
高直链淀粉的生物合成和应用研究进展
2023-02-23 05:22刘雨霏罗慧琳潘云云申雨韩张浩宇姚长洪
化学与生物工程 2023年2期
刘雨霏,罗慧琳,陈 响,潘云云,申雨韩,张浩宇,姚长洪
(四川大学化学工程学院,四川 成都 610065)
淀粉作为一类可再生的物质一直备受关注,它在自然界中的来源广泛、含量丰富,并且具备良好的性能,在制药、食品、材料等领域都有广泛的应用。淀粉由直链淀粉(amylose,Am)和支链淀粉(amylopectin,Ap)组成,其中直链淀粉主要由葡萄糖残基通过α-1,4糖苷键线性连接而成,支链淀粉在线性分子链的基础上还存在通过α-1,6糖苷键形成的大量侧链,其分子量较直链淀粉的更大[1]。直链淀粉和支链淀粉所占的比例决定了淀粉的性质和品质[1]。普通淀粉一般由20%~30%的直链淀粉和70%~80%的支链淀粉组成[2],而经过基因工程育种[3]后,可以得到直链淀粉含量达到55%~90%的高直链淀粉(high amylose starch,HAS)[4-5]。与普通淀粉相比,高直链淀粉的颗粒形貌不同,并具有糊化温度较高、成膜性能较好等特殊的理化性质,因此具有更广阔的应用前景[6]。作者在此从生物合成、理化性质、应用等三方面综述高直链淀粉的研究进展,为高直链淀粉的研究、产品开发和工业应用提供参考。
1 高直链淀粉的生物合成
1.1 淀粉的合成
高等植物中的淀粉可分为暂时性淀粉(transient starch)和贮藏性淀粉(storage starch)两种,分别在光合组织的叶绿体和储存组织的淀粉体中合成[7]。日间利用光合作用合成的暂时性淀粉在夜间会降解为代谢所需的碳源和能量,运输到淀粉体合成贮藏性淀粉[7]。贮藏性淀粉为种子的生长发育提供能量和营养,在种子萌发等生命活动中起重要作用。
1.1.1 淀粉合成途径
植物以蔗糖为碳源进行贮藏性淀粉的生物合成,如图1所示。
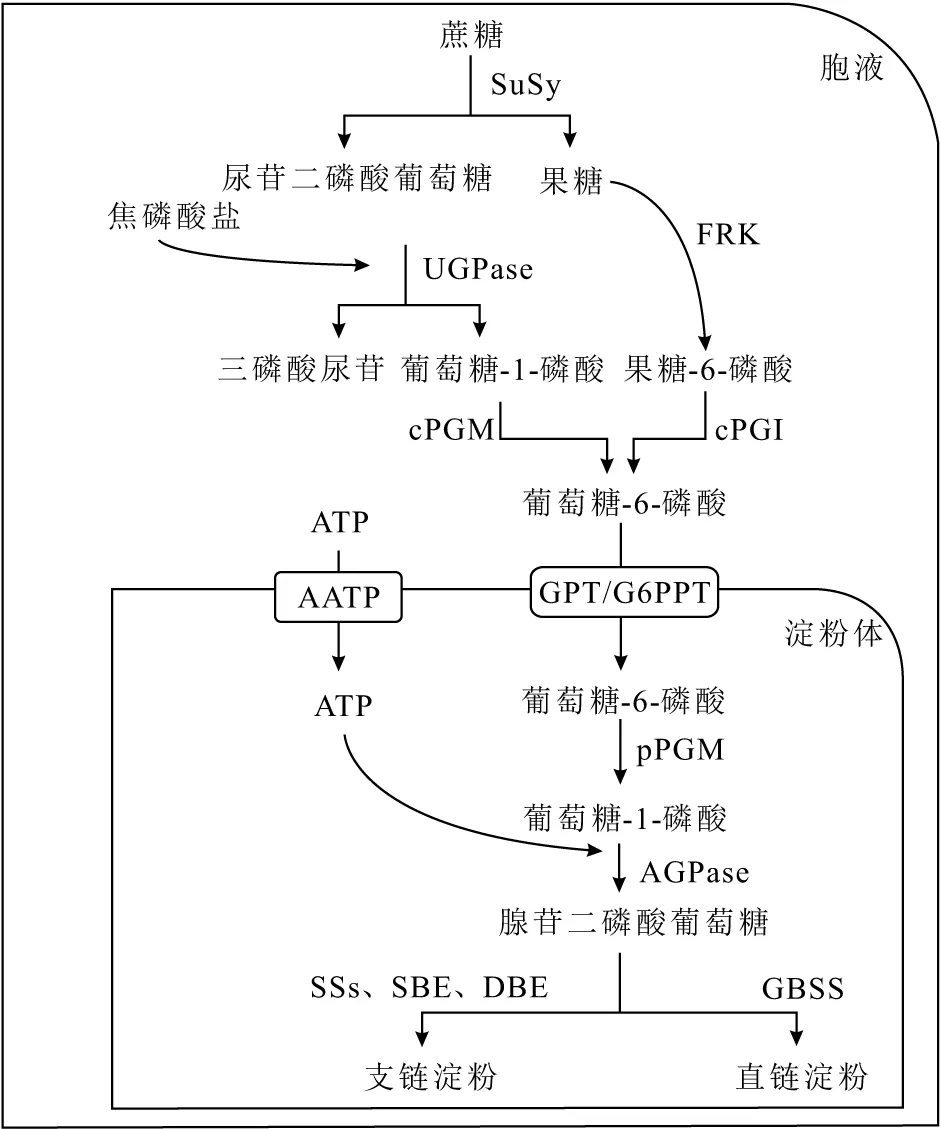
图1 贮藏性淀粉的生物合成途径Fig.1 Biosynthesis route of storage starch
蔗糖合成酶(SuSy)利用尿苷二磷酸(UDP)催化裂解蔗糖为尿苷二磷酸葡萄糖(UDPG)和果糖,尿苷二磷酸葡萄糖和胞液中的焦磷酸盐(PPi)在尿苷二磷酸葡萄糖焦磷酸化酶(UGPase)的作用下生成葡萄糖-1-磷酸(G1P)和三磷酸尿苷(UTP),胞质磷酸葡萄糖酶(cPGM)又将G1P代谢为葡萄糖-6-磷酸(G6P);果糖则在果糖激酶(FRK)的作用下形成果糖-6-磷酸(F6P),再借助胞质磷酸葡萄糖异构酶(cPGI)转化为G6P。通过尿苷二磷酸葡萄糖及果糖两种途径生成的G6P借助葡萄糖-6-磷酸盐/磷酸转运蛋白(GPT/G6PPT)进入淀粉体;同时,三磷酸腺苷(ATP)也在ATP/ADP转运蛋白(AATP)的帮助下进入淀粉体。在淀粉体中,质体磷酸葡萄糖变位酶(pPGM)将G6P再次转化为G1P,随后质体ADP-葡萄糖焦磷酸化酶(AGPase)又将G1P和转运来的ATP转化为腺苷二磷酸葡萄糖(ADPG)作为淀粉合成的前体[8]。直链淀粉由颗粒结合淀粉合成酶(GBSS)催化腺苷二磷酸葡萄糖与葡萄糖糖链的非还原末端连接,使糖链延长而得到;支链淀粉的合成则需要可溶性淀粉酶(SSs)、淀粉分支酶(SBE)和去分支酶(DBE)的共同作用。其中,SSs催化α-1,4葡聚糖链的延长;SBE则催化α-1,4糖苷键断裂,所得短链的C1位再在相同或另一链中葡萄糖基的C6位形成α-1,6糖苷键,形成分支;DBE能够修饰不规则的糖链分支,形成具有晶型结构的支链淀粉。
1.1.2 淀粉合成关键酶
SuSy是一种重要的可逆酶,它能实现蔗糖和尿苷二磷酸向尿苷二磷酸葡萄糖和果糖的可逆转化,促进蔗糖的合成与降解,从而影响植物细胞壁组分和淀粉合成[9]。SuSy对蔗糖的代谢有一定的影响,当活性降低时,淀粉合成受阻,且合成的淀粉含量下降。玉米SuSy的活性与zmsh1、zmsus1和zmsus3基因的表达相关。zmsh1是一个重要的蔗糖合成酶编码基因,SuSy的活性随着zmsh1基因表达量的升高而降低;zmsus1基因在玉米的根、茎、叶、胚中都有所表达,是淀粉合成前体尿苷二磷酸葡萄糖的供源;zmsus3在胚乳、胚珠、根和幼芽中表达[10]。
AGPase是淀粉合成中的一个重要限速酶,其作用机制是通过促进G1P、ATP向腺苷二磷酸葡萄糖和焦磷酸盐的转化,从而实现对谷类胚乳及其它器官淀粉含量的调控[1]。在绝大多数植物中,AGPase仅存在于叶绿体和淀粉体(合称为质体)中,但在禾谷类植物胚乳细胞中只有15%为质体型AGPase,其余均为胞质型AGPase,而胞质型AGPase在催化腺苷二磷酸葡萄糖生成的同时还参与细胞内糖的代谢调控。玉米中质体型AGPase有AGPLLZM、AGPSLZM、AGPLEMZM、AGPSEMAM等4种亚基,其中AGPSLZM与AGPLEMZM能够构成有活性的腺苷二磷酸葡萄糖焦磷酸化酶(ADPGase)[11]。有研究表明,AGPase的活性直接影响淀粉含量,如在马铃薯中AGPase的活性与淀粉含量成正比[12]。在玉米籽粒胚乳中AGPase由2个大亚基和2个小亚基组成,分别为结构调节中心和酶催化中心,其对应编码基因sh2和bt2的变异均能使AGPase活力下降15%~20%,淀粉合成受到阻碍,淀粉含量下降75%[1]。但也有研究表明,AGPase的活性与淀粉含量没有显著联系,它的活性变化与淀粉积累速率变化并不一致,可能原因是合成淀粉的供应途径不止一条[13]。
淀粉合成酶(SS)由GBSS和SSs组成,通过在葡聚糖引物的非还原末端引入葡萄糖残基延长链长度。GBSS和淀粉颗粒紧密地结合在一起,主要参与直链淀粉合成,此外还参与超长链(ELCs)的支链淀粉合成[14]。GBSS可分为两类:GBSSⅠ和GBSSⅡ。GBSSⅠ是催化腺苷二磷酸葡萄糖合成直链淀粉的重要酶,在胚乳、花药中所占比例较高,其编码基因为wx。在糯玉米中,由于wx基因的变异,GBSSⅠ酶活力丧失,在胚乳中仅能合成支链淀粉[1]。在水稻胚乳中,GBSS由第6染色体wx基因编码,负责直链淀粉的合成[15]。GBSSⅡ与GBSSⅠ具有相同活性,但主要存在于瞬态器官中,如在叶片中参与直链淀粉的合成[16]。SSs有SSⅠ、SSⅡ、SSⅢ、SSⅣ、SSⅤ等5种亚型,主要负责支链淀粉链的延长[17]。SSⅠ对直链淀粉含量没有显著影响,主要影响水稻籽粒胚乳中支链淀粉的结构[18]。玉米中SSⅡ有SSⅡa、SSⅡb、SSⅡc等3种类型,其中,SSⅡa在胚乳中高表达,SSⅡb主要在叶片中高表达,SSⅡc在叶片、胚乳和根中均有表达;SSⅡb和SSⅡc缺乏突变体,对其功能的研究尚未有进展[19]。SSⅡa的编码基因为su2,SSⅢ的编码基因为du1,这两个基因突变会分别使SSⅡa和SSⅢ的活性降低,导致胚乳中的直链淀粉含量升高[1]。SSⅣ控制淀粉颗粒的起始形成,它的过表达使拟南芥叶片中淀粉含量升高30%~40%[20]。SSⅤ是一种不具有催化葡萄糖基转移酶功能的非典型亚型,其与SSⅣ类似,具有与糖链结合和与类肌球蛋白的叶绿体蛋白(MRC)相互作用的保守区域,控制淀粉颗粒的起始形成,进而影响拟南芥叶片中淀粉颗粒的大小和数量[17]。
SBE可以使原本的α-1,4糖苷键断裂,或在α-1,4糖苷键的帮助下实现受体链C6位与切下的短链相连接。SBE有SBEⅠ、SBEⅡ等2种亚型,SBEⅡ又有2种类型:SBEⅡa和SBEⅡb。在禾谷类植物中,SBEⅠ和SBEⅡa主要在胚乳中表达,SBEⅡb则在水稻的所有器官中表达[21]。此外,SBEⅡb是主要的支链淀粉合成酶,SBEⅡb突变的玉米中的直链淀粉含量明显升高,但淀粉总量下降20%,且突变体胚乳没有光泽,籽粒晶莹剔透,有一定程度的褶皱[1]。SBEⅠ突变对玉米直链淀粉的合成有一定影响,但不影响支链淀粉的结构[1]。
DBE的功能主要是通过水解支链淀粉的α-1,6葡聚糖对支链进行微细的结构修饰,这样能消除不规则的主链分支,从而使淀粉的螺旋状结构更易结晶。DBE包括异淀粉酶(ISA)和普鲁兰酶(PUL)[22]。ISA水解支链淀粉和可溶性植物糖原的α-1,6糖苷键,主要有ISA1、ISA2、ISA3等3种亚型,ISA1对不规则的支链淀粉分支进行修剪,使其突变株中的淀粉含量下降;ISA2只有与ISA1形成异源复合体时才具有催化功能;ISA3与糖原积累有关。PUL主要作用对象是普鲁兰糖和支链淀粉,在谷物萌发时的淀粉降解中起作用。
1.2 提高直链淀粉含量的途径
通过培育不同的高直链淀粉作物,可以扩大高直链淀粉品种的选择范围。目前高直链淀粉主要来源于玉米、大麦、水稻等作物,在市场上竞争力大,商业价值高。提高作物的直链淀粉含量可从增强GBSSⅠ表达、抑制SSs活性、抑制SBE活性和抑制葡聚糖水二激酶(GWDs)活性等几方面着手,如表1所示。
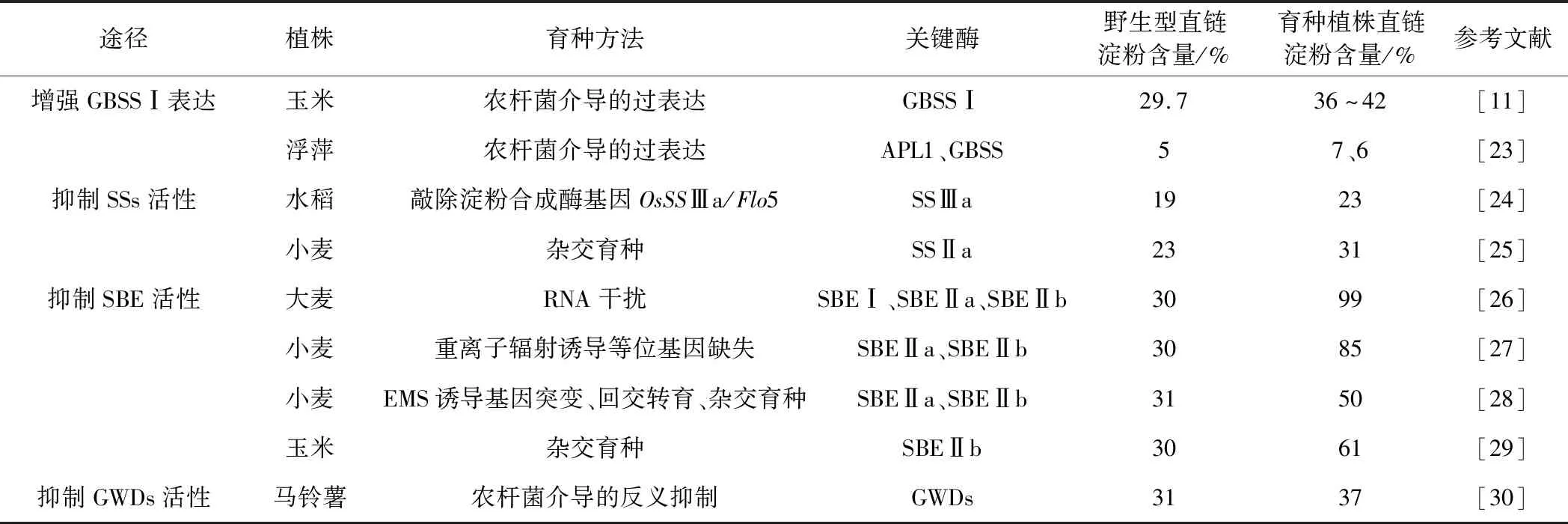
表1 提高直链淀粉含量的途径Tab.1 Methods for increasing amylose content
1.2.1 增强GBSSⅠ表达
GBSSⅠ直接负责胚乳中直链淀粉的合成。GBSSⅠ也被称为Waxy蛋白,能够催化淀粉糖链的延长。增强GBSSⅠ的表达可以使淀粉中直链淀粉的含量显著升高。赵丫杰[11]将编码GBSSⅠ的zmwx基因与质粒的融合基因 “p27kd::zmwx” 转入玉米自交系得到转基因纯合的子代T3植株,其直链淀粉含量升高了9%~11%。浮萍的转基因株系GBSS-21和GBSS-24的直链淀粉含量较野生型分别升高了40.4%和43.7%[23]。
1.2.2 抑制SBE活性
编码SBEⅡb的sbeⅡb基因、编码SSⅢ和SBE的du1基因、编码SSⅡ的su2基因的变异均可导致相应酶活力下降,支链淀粉合成能力下降的同时直链淀粉含量升高。SBEⅡ突变株又称ae突变体,其直链淀粉含量能达到61.7%~67.7%[31]。ae基因显性突变基因ael-5180通过回交转育导入优良玉米品种自交系可以提高直链淀粉的含量[29]。降低小麦中SBEⅡa和SBEⅡb的活性能使分支减少、直链淀粉含量升高[32]。sbeⅡ与其它基因(如ssⅢa)组合突变株的直链淀粉含量较单基因突变株的更高[33],如籼稻中sbeⅡb与sbeⅠ基因组合突变株的直链淀粉含量从单一sbeⅡb基因突变株的27.2%提高到60.0%[34]。抑制马铃薯的SBEⅡa和SBEⅡb活性,能将直链淀粉含量由28%提高到72%[35]。
1.2.3 抑制GWDs活性
在玉米中,GWDs不仅能催化叶绿体中暂时性淀粉的降解,还与贮藏性淀粉的磷酸化有关[36]。GWDs可以催化支链淀粉葡萄糖残基C6或C3位与ATP上γ-磷酸基团水解后的β-磷酸盐相结合,破坏淀粉表面的颗粒结构,使其易于与淀粉合成相关酶结合[37]。GWDs表达降低会使淀粉中支链淀粉含量降低,从而提高直链淀粉的相对含量。与野生型马铃薯淀粉比较,抑制GWDs活性的马铃薯淀粉中的直链淀粉相对含量提高了5%~6%[30]。但GWDs影响直链淀粉的合成机制尚不清楚。
2 高直链淀粉特殊的理化性质
2.1 颗粒形貌
淀粉的存在形式为颗粒状。根据颗粒直径的大小,淀粉颗粒可分为A型(大颗粒)和B型(小颗粒),其中,A型的直径≥10 μm,B型的直径<10 μm[38]。在颗粒形貌上,高直链淀粉与普通淀粉呈现出明显的差别,普通淀粉颗粒多为饱满光滑的小球体或多面体,而高直链淀粉颗粒则多为没有蜡质光泽的不规则形状。李志伟等[39]通过对不同直链淀粉含量的淀粉颗粒进行扫描电子显微镜表型特征检测,发现在普通玉米淀粉中,A型和B型淀粉颗粒均是带有明显蜡质光泽、饱满光滑的小球体或多面体;而在直链淀粉含量为55.53%的高直链玉米淀粉中,A型淀粉颗粒出现皱缩、扭曲的情况,B型淀粉颗粒则出现拉长的情况,二者表面均带有蜡质光泽;随着直链淀粉含量的上升,A型淀粉颗粒皱缩、扭曲的情况进一步加重,颗粒直径也出现减小的情况,B型淀粉颗粒则出现扭曲的情况,并且二者表面没有明显的蜡质光泽。
2.2 糊化温度
淀粉的糊化过程是指水分充足的淀粉加热到一定温度时,淀粉颗粒及其晶体结构开始破坏,出现体积膨胀、黏度迅速增大的现象[5]。Liu等[40]通过研究不同直链淀粉含量的玉米淀粉的糊化过程,发现直链淀粉含量较低的蜡质玉米淀粉和普通玉米淀粉在约70 ℃时便开始出现大量的糊化吸热,而直链淀粉含量更高的淀粉则在80 ℃以上时才开始发生糊化,因此随着直链淀粉含量的上升,往往糊化温度也会升高。然而,与普通淀粉相比,高直链淀粉的糊化温度虽然明显较高,但是在糊化过程的热流变化却比普通淀粉更加平缓,糊化焓也相对更低[40]。
2.3 老化性质
淀粉的老化是指部分或完全糊化的淀粉在冷却时发生的淀粉分子由无序到有序的变化过程[41]。淀粉的老化性质与其分子结构紧密相关,一般而言,直链淀粉比支链淀粉更容易发生老化[3],不同链长的直链淀粉发生老化的速率也有所不同。在淀粉的老化过程中,最先发生的是直链淀粉分子的快速重结晶,之后发生支链淀粉分子的缓慢重结晶[41]。淀粉分子结构是影响其老化过程的重要因素,决定了老化速率以及所形成淀粉微晶的结构[42]。通常情况下,高直链淀粉具有更快的老化速率。
2.4 成膜性能
直链淀粉的长线性特征使之易于自我缔合[43],其含量的升高将有利于增强淀粉的成膜性能。直链淀粉可以形成拉伸性能较好的内聚膜,被认为是一种良好的成膜剂,而支链淀粉所形成的膜则较易破裂[44]。Menzel等[44]使用透射电子显微镜对不同的淀粉薄膜进行检测,发现支链淀粉形成的膜结构孔径较小,约为10 nm,并且多为圆形小孔;而直链淀粉形成的膜结构孔径可达60 nm,并且多为不规则小孔,分布也更不均匀。与普通淀粉相比,高直链淀粉不仅具有更好的成膜性能,其所形成的膜还具有更优良的阻隔性和拉伸性能。
2.5 抗消化性
抗性淀粉(resistant starch,RS)是指在人体小肠中难以被降解,在体内消化、吸收都较缓慢的淀粉。抗性淀粉一般有5个亚型RS1~RS5,其中高直链淀粉可作为RS2的一种,是一类天然抗性淀粉[45]。杨光等[46]发现,直链淀粉可以参与抗性淀粉的形成,并且直链淀粉的含量与抗性淀粉的产率呈明显的正相关,随着直链淀粉含量的升高,抗性淀粉含量也大幅升高。
3 高直链淀粉的应用
3.1 制药工业
高直链淀粉具有无毒性、水溶性、生物可降解性等特点,可作为缝合线、绷带等应用于医疗行业[6]。基于其良好的生物相容性,可用于构建药物载体,并通过结构改造等手段改变淀粉的水解速率,以控制药物的释放、扩散,可发挥缓控释作用,达到提高药物的生物利用度并降低药物毒副作用的目的。Zhang等[47]采用酸化法合成了直链淀粉-布洛芬(Ibuprofen,IBF)包合物,直链淀粉与布洛芬之间存在较强的相互作用,通过直链淀粉与布洛芬的复合可以在肠道中实现布洛芬的缓慢释放,提高布洛芬的疗效并且减轻其对胃肠道的副作用。Senthil等[48]将高直链淀粉化学改性为羧甲基高直链淀粉(CM-HAS),并以尼莫地平为模式药物,制备出CM-HAS骨架片,表明高直链淀粉可作为赋形剂构建口服药物控释体系。
3.2 食品工业
3.2.1 抗性淀粉:新型的膳食纤维
高直链玉米淀粉含有高达60%的抗性淀粉[1],同时,高直链淀粉也可用于高效生产抗性淀粉。抗性淀粉在大肠中经过微生物发酵,可以发挥传统膳食纤维的生理功能,并且其生产短链脂肪酸的能力较传统膳食纤维更高,是一种新型的膳食纤维。基于抗性淀粉难以被人体小肠和胃消化、吸收的特点,还可将其替代普通淀粉生产出低热量、低吸收的减肥食品[3]。
3.2.2 低脂食品
高直链淀粉可以与脂质作用形成复合物,替代全脂或高脂的材料生产更健康的低脂食品。在热处理等加工条件下,直链淀粉的链式结构会发生旋转,形成螺旋空腔结构,并且易于与其它分子的疏水基团或非极性部分发生络合,形成螺旋复合物[49]。Singh等[50]研究了高直链淀粉-起酥油复合物的物理特性、流变学性质、质地及储存等,发现该复合物可替代起酥油用于低脂蛋糕糖霜的制备。
3.2.3 休闲膨化食品
淀粉及其衍生物常用于膨化食品的生产。与普通淀粉相比,高直链淀粉具有更好的抗拉伸性能,可以增加食品的脆性[1],赋予膨化食品更好的口感。在食品生产过程中通过调整直链淀粉的含量也可以生产出不同膨化效果的食品[51]。
3.2.4 食品包装材料
利用高直链玉米淀粉可以制备出允许CO2透过、而O2和N2无法透过的可食用半透明纸,用于糖果、糕点等甜品的包装[1]。陈雪风等[52]利用高直链玉米淀粉制备出pH响应性的食品包装材料,该材料能够以颜色的变化为指示信号,有助于对食品的新鲜程度进行间接判断,实现对食品安全的实时监测。
3.3 材料工业
3.3.1 可降解薄膜及包装材料
淀粉是一种可再生、环境友好型自然资源。与普通淀粉相比,高直链淀粉通过聚合形成的透明、无味的膜具有更好的力学性能和阻隔性能[6]。但同时高直链淀粉所形成的薄膜具有脆性较大的缺陷,可通过将增塑剂与高直链淀粉混合对其进行改性来克服该缺点[6],起到增强膜的拉伸性能的作用,以利于高直链淀粉膜的加工和应用。孙炳新等[53]对比了不同增塑剂对高直链淀粉膜力学性能的影响,发现甘油、山梨醇对高直链淀粉膜具有较好的改性作用,而木糖的增塑效果相对较差,并且将甘油与山梨醇或木糖混合制成的复合增塑剂的改性效果要比单一增塑剂更好。
3.3.2 淀粉基可降解材料
由淀粉经过热塑加工制备出的淀粉基可降解塑料或将成为解决白色污染的一个有效途径[5]。淀粉基可降解塑料经过微生物发酵分解后产生的CO2和水对环境无污染,并且高直链淀粉基可降解塑料与普通淀粉相比具有更好的机械性能,有利于可降解塑料的广泛应用。Li等[54]研究了不同直链淀粉含量的淀粉基薄膜的挤压加工工艺以及薄膜性能,发现直链淀粉含量对淀粉基薄膜的微观结构、力学性能及热性能等都有较为显著的影响,高直链淀粉基薄膜表现出更好的机械性能,包括更高的模量、拉伸强度及冲击强度等。
4 结语
高直链淀粉具有较多不同于普通淀粉的理化性质,在制药、食品和材料等领域都有着十分广阔的应用前景。目前,高直链淀粉的生产及应用还面临着不少问题。在育种方面,高直链淀粉作物存在产率较低、培育周期较长的问题,而经过诱变育种或转基因育种的作物,则还要面对如何精准地改变基因或调控基因的表达、如何实现性状的稳定遗传、如何确保产品的安全性等问题[5]。在应用方面,高直链淀粉的高成本限制了其应用范围,而将高直链淀粉加工成材料的过程,也还存在加工工艺繁琐、材料性能不佳等问题[6]。此外,高直链淀粉对食品口感和品质有影响,比如增大了馒头、面包等食品的硬度和咀嚼度[55]。
随着相关研究的不断深入和生物技术的不断发展,针对上述问题已经有了一些解决思路和方法。CRISPR/Cas(clustered regularly interspaced short palindromic repeats/CRISPR-associated proteins)技术是近年来迅速发展的第三代基因编辑技术,代表性的CRISPR/Cas9系统可以精准地调控基因编辑[56],可能成为高直链淀粉植物育种的一条有效路线。Sun等[57]使用CRISPR/Cas9技术对水稻中的sbeⅠ和sbeⅡb基因进行靶向诱变,所得的sbeⅡ突变体中直链淀粉含量比野生型水稻显著提高,证明了使用CRISPR/Cas9对sbeⅡb进行基因编辑以获得高直链淀粉水稻的可行性。此外,发展高直链淀粉的非作物新来源也是解决生产问题的一条优良路线。微藻是一类可以通过光合作用固碳并积累淀粉、油脂等储能物质的微生物[58]。在营养限制等条件下,一些微藻会大量地积累淀粉[58-59];通过培养条件的优化和环境因子的调节,可以对微藻淀粉总量、直链淀粉含量等进行调控[60-62];尤其是海洋微藻生长迅速,培养可控,不占用耕地和淡水资源,在高直链淀粉的高效生产上具有广阔的应用前景。在高直链淀粉的应用方面,可以对高直链淀粉材料进行结构改造和性能改良,例如通过丙酰化处理,可增强高直链淀粉的热稳定性;通过羟丙基化处理,可增强高直链淀粉薄膜的柔韧性[6]。在改善食品口感和品质方面,可通过将高直链淀粉与普通淀粉、食品添加剂等复配使用[5],或是通过改变淀粉中直链淀粉的链长[63]等手段来实现。同时,优化出经济、高效且环保的高直链淀粉加工工艺,将有助于实现高直链淀粉的产业化应用。
随着高直链淀粉生产路线的升级、加工工艺的优化和材料性能的改良,高直链淀粉将广泛地应用于日常生活和工业生产中。
猜你喜欢
中华老年多器官疾病杂志(2022年10期)2022-11-21
生物加工过程(2020年3期)2020-06-17
中国现代药物应用(2020年6期)2020-01-10
中国粮油学报(2019年4期)2019-07-12
中西医结合心血管病杂志(电子版)(2018年8期)2018-01-12
中国塑料(2016年2期)2016-06-15
医学综述(2015年15期)2015-03-04
北京航空航天大学学报(2014年1期)2014-12-19
现代检验医学杂志(2014年1期)2014-02-06
中国机械工程(2012年15期)2012-07-25
