体功能健康[4]。
2.1 大豆蛋白源性肽对胆固醇的调节作用
胆固醇又称胆甾醇,属于环戊烷多羟菲的衍生物,它在人体内有广泛的生理作用和功能,当在人体内含量高时,会导致高胆固醇血症,进而引起一系列心血管疾病,对人体健康造成威胁。大豆蛋白源性肽主要通过抑制胆固醇的吸收和调节胆固醇的体内代谢两种途径调节、改善人体胆固醇代谢。
2.1.1 抑制胆固醇的吸收大豆蛋白源性肽之所以能够降低血清胆固醇含量,部分源自大豆蛋白源性肽中的疏水肽在抑制肠道胆固醇吸收方面起重要作用[5-6]。胆固醇吸收主要在小肠中进行,吸收过程中,胶束形成是胆固醇吸收的前提,也是胆固醇吸收的主要途径。胆固醇在小肠中形成胆固醇胶束后,穿越小肠的绒毛壁,被吸收进入血液,最终导致血液中胆固醇含量升高。因此,抑制胆固醇胶束溶解度可以降低小肠对胆固醇的吸收。胶束由亲水区和疏水区组成,疏水肽进入胶束后,能够破坏胆固醇胶束的形成,降低其稳定性,被破坏的胶束释放出的胆固醇被排泄出体外[7]。
使用放射性胆固醇进行动物吸收实验表明,大豆蛋白源性肽能抑制大鼠对胆固醇的吸收。进一步对大豆蛋白源性肽成分分析发现,其存在一种最疏水的序列VAWWMY,以及疏水性氨基酸残基Trp和Tyr。它能够降低胆固醇胶束溶解度,是目前唯一一种在体内抑制胆固醇吸收的寡肽,因此,VAWWMY序列又被命名为大豆抑素[8]。
2.1.2 调节胆固醇的体内代谢胆固醇代谢主要发生在肝脏处,胆固醇受前蛋白转化酶枯草杆菌蛋 白 酶9(proprotein convertase subtilisin/kexin type 9,PCSK9)、载脂蛋白E、调节原件结合蛋白2
(sterol regulatory element binding protein2,SREBP2)和胆固醇7α-羟化酶(cholesterol 7α-hydroxylase,CYP7A1)等一系列蛋白和酶的作用,在肝脏内,转化为胆汁酸,大部分胆汁酸会随着胆汁进入小肠,通过肝肠循环重新进行肝脏代谢[9-10]。
PCSK9是一种由PCSK9基因编码的酶,可与肝细胞表面的低密度脂蛋白胆固醇受体(low density lipoprotein cholesterol receptor,LDL-R)相结合,血液中的低密度脂蛋白胆固醇(low-density lipoprotein cholesterol,LDL-C)主要依赖LDL-R清除,如果PCSK9被抑制,则会有更多的LDL-R出现在肝脏表面,从而清除血液中更多的LDL-C[11]。
体外HepG2细胞培养实验发现,大豆蛋白源性肽中存在的露那辛肽能够降低PCSK9的表达水平,提高LDL-R蛋白的表达,促进HepG2细胞对LDL-C的摄取[12]。在载脂蛋白E基因缺陷型小鼠模型中,腹腔注射露那辛肽后,可降低血清中总胆固醇和低密度脂蛋白胆固醇水平,其机制是露那辛肽在体内促进PCSK9和LDLR的表达[12-13]。因此,在临床上,露那辛肽与辛伐他汀联合使用可降低患者血清的总胆固醇和低密度脂蛋白胆固醇水平[14]。
SREBP2蛋白在调节肝脏中胆固醇水平时,负责外源胆固醇的摄入和内源性胆固醇的合成。从大豆球蛋白源性肽中提取的3种肽(IAVPGEVA、IAVPTGVA和LPYP)能够在体外激活HepG2细胞中SREBP2途径[15],从而促进胆固醇代谢。此外,通过对上述3种肽采用荧光染料标记后,发现这3种肽能够进入HepG2细胞[15]。推测这3种大豆蛋白源性肽直接调节肝细胞中的胆固醇代谢,然而,这3种肽在体内的降胆固醇作用还需要进一步证实[15]。
CYP7A1是胆固醇转化成胆汁酸过程的重要限速酶。有报道显示,IIAEK(一种来源于大豆蛋白源性肽或牛乳蛋白水解物的寡肽,具有调节脂质代谢的作用)可能通过和特定受体[5,11]或细胞膜中的特定蛋白质结合,从而在体内调节胆固醇代谢[16]。构成IIAEK的氨基酸序列中C末端赖氨酸残基(K)诱导了CYP7A1表达,二肽EK是CYP7A1 mRNA的IIAEK诱导中最小的单位,因此推测,EK是低胆固醇的关键结构分子[17-18]。
大豆蛋白源性肽具有降低血清胆固醇含量的作用,这与胆汁酸的结合活性有关。机体内的胆固醇由肝脏分解为胆汁酸,胆汁酸与胆汁酸螯合剂结合后形成不溶性的螯合复合物,这些不溶性的螯合复合物不能被机体再吸收,从而被排出体外,降低机体内胆固醇的水平。因此,具有胆汁酸结合力的物质可以降低胆汁酸的含量,降低内源性胆固醇的含量。
就这样,多变的情境,充满挑战的活动,许多同伴的参与,可供选择的丰富的器械和材料……动态的场景、动态的人满足了幼儿好动的天性,让幼儿拥有了更多活动的选择权;而共同的活动、共同的挑战形成了同伴之间的和谐互动,可以提高幼儿之间的交往能力和语言表达能力,从而培养幼儿良好的个性和养成自觉参加户外体育活动的好习惯。
先前有研究表明,经胃蛋白酶水解获得的大豆蛋白源性肽可以作为开发获得大豆蛋白源性肽活性成分的有效途径[17]。结肠腺癌细胞(colon adenocarcinoma cells,Caco-2)能自然分化成与成熟小肠细胞具有相似形态和生化特征的细胞,因此Caco-2细胞可以作为研究小肠吸收的体外模型。对Caco-2体外培养实验研究表明,SPH可以降低血清中胆固醇含量[19]。这是由于SPH中存在一种疏水性胆汁酸结合肽,它可以与肠道内的胆汁酸结合,抑制胆固醇胶束的形成以及胆固醇在肠道内的吸收[20]。通过胆汁酸结合能力测试实验发现,SPH中的VAWWMY序列具有很强的胆汁酸结合能力,在大鼠体内大豆蛋白源性肽与胆苯丙胺类药物具有相同的活性[8]。
大豆蛋白源性肽的磷脂复合物(soy protein peptic hydrolysate with phospholipids,SPHP)也被证明在大鼠体内具有降低胆固醇的作用[21]。与传统大豆蛋白源性肽相比,SPHP对胆固醇代谢改善效果更加显著[20-22]。患有高胆固醇血症的成年男性,每天摄入含有3 g SPHP的保健食品,连续服用3个月,可观察到血清胆固醇显著降低[21]。美国食品药物管理局(Food and Drug Administration,FDA)和其他机构的人体研究也表明,通过对SPHP的有效剂量换算,成人每天摄入25 g大豆蛋白源性肽能显著降低血脂[21]。
鉴于大豆蛋白源性肽拥有上述的胆固醇调节功能,从大豆蛋白中如何获得有活性的大豆蛋白源性肽[11,23],以及大豆源性肽在降低胆固醇方面的作用机制,已成为现在植物源性肽研究的热门领域。
2.2 大豆蛋白源性肽对甘油三酯的调节作用
甘油三酯,又称脂肪,是由甘油和脂肪酸组成的,脂肪酸的种类和长短不同,脂肪酸又分为饱和脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸。高甘油三酯血症比单纯的高胆固醇血症更为常见,流行病学研究表明,高甘油三酯血症也是冠心病、动脉粥样硬化等疾病的诱因。大豆蛋白源性肽具有抑制脂肪酸合成、降低甘油三酯含量和预防肥胖等生物学活性[24]。
2.2.1 抑制脂肪酸合成抑制脂肪酸的合成重要的是抑制脂肪酸合成酶的活性。脂肪酸合成酶(fatty acid synthase,FAS,EC 3.2.1.85)是一种多组分酶,通过NADPH依赖的循环反应催化长链脂肪酸的生物合成。FAS为同型二聚体,每条多肽链带有7个催化域,整合了脂肪酸合成所需的所有步骤。FAS抑制剂的发现和开发,可以预防肥胖、代谢紊乱以及癌症等疾病的发生[25-26]。通过对中枢神经系统中FAS的抑制,可显著减少动物的食物摄入和体质量[27]。抑制下丘脑和胰腺β细胞中的FAS,可保护小鼠免受高脂肪饮食诱导的代谢综合征[28]。
研究发现,肥胖大鼠模型中,通过摄入大豆蛋白源性肽,可有效抑制脂肪酸合成和肥胖的发生[29]。进一步研究发现,大豆蛋白源性肽中的3种肽(KNPQLR、EITPEKNPQLR和RKQEEDEEQQRE)可以抑制FAS活性[30]。这3种肽除了具有抑制FAS的活性,还有减少机体前脂肪细胞(3T3-L1脂肪细胞)中脂质积聚(r=0.7)的作用。大豆蛋白源性肽对FAS的抑制效力,也与其分子量、pI值、所带负电以及亲水残基的数量相关。通过分子模拟预测发现,分子量较大的FAS抑制肽(EITPEKNPQLR和RKQEEDEEQQRE)和低于经典硫酯酶抑制剂(奥利司他)的相互作用与人类FAS的硫酯酶结构域结合。进一步的研究表明,大豆蛋白源性肽通过与FAS硫酯酶结构域中的疏水沟槽相互作用,阻断FAS活性位点。人工合成的EITPEKNPQLR和RKQEEDEEQQRE显示的FAS硫酯酶抑制活性,高于竞争性FAS抑制剂C75的活性,但低于奥利司他。
体外实验显示,大豆蛋白源性肽中含有的赖氨酸—丙氨酸(KA)、缬氨酸—赖氨酸(VK)和丝氨酸—酪氨酸(SY)成分,能抑制人肝HepG2细胞中乙酸合成甘油三酯,但这些二肽在体内的作用机制尚不清楚[30]。
2.2.2 降低甘油三酯的作用在动物体内实验中观察到,OLETF大鼠摄入大豆蛋白源性肽(含有约80%分子量为500或更小肽的混合物)后,可降低肝脏中的甘油三酯水平。同时还比较了大豆蛋白源性肽和酪蛋白水解物(动物蛋白源性肽)的差异,通过离体肝脏灌注法,大鼠在摄入大豆蛋白源性肽后,肝脏分泌的甘油三酯低于摄入的酪蛋白,这表明大豆蛋白源性肽在降低体内血清中的甘油三酯水平方面具有明显的优势[31]。
大豆蛋白源性肽可以促进小鼠3T3-L1细胞中脂联素的表达和分泌[32-33]。脂联素(adiponectin,ADPN)是脂肪细胞分泌的一种内源性生物活性多肽或蛋白,它通过促进过氧化物酶体增殖物激活受体γ(peroxisome proliferator activated receptorγ,PPARγ)的表达,进而促进脂肪细胞分化,并发挥抗糖尿病和抗动脉硬化作用。PPARγ具有脂肪组织特异性,能被脂肪酸及外源性过氧化物酶体增殖剂激活,以调控某些参与脂质代谢的酶的表达,是调节目标基因表达的核内受体转录因子一类超家族成员。
此外,脂肪细胞中甘油三酯的脂解反应,被认为是治疗肥胖的潜在靶点。据报道,从大豆蛋白源性肽中提取的异亮氨酸-亮氨酸-亮氨酸(Ile-Leu-Leu,ILL)、亮氨酸-亮氨酸-亮氨酸L(Leu-Leu-Leu,LL)和缬氨酸-组氨酸-缬氨酸-缬氨酸(Val-His-Val-Val,VHVV)具有刺激脂解反应的活性[13]。在饲喂高脂饮食的小鼠中,腹腔注射VHVV可降低血浆中的甘油三酯和低密度脂蛋白含量,具有抗肥胖作用[2]。一些植物蛋白源性肽具有抑制食欲的减肥作用[34-35]。胆囊收缩素受体1型(cholecysto kinintypel receptorⅠ,CCK1R)是一种位于动物胃肠道的G蛋白偶联受体,植物蛋白源性肽可以作为配体,从而激活受体,导致饱腹感。因此,大豆蛋白源性肽可以激活CCK1R途径,作为肥胖的有效治疗策略[36]。
因此,研究者认为,每天摄入5 gβ-伴大豆球蛋白可降低人体血清甘油三酯和内脏脂肪水平,减少脂肪细胞中脂肪的积累[13]。美国食品药物管理局和其他机构的人体研究也表明,通过SPHP的有效剂量换算,成人需要每天摄入25 g大豆蛋白可以显著降低血脂水平[5]。
3 大豆蛋白源性肽对糖代谢的调节作用
由于葡萄糖无法穿过细胞膜的脂质双层,需要在葡萄糖转运蛋白(glucose transporter,GLUT)和钠-葡萄糖协同转运蛋白(sodium-dependent glucose transporters,SGLT)协同作用下才能被摄取[37]。因此,GLUT和SGLT是改善糖尿病的重要靶点。体外实验表明,大豆蛋白源性肽中AKSPLF、ATNPLF、FEELN和LSVSVL降低了Caco-2细胞对葡萄糖的摄取。分子对接研究还表明,这些肽与葡萄糖转运蛋白SGLT-1和GLUT-2相互作用后,阻断了SGLT-1和GLUT-2对葡萄糖的运输能力[38],在高血糖大鼠模型中,上述大豆蛋白源性肽降低了机体的血糖水平[39]。
2型糖尿病的发生是由于胰岛素抵抗或胰岛素分泌不足,从而无法维持正常的血糖水平。胰安肽(aglycin)是从大豆中分离的由37个氨基酸组成的大豆蛋白源性肽。对高糖高脂膳食和链脲佐菌素诱导2型糖尿病小鼠模型,口服摄入胰安肽后,可以促进胰岛素受体信号通路相关蛋白的表达(增加胰岛素受体和膜GLUT-4的表达),最终降低血糖浓度[40]。据报道,在GK大鼠(非肥胖型2型糖尿病大鼠模型)实验中,大豆蛋白源性肽的摄入,可有效改善胰岛素抵抗作用,最终降低机体的血糖水平[41]。
此外,在KKAy小鼠(一种2型糖尿病小鼠模型)实验中,通过饮用水途径摄入抗焦虑肽酪氨酸-脯氨酸-苯丙氨酸-缬氨酸-缬氨酸(Tyr-Pro-Val-Val,YPFVV),一种源自大豆蛋白源性肽的肽类,也可以降低血糖水平。研究者认为,这种大豆蛋白源性肽的摄入,可以提高胰岛素反应性GLUT4的表达水平,并促进胰岛素对于葡萄糖的摄取[42],从而降低小鼠体内的血糖水平。血糖水平的降低可能与脂肪β-氧化的激活以及脂联素和PPARα增加能量消耗有关[43],但促进脂联素表达和分泌的靶基因尚未发现。另外,糖尿病与抗焦虑肽之间的关系以及糖尿病发生与阿片受体的激活之间的关系尚不清楚。
由于二肽基肽酶IV(DPP-IV)、α-淀粉酶和α-葡萄糖苷酶是直接参与血糖调节的关键酶,抑制这些酶活性的肽是治疗2型糖尿病的有效策略[44]。其中,二肽基肽酶(di-peptidyl peptidase IV,DPP-IV)是一种存在于血液和细胞膜上的酶,可以促使肠促胰岛素激素、胰高血糖素样肽-1(glucagon like peptide-1,GLP-1)和胃抑制多肽(gastric inhibitory polypeptide,GIP)失活。据报道,大豆蛋白源性肽(IAVPTGVA、LTFPGSAED)可抑制DPP-IV活性[45]。因此,近年来,来源于天然替代品(大豆蛋白源性肽)的DPP-IV抑制肽,在治疗2型糖尿病中已成为人们关注的焦点。
4 其他植物源性肽在糖脂代谢方面的调节作用
4.1 胆固醇代谢
除了大豆蛋白源性肽具有调节糖脂代谢的作用,据报道,其他植物蛋白源性肽也具有这方面的活性(表1,2)。一项从豇豆蛋白水解物中提取、鉴定出的GCTLN寡肽和QDF寡肽,在体外具有抑制3-羟基-3-甲基戊二酰辅酶A(3-hydroxy-3-methylglutaryl coenzyme A,HMG-CoA)还原酶活性,HMGCoA还原酶是肝细胞合成胆固醇过程中的限速酶,抑制HMG-CoA还原酶能阻碍胆固醇的合成[46]。
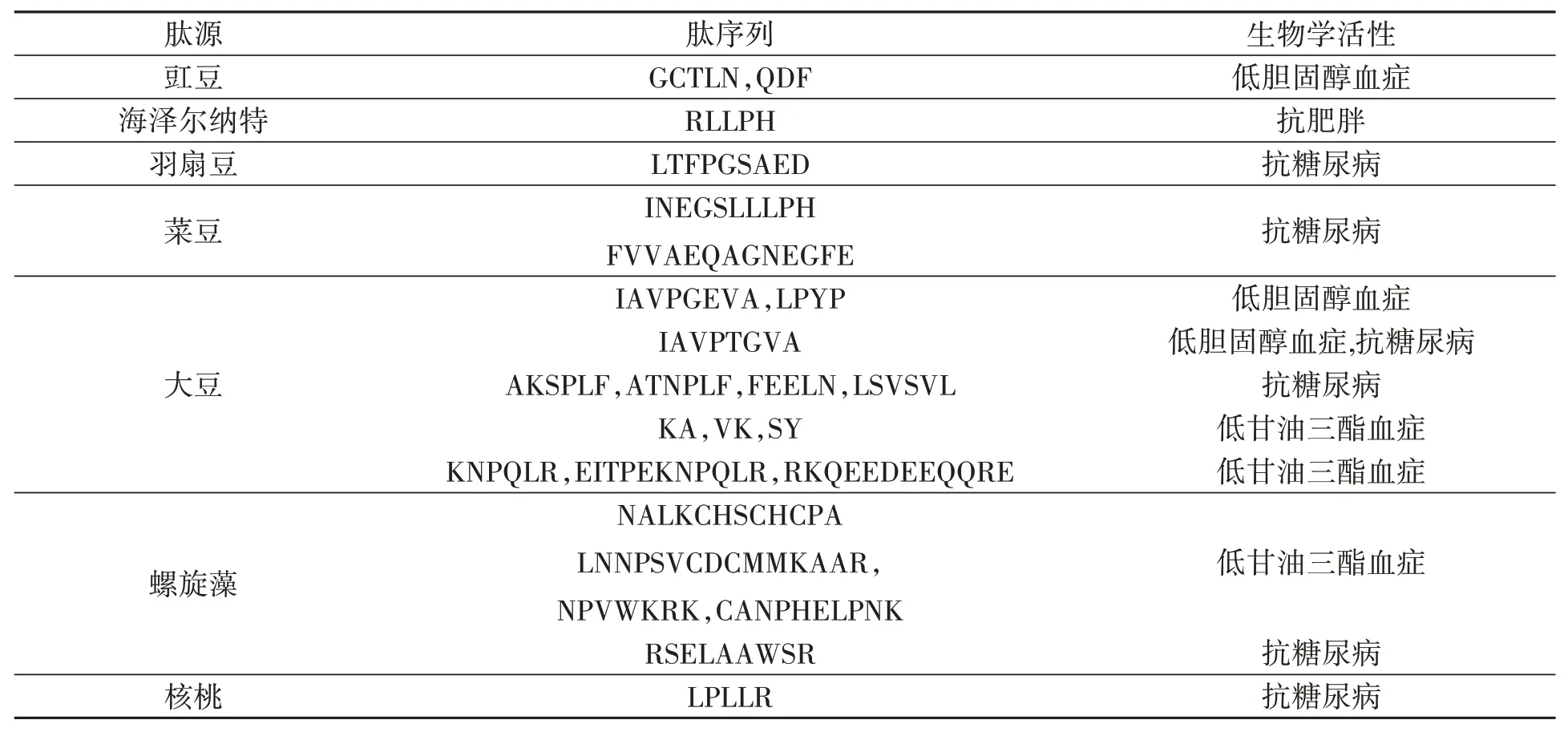
表1 体外活性代谢调节的植物肽Table1 Plant peptides regulating active metabolism in vitro

表2 体内活性代谢调节的植物肽Table2 Plant peptides regulating active metabolism in vivo
此外,小麦面筋蛋白水解物对大鼠也有降低胆固醇的作用[47]。与酪蛋白胰蛋白酶水解物(casein tryptic hydrolysate,CTH)相比,大鼠摄入结合磷脂的小麦面筋水解物复合物(wheat gluten hydrolysate with pospholipids,WGHP)或小麦面筋水解物(wheat gluten hydrolysate,WGH)后,能显著降低血清和肝脏胆固醇水平[48]。WGHP的胆汁酸结合能力显著高于WGH,WGH胆汁酸结合能力高于CTH。研究也发现,与CTH相比,WGH或WGHP存在时,胆固醇的胶束溶解度显著降低。饲喂WGH或WGHP大鼠的粪便中类固醇排泄量显著高于CTH大鼠[48]。这些结果表明,WGH或WGHP抑制胆固醇胶束溶解度导致胆固醇吸收的抑制,胆汁酸的高结合能力促进了胆固醇的代谢,降低血浆中胆固醇的含量。
4.2 甘油三酯的代谢
从钝顶螺旋藻中提取的NALKCHSCPA、LNNPSVCDCMMKAR、NPVWKRK和CANPHELPNK 4种肽,对小鼠3T3-L1前脂肪细胞具有抑制作用[49],其中NPVWKRK和CANPHELPNK体外结果显示,对胰脂肪酶活性有抑制作用,从而减少甘油三酯的积累[49]。从榛子蛋白水解物中提取的RLLPH寡肽,通过与PPARγ、C/EBPα、aP2 FAS、SREBP1c、ACC1和HMGCR等蛋白的相互作用,能够在小鼠3T3-L1前脂肪细胞中减少脂肪生成[50]。另一项研究表明,鹰嘴豆蛋白水解物中提取的VFVRN肽在高脂饮食喂养的小鼠中有降血脂作用[51]。
据报道,黑豆蛋白源性肽摄入后,肽类通过瘦素信号转导子和转录激活子3(signal tranducers and activators of transcription,STAT3)的磷酸化和AMP依赖的蛋白激酶[adenosine 5-monophosphate(AMP)-activated protein kinase,AMPK]激活,再辅助合理的运动,可以有效降低食欲和高脂饮食所诱导的体质量增加[52-53]。
4.3 糖代谢
向大鼠饲喂添加小豆蛋白源性肽的水可降低餐后的血糖水平。在小豆[54]和孜然种子[55-56]中发现了一些α-淀粉酶抑制肽。核桃蛋白碱性蛋白酶水解物中的LPLLR肽,在体外具有对α-淀粉酶的抑制活性,并减轻HepG2细胞的胰岛素抵抗[57]。从豌豆发酵中获得的ACE和α-淀粉酶抑制活性物质,抑制活性最高的肽为INEGSLPH和FVVAEQAGNEGFE[58-59]。此外,在钝顶螺旋藻中发现了一种DPP-IV抑制肽(RSELAAWSR)[60-61]。这些肽类的发现,为治疗和预防糖尿病的发生提供了新的选择。
5 展望
本文综述了大豆蛋白源性肽在调节糖脂代谢过程中的体内外活性等方面的研究,同时还综述了可以作为糖脂代谢调节的其他候选的植物源性肽[62-63]。未来,关于大豆蛋白源性肽是否具有体内调节糖脂代谢还需要更进一步的功能研究,包括动物实验和临床研究。其中,将植物尤其是大豆蛋白源性肽中具有体外生物活性的肽应用于疾病管理,应侧重于生物活性和分子机制的研究,包括肠-脑-肝轴稳态和肽代谢,体内肽转运体等[64-66]。此外,大豆蛋白源活性肽的理化性质,取决于氨基酸的性质、疏水性/亲水性氨基酸在肽链中的序列,以及距离N和C末端的远近等[37]。目前,关于大豆蛋白源性肽的结构和生理调节功能之间的关系尚不十分清楚,但肽的结构是肽生物活性的重要因素[37],这些问题对于进一步推广大豆蛋白源性肽在调节糖脂代谢应用至关重要。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
锦州医科大学报(2021年8期)2021-11-18
中西医结合肝病杂志(2020年2期)2020-10-27
中国中医急症(2019年10期)2019-05-21
祝您健康(2018年12期)2018-11-27
中国油脂(2017年12期)2017-01-18
中国卫生标准管理(2015年16期)2016-01-20
川北医学院学报(2015年5期)2015-12-05
中国医疗美容(2015年1期)2015-07-12
中医研究(2014年8期)2014-03-11
