
添加玉米秸秆和根茬对不同肥力黑土微生物残体碳氮的影响
2023-03-09 08:41马南安婷婷张久明汪景宽
中国农业科学 2023年4期
马南,安婷婷✉,张久明,汪景宽
1沈阳农业大学土地与环境学院/土肥高效利用国家工程研究中心/农业农村部东北耕地保育重点实验室,沈阳 110866;2黑龙江省农业科学院土壤肥料与环境资源研究所,哈尔滨 150086
0 引言
【研究意义】作物秸秆和根茬是农田土壤有机质的重要来源。秸秆和根茬还田不仅可以增加土壤养分库容,而且可以提高微生物活性,被认为是培肥土壤的重要措施。土壤微生物利用外源有机质合成微生物生物量,然后以微生物残体的形式累积并稳定固存在土壤中。微生物残体是土壤碳源和氮源的中间过渡库,对土壤稳定有机质的形成具有重要的贡献[1]。因此,研究秸秆和根茬还田后土壤微生物残体碳氮的累积特征,对深入理解土壤有机质的形成和稳定的机制及土壤肥力提升具有重要的意义[2]。【前人研究进展】外源有机物添加促进了微生物残体的积累,提高了土壤有机碳(SOC)的稳定性[3-5]。氨基糖是土壤微生物残体的重要标识物。在可被定量的氨基糖中,氨基葡萄糖(GluN)主要来源于真菌几丁质,胞壁酸(MurA)唯一来源于细菌。由于氨基糖的相对稳定性和异源性,其含量被认为是微生物残体对SOC和全氮(TN)积累和贡献的可靠指标[2]。作物秸秆和根茬的化学组成存在高度异质性。作物秸秆一般具有较高比例的碳水化合物和较低的碳氮比,在土壤中优先分解;根茬则含有较高比例的木质素等难分解的化合物和较高的碳氮比,较难被微生物分解[6-7],因此秸秆和根茬添加到土壤后可能影响微生物的分解及同化过程[8]。微生物利用小麦秸秆合成氨基糖的速度大于利用根茬,微生物残体的形成受小麦残体类型影响显著[9]。玉米根茬和秸秆还田后,在分解前期添加秸秆处理较有利于微生物残体的积累,而在培养结束后(第500 天)添加根茬处理微生物残体的累积量及微生物残体碳占SOC的比例均较高[10]。但近来有研究却发现分子结构稳定的有机碳(例如木质素)在土壤中较易分解,拥有较快的周转速率[11-12]。外源底物添加促进了低肥棕壤真菌残体和高肥棕壤细菌残体的累积[10]。低碳氮比的土壤和无机氮的添加有利于微生物的合成代谢,促进了微生物残留物的形成和积累[13-16]。由此可见,微生物驱动土壤中外源有机物的分解和转化过程受外源有机物类型和土壤性质等因素的影响。【本研究切入点】我国东北黑土区农田土壤出现有机质含量下降、土层变薄等土壤退化问题,已引起广泛关注。秸秆和根茬还田被认为是黑土地区培肥土壤、作物增产增效的重要措施[17-19]。黑土中秸秆还田不仅促进了真菌的生长代谢,提高了表层土壤微生物量[20],而且改善了表层和亚表层土壤微生物群落结构和多样性,提高了土壤的固碳潜力[18]。李丽东等[21]通过室内培养试验发现,高有机质的黑土氨基糖累积数量高于低有机质的棕壤,而且秸秆分解有利于提高低有机质土壤的微生物生物量和底物利用效率,促进微生物残体的累积。黑土本身有机质含量较高,不同肥力水平黑土微生物如何响应不同类型外源有机物(秸秆和根茬)添加?微生物残体碳氮对SOC和TN的贡献如何?关于这个问题仍不很明确。【拟解决的关键问题】因此,本研究以东北不同肥力黑土为研究对象,结合13C和15N双标记方法,分析秸秆和根茬还田后外源碳氮在土壤中的累积动态,探讨不同类型外源有机物添加对不同肥力黑土微生物残体碳氮在土壤中累积的影响,量化微生物残体碳氮对SOC和TN的贡献,以期为东北黑土地区土壤有机碳库的稳定和氮库的扩容提供依据。
1 材料与方法
1.1 供试材料
供试土壤样品采自黑龙江省农业科学院哈尔滨黑土长期定位试验站(45°62′N, 126°27′E)。该试验站始建于1980年,所在地区属于北温带季风气候,年均温3.50℃,年均降水量533 mm,土壤类型为厚层黑土。本试验对两种不同肥力土壤进行培养试验。其中低肥土壤(LF)采自不施肥土壤;高肥土壤(HF)采自高量有机肥配施氮磷肥的土壤(年施氮肥 150 kg N·hm-2,磷肥 75.0 kg P2O5·hm-2,有机肥折合纯 N为 18.6 t·hm-2)(表 1)。

表1 不同肥力土壤的基本理化性质(2019年)Table 1 Basic characteristics of Black soil samples with different fertility levels (in 2019)
2019年9月下旬采集低肥和高肥土壤0—20 cm土层土壤样品,新鲜土样装入PVC盒用冷藏箱迅速带回实验室,剔除可见根茬和石块后风干过2 mm筛备用。
1.2 室内培养试验
共设置6个处理:(1)高肥土壤+13C15N双标记玉米根茬(HF+R);(2)高肥土壤+13C15N双标记玉米秸秆(HF+S);(3)低肥土壤+13C15N双标记玉米根茬(LF+R);(4)低肥土壤+13C15N双标记玉米秸秆(LF+S)。同时设置高肥和低肥土壤不添加秸秆和根茬的对照处理。每个处理3次重复。13C15N双标记玉米根茬的基本性质:全碳 444 g·kg-1、全氮 6.14 g·kg-1、C/N 为 72.4、δ13C 值 298‰、δ15N 值 11 003 ‰;13C15N双标记玉米秸秆的基本性质:全碳408 g·kg-1、全氮8.49 g·kg-1、C/N为48.0、δ13C 值 386‰、δ15N 值11 006‰。
首先将风干土壤样品(250 g,烘干重)避光预培养7 d(25℃,含水量为田间持水量的40%)。其次,分别将13C15N双标记的玉米根茬和秸秆(5 g,大小为2 mm)与预培养土壤(相当于2%归还量)充分混匀,调节土壤含水量至田间持水量的60%,在恒温恒湿条件下(25℃)避光培养,每周按称重法进行补水。在培养的第 30天和 180天分别从每个处理随机取出 3个培养瓶。土壤样品自然风干后研磨过100目筛,供分析SOC含量和δ13C值、TN含量和δ15N值、氨基糖含量。
1.3 分析方法
SOC和TN含量、δ13C和δ15N值采用元素分析仪——稳定同位素比例质谱仪(EA-IRMS,Elementar vario PYRO cube-IsoPrime100 Isotope Ratio Mass Spectrometer,德国)测定。
土壤中氨基糖含量采用盐酸水解,经纯化和衍生后利用气相色谱法测定[22]。即称取含有0.4 mg N的土壤样品,加入 10.0 mL HCl(6 mol·L-1),在 105 ℃下水解、过滤,加入100 μL 肌醇(内标1)蒸干后调节pH至6.6—6.8,离心后上清液再次蒸干,残留物质用无水甲醇溶解、离心后转移到衍生瓶中,N2吹干溶液,加入1mL蒸馏水和100 μL 内标2(N-甲基氨基葡萄糖,MGlcN),冷冻干燥后进行衍生,利用气相色谱进行测定(GC-7890B,Agilent,USA;DB-5毛细色谱柱30 m×0.25 mm×0.25 μm)。
1.4 数据处理
外源碳对SOC的贡献率(Fc,%)、外源氮对TN的贡献率(Fn,%)的计算公式如下[23]:
公式(1)和(2)中δ13Csc(‰)和δ15Nsn(‰)分别为添加外源有机物处理SOC的δ13C值和δ15N值;δ13Cs(‰)和 δ15Ns(‰)分别为无外源有机物添加处理SOC的δ13C值和δ15N值;δ13Cc(‰)和δ15Nc(‰)分别为初始外源有机物(秸秆和根茬)的δ13C值和δ15N值。
SOC中外源碳的含量(Mc,g·kg-1)和TN中外源氮的含量(Mn,g·kg-1)的计算公式如下[24]:
公式(3)和(4)中 Cmc(g·kg-1)和 Nmc(g·kg-1)分别为添加外源有机物处理SOC和TN含量。
土壤中外源碳残留率(Rc,%)和外源氮残留率(Rn,%)的计算公式如下:
公式(5)和(6)中Cm0(g)为初始玉米秸秆和根茬的碳含量;Nm0(g)为初始玉米秸秆和根茬的氮含量。
土壤中真菌残体碳(Fresidue-c,mg·kg-1)、细菌残体碳(Bresidue-c,mg·kg-1)、真菌残体氮(Fresidue-n,mg·kg-1)和细菌残体氮(Bresidue-n,mg·kg-1)的含量根茬据 LIANG等[25]所提供公式进行估算:
公式(7)、(8)、(9)和(10)中GluN(mg·kg-1)代表真菌衍生的氨基葡萄糖,MurA(mg·kg-1)代表细菌衍生的胞壁酸;179.2和251.2分别为氨基葡萄糖和胞壁酸的分子量;假定细菌细胞中氨基葡萄糖和胞壁酸的摩尔比为2∶1[26]。转化系数9和45分别用来将氨基葡萄糖转化为真菌残体碳含量和将胞壁酸转化为细菌残体碳含量;转化系数1.4和6.67分别将氨基葡萄糖转化为真菌残体氮含量和将胞壁酸转化为细菌残体氮含量[25,27-28]。
利用Microsoft Office Excel 2016和SPSS 25.0软件进行数据处理和统计分析。采用邓肯法(Duncan)和单因素方差分析方法(one-way ANOVA)进行差异显著性检验和多重比较。显著性水平均为0.05。采用Origin Pro 2017进行作图。图表中数据为平均值±标准误。
2 结果
2.1 土壤中外源碳氮的残留率
培养第 30天土壤外源碳残留率(Rc)是第 180天的1.36—1.90倍(图1-a)。培养第30天,添加秸秆处理土壤Rc较添加根茬处理增加了31.7%—44.1%;低肥土壤Rc平均比高肥土壤增加了14.8%。第180 天,添加秸秆和根茬处理Rc平均分别为36.3%和31.7%。培养第180天,土壤中外源氮残留率(Rn)受外源有机物类型和土壤肥力水平交互作用的影响显著(P<0.05)。添加秸秆和根茬处理Rn平均分别为95.8%和79.3%;秸秆碳和根茬碳在土壤中的平均残留率分别为 36.3%和 31.7%,秸秆氮和根茬氮则平均为 95.8%和79.3%。高肥土壤添加根茬处理Rn与低肥土壤相比增加了26.4%,然而高肥土壤添加秸秆处理Rn与低肥土壤相比降低了7.9%(图1-b)。

图1 不同肥力土壤中外源碳和氮的残留率Fig.1 Percentages of exogenous carbon and nitrogen remaining in black soil with different fertility levels
2.2 外源碳氮对土壤有机碳和全氮的贡献率
培养第30天,外源碳对土壤有机碳的贡献率(Fc)受外源有机物类型和土壤肥力水平影响显著(P<0.05),且添加秸秆处理Fc平均比添加根茬处理增加了26.4%(表2)。培养第180天,外源有机物类型、土壤肥力水平及它们的交互作用对 Fc影响差异不显著(P>0.05)。培养期间,外源有机物类型和土壤肥力水平显著影响(P<0.05)外源氮对土壤全氮的贡献率(Fn,表2)。培养第30天和第180天,添加秸秆处理 Fn平均分别比添加根茬处理增加了 42.4%和55.1%;低肥土壤添加外源有机物处理Fn平均分别比高肥土壤处理增加了19.4%和14.7%。
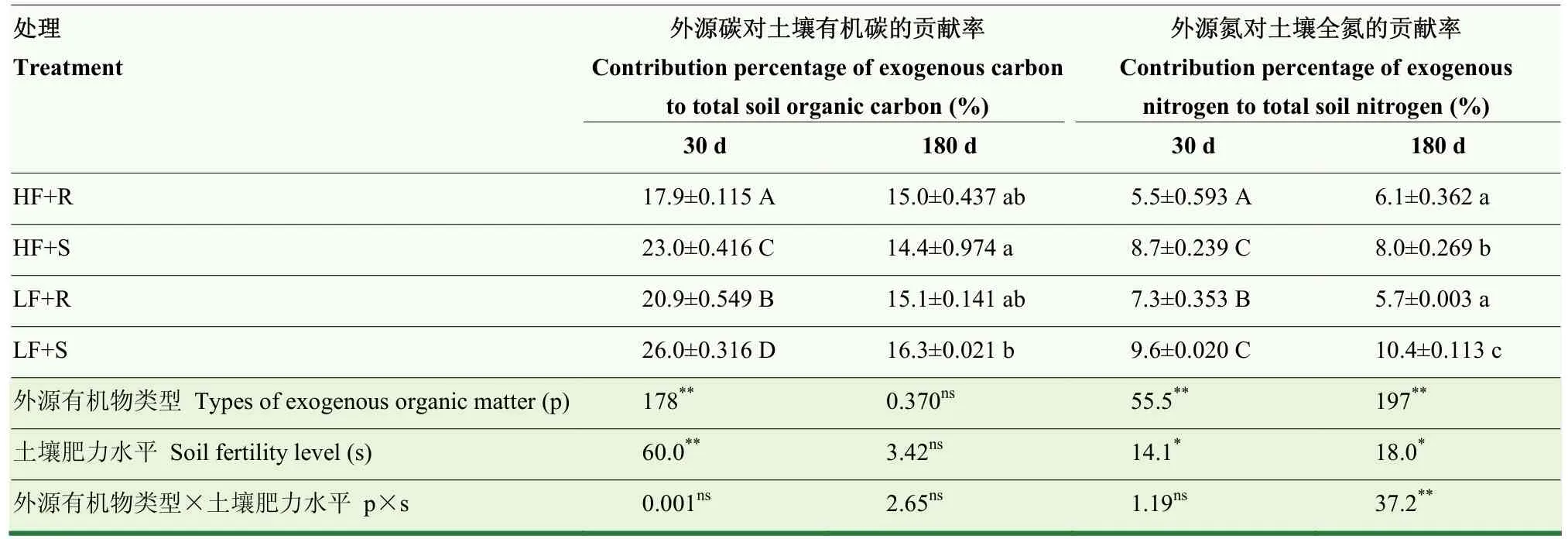
表2 不同肥力土壤外源碳对土壤有机碳和外源氮对土壤全氮的贡献率Table 2 Contribution percentages of exogenous carbon and nitrogen to total soil organic carbon and total soil nitrogen in different fertility levels of black soil, respectively
2.3 土壤有机碳与全氮的比值
培养第180天,外源有机物类型和土壤肥力水平显著影响(P<0.05)土壤有机碳与全氮的比值(SOC/TN)。土壤有机碳与全氮的比值在第30天为11.7—14.3,在第180天为 10.2—12.1(表 3)。培养第30天与第180天相比,添加根茬和秸秆处理SOC/TN平均分别下降了11.1%和14.9%;低肥土壤SOC/TN平均分别比高肥土壤增加了17.9%和11.6%。
土壤有机碳中外源碳与全氮中外源氮的比值(13C-SOC/15N-TN)受外源有机物类型的影响显著(P<0.05)(表3)。土壤中13C-SOC/15N-TN在第30天为 31.7—41.0,在第 180天为 16.7—32.0。第180天,添加根茬和秸秆处理13C-SOC与15N-TN的比值在平均分别为28.5和17.6,与第30天相比平均分别下降了28.2%和47.9%;高肥和低肥土壤13CSOC/15N-TN平均分别降低了38.0%和38.1%。

表3 不同肥力土壤总有机碳与全氮的比值及外源有机物来源碳氮的比值Table 3 Ratio of total organic carbon (SOC) to total nitrogen (TN) and that of SOC derived from exogenous carbon (13C-SOC) to TN derived from exogenous nitrogen (15N-TN) in different fertility levels of soils
2.4 土壤真菌、细菌残体碳氮的含量
培养期间,高肥土壤真菌和细菌残体碳含量分别是低肥土壤的1.17和1.31倍;添加秸秆处理微生物残体(真菌和细菌)碳含量平均比添加根茬处理增加了8.5%。培养第180天与第30天相比,高肥和低肥土壤真菌残体碳含量平均分别增加了 8.5%和 9.7%(图2-a)。第180天与初始土壤(表1)相比,高肥和低肥土壤细菌残体碳含量平均分别增加了 15.0%和31.9%(图2-b)。第180天,高肥和低肥土壤添加玉米秸秆处理与添加根茬处理相比,真菌残体碳含量分别增加了 7.3%和 6.1%,细菌残体碳含量分别增加了14.3%和16.6%,高肥和低肥土壤中真菌残体碳含量平均分别为细菌残体碳的 3.30和3.69倍。培养第 180天,添加玉米秸秆处理较添加根茬处理相比真菌和细菌残体碳含量分别平均增加了6.7%和15.6%;添加根茬和秸秆处理真菌残体碳含量平均分别为细菌残体碳含量的3.65和3.26倍。
真菌残体氮和细菌残体氮受外源有机物类型和土壤肥力水平的影响与真菌残体碳和细菌残体碳基本一致(图 2)。整个培养期间,高肥和低肥土壤中真菌残体氮含量平均分别为细菌残体氮含量的3.46和3.87倍;添加根茬和秸秆处理真菌残体氮含量平均分别为细菌残体氮含量的3.65和3.26倍(图2-c和2-d)。第180天与初始土壤(表1)相比,高肥和低肥土壤细菌残体氮含量平均分别增加了 15.0%和 31.9%(图2-d)。

图2 不同肥力土壤中真菌、细菌残体碳氮含量Fig.2 Contents of fungal residue-carbon and -nitrogen and those of bacterial residue-carbon and -nitrogen in different fertility levels of black soils
2.5 土壤微生物残体碳氮对土壤有机碳和全氮的贡献率
土壤肥力水平显著影响(P<0.05)微生物残体碳(细菌残体碳和真菌残体碳)对SOC的贡献率(图3)。培养第180天,真菌残体碳对SOC的贡献率为 33.0%—37.4%(图 3-a),细菌残体碳对 SOC的贡献率为8.5%—11.7%(图 3-b)。真菌残体碳对高肥和低肥土壤 SOC的贡献率平均分别为 37.0%和33.8%,细菌残体碳的贡献率平均分别为 11.2%和9.2%;添加秸秆和根茬的处理真菌残体碳对SOC的贡献率平均分别为 36.0%和 34.7%,细菌残体碳的贡献率平均分别为10.8%和9.6%。第180天与第30天相比,低肥土壤真菌和细菌残体碳对SOC的贡献率平均分别增加了33.0%和19.1%。与初始土壤(表1)相比,第180天时高肥和低肥土壤细菌残体碳对SOC的贡献率增加了 4.8%和 20.3%。高肥土壤添加根茬和秸秆处理微生物残体碳对SOC的贡献率与低肥土壤相比,第30天平均分别增加了24.3%和17.0%;第180天平均分别增加了13.5%和10.5%(图3)。

图3 不同肥力土壤中微生物残体碳对土壤有机碳的贡献率Fig.3 Contribution percentage of microbial residue carbon to total soil organic carbon (SOC) in different fertility levels of black soils
培养第30天,真菌残体氮和细菌残体氮对TN的贡献率平均分别为55.2%和16.3%(图 4)。第180天,低肥和高肥土壤真菌残体氮对TN的贡献率平均分别为 63.5%和 60.5%,细菌残体氮的贡献率平均分别为 16.4%和 17.5%;高肥和低肥土壤添加秸秆处理细菌残体氮对TN的贡献率较添加根茬处理分别提高了3.2%和14.6%(图 4-b)。培养180天与初始土壤相比,细菌残体氮对高肥和低肥土壤TN的贡献率平均分别增加了7.4%和32.5%。

图4 不同肥力土壤中微生物残体氮对土壤全氮的贡献率Fig.4 Contribution percentage of microbial residue nitrogen to total nitrogen (TN) in different fertility levels of black soils
3 讨论
3.1 土壤肥力水平对土壤微生物残体碳氮累积的影响
无论是秸秆还是根茬还田后第180天,外源碳和氮的残留率受土壤肥力的影响不明显(P>0.05)(图1),这与李丽东等[21]研究结果不一致。这可能与本研究黑土本身有机质含量较高有关[18],再加上恒温恒湿的室内培养环境(25 ℃和 60%田间持水量)使土壤微生物活性一直处于较高状态,促进了外源有机物的分解,并使其进入了稳定分解的阶段[29]。
微生物残体碳是 SOC库的重要组成部分[25],在SOC的积累和稳定过程中起着至关重要的作用[30-31]。农田土壤氮磷钾肥配施显著增加了土壤氨基糖单糖和总氨基糖的含量,促进了微生物残体碳在土壤中的固存[32]。有研究表明[33]有机碳含量低的土壤中微生物尤其是细菌群落对活性碳的响应更强烈。与初始土壤(表1)相比,培养第180天低肥和高肥土壤细菌残体碳氮含量平均分别增加了 31.9%和 15.0%,这说明肥力水平较低的土壤由于长期受养分限制,细菌对可利用底物的响应更为强烈[33]。细菌能够快速分解底物以合成自身生物量,进而促进细菌残体碳氮在土壤中的累积,这与李丽东等研究结果一致[21]。本研究中土壤肥力水平显著影响(P<0.05)真菌和细菌残体碳对SOC的贡献率和真菌残体氮对TN的贡献率,这与以往研究结果一致[3,34]。高肥土壤添加外源有机物真菌和细菌残体碳对SOC的贡献率大于低肥土壤,这可能是因为高肥土壤本身具有较高的有机碳和全氮含量及微生物量,从而促进了微生物残体碳累积。培养第180天与初始土壤(表 1)相比,高肥土壤细菌残体碳和氮对SOC和TN的贡献率分别增加了4.8%和7.4%,低肥土壤则分别增加了20.3%和32.5%(图3),这说明外源有机物添加较有利于低肥土壤细菌残体碳氮的积累及其对SOC和TN的贡献。高肥土壤微生物的生物量和活性较高,为重新代谢微生物残体创造了合适的环境,使微生物残体碳氮在土壤中积累缓慢[35]。对于低肥土壤来说,外源底物添加激发了土壤微生物活性,且在饥饿条件下细菌群落对外源底物利用反应比真菌迅速[33];另外,细菌自身的碳氮比较低,对氮的需求量更高,同化碳的同时也利用了大量的氮,而且细菌残体氮作为有效氮源,可能再次被活体微生物分解利用[36],因此促进低肥细菌残体碳和氮对SOC和TN贡献率的提高[37]。
3.2 外源有机物类型对土壤微生物残体碳氮累积的影响
本研究中,随着微生物对秸秆和根茬的分解,外源碳的残留率逐渐减少(Rc)(图1-a),一方面可能是由于玉米秸秆和根茬的分解导致SOC含量降低,另一方面可能是培养试验的前30 d是玉米秸秆和根茬的快速分解时期。在养分条件较好的黑土中,微生物活性最大,种类较多[29],外源有机物中可溶性有机物被微生物快速分解,因此秸秆分解速度较快,残留率降低;30 d后秸秆分解速度减慢,说明微生物开始分解秸秆中蛋白质、磷脂、木质素等难分解物质。秸秆和根茬化学组分的差异可能是影响秸秆碳残留率变化的主要原因[38]。
外源底物添加的数量和类型是影响微生物过程的重要因素[39]。研究表明施入不同种类有机物料对农田黑土微生物残体积累数量影响显著,同时真菌和细菌对有机物料添加的响应不同[34,40]。本研究发现外源有机物类型显著影响微生物残体碳含量(P<0.05)。培养第180天,添加玉米秸秆处理与添加根茬处理相比,真菌和细菌残体碳含量分别平均分别增加了 6.7%和15.6%。秸秆与根茬相比具有较低的C/N和较高比例的易分解组分[6-7],易于被微生物分解,从而促进了微生物残体碳尤其是细菌残体碳在土壤中的积累[5],增加了微生物残体碳对SOC的贡献率。同时微生物残体的“续埋效应”可能是影响SOC稳定积累的主要因素[41]。
作物秸秆和根茬是农田土壤氮素的重要来源,微生物通过同化外源氮,合成微生物残体氮,然后固存在土壤氮库中。有研究表明[42-43],外源氮素的添加提高了土壤微生物生物量碳和微生物残体的积累,微生物对外源底物难降解组分的利用程度显著影响外源氮向微生物氮的转化。与初始土壤(表 1)相比,培养第 30天时添加玉米秸秆和根茬处理细菌残体氮含量分别增加了 22.5%和 23.5%,真菌残体氮含量分别增加了1.6%和3.0%(图2),说明培养初期细菌群落较易利用外源有机物进行自身体内氮的合成。随着外源有机物的分解,易分解有机物的逐渐减少,真菌具有较强底物同化能力[39],容易利用难分解的有机物,因此培养第180天与第30天相比,真菌和细菌残体氮含量分别增加了12.4%和3.6%。培养第180天与第30天相比,添加玉米秸秆和根茬处理微生物残体氮含量平均分别增加了12.2%和2.2%(图2-c和2-d)。玉米秸秆含氮量高于根茬,可为微生物生长持续供应所需养分,促进了微生物残体氮的累积。
4 结论
土壤肥力水平和外源有机物类型显著影响不同微生物群落残体碳氮在土壤中的累积。微生物残体氮对土壤全氮的贡献率达到一半以上,说明微生物残体氮对土壤氮库的扩容起重要作用。低肥土壤添加秸秆和根茬提高了细菌残体碳氮的累积及其对土壤有机碳和土壤全氮的贡献,从而能有利于细菌残体碳和氮向土壤有机碳库和氮库的转化。玉米秸秆较玉米根茬更能促进微生物残体碳氮在土壤中的累积,更有利于土壤有机碳库的稳定和氮库的扩容。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
土壤学报(2021年4期)2021-09-10
军事运筹与系统工程(2020年2期)2020-11-16
西藏农业科技(2019年3期)2019-11-04
生态学报(2019年8期)2019-05-31
军事运筹与系统工程(2018年3期)2018-03-26
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
中亚信息(2016年10期)2016-02-13
西藏大学学报(自然科学版)(2015年1期)2015-12-25
