
高温胁迫对不同耐热型马铃薯块茎形成期生长和光合特性的影响*
2023-05-21 02:56周进华李有涵张兴胡荣海郭华春
中国生态农业学报(中英文) 2023年5期
周进华 ,李有涵 ,张兴 ,胡荣海 ,郭华春**
(1. 云南农业大学农学与生物技术学院薯类研究所 昆明 650201;2. 云南泵龙马铃薯种植有限公司 昆明 655200)
全球变暖日益加剧,在过去的50 年中,气温升高的速度是20 世纪的2 倍,到21 世纪末,全球平均气温预计将上升1.5~4.5 ℃,部分地区已出现极端高温气候[1-2]。目前,马铃薯(Solanum tuberosumL.)已成为谷类作物外,最重要的粮食作物。其喜性冷凉,对高温敏感。高温对马铃薯植株形态、光合作用、块茎形成和产量品质等均有显著影响。与茎叶相比,马铃薯的块茎发育过程对高温更为敏感,当环境温度超过25 ℃时,块茎则停止生长,温度超过39 ℃时,茎叶停止生长[3]。Kooman等[4]发现,高温会限制马铃薯叶片的扩展,导致叶面积缩小,同时影响茎的伸长。
光合作用是作物生长、产量和品质形成的前提,也是响应高温胁迫最为敏感的生理过程之一,因此光合作用相关生理参数可作为评价作物高温抗性的重要指标[5-6]。高温胁迫对植物的光反应和暗反应均有明显影响,不同植物或同种植物的不同耐热型种质资源在受到高温胁迫时,叶片光合特征参数的变化规律也不一致[7]。马铃薯光合作用的最低温度为0~7 ℃,最适温度为16~25 ℃,最高温度为40 ℃[8]。马铃薯净光合速率与暗呼吸速率比例最适的环境温度为16~20 ℃,当温度每升高10 ℃,暗呼吸速率增加1倍[9]。欧洲大部分马铃薯栽培种光合作用的最适温度为20 ℃,当温度降至10 ℃时,净光合速率降低25%;当温度在20 ℃的基础上每增加5 ℃时,净光合速率降低大约25%,当温度高于30 ℃时,马铃薯植株的净光合速率降低至0[10]。高温会抑制马铃薯的光合作用,与耐热型马铃薯相比,热敏感型马铃薯在40 ℃下处理9 d 后,叶片的CO2固定受到更严重的抑制,并且光合色素损失更为明显[11]。Wang等[12]以‘陇薯3 号’为研究对象,分别在马铃薯苗期和分枝期进行6 h 的高温处理,发现高温胁迫对分枝期马铃薯光合作用的影响大于苗期。
目前,高温胁迫对马铃薯几个关键生长时期叶片光合作用的影响已有报道,但针对块茎发育时期的相关研究鲜有报道。块茎形成期作为马铃薯经济产量形成最基础的时期,在生产中具有决定性作用,同时耐热型和热敏感型马铃薯种质资源在高温胁迫下的光合特性差异还不明确。本研究在马铃薯块茎形成期,比较分析高温胁迫下热敏感型品种‘青薯9 号’和耐热型品系‘滇187’的植株形态、结薯性状和叶片光合特性的差异,从形态和光合特性方面综合解析马铃薯耐热性存在差异的生理机理,为马铃薯高温抗性的机制研究提供参考依据,为耐热品种选育奠定理论基础。
1 材料与方法
1.1 试验材料
供试马铃薯材料由云南农业大学薯类研究所提供,分别为热敏感型品种‘青薯9 号’和耐热型品系 ‘滇187’[13]。
1.2 植物材料培养环境
试验材料采用盆栽种植于人工气候箱(赛福实验仪器公司)内,培养箱长、宽、高分别为160 cm、60 cm、120 cm,采用LED 植物生长灯垂直光照,控光范围:0~200 μmol·m-2·s-1,控温范围:10~50 ℃,控湿范围:40%~95%。营养钵高15 cm,直径12 cm,容积为1 L。单个培养箱可放置65 盆。
1.3 植物材料高温胁迫处理方法
将生长一致的马铃薯组培苗顶芽转接于MS 培养基中培养20 d [16 h 光照/8 h 黑暗、光强度100 μmol·m-2·s-1、温度(20±1) ℃],随后将组培苗移栽至经高温灭菌的基质土中,置于人工气候箱中炼苗20 d[16 h 光照/8 h 黑暗、光强度为100 μmol·m-2·s-1、温度(20±0.5) ℃、相对湿度为75%]。选择生长一致的植株进行试验处理,将人工气候箱培养条件调整为12 h 光照/12 h 黑暗,光强200 μmol·m-2·s-1,相对湿度45%,温度(20±0.5) ℃,在此条件下培养至植株匍匐茎形成期。再选取20 株生长势一致的植株进行高温处理[(30±0.5) ℃],20 株继续在(20±0.5) ℃下培养。2 个处理除温度不同外,其余环境因子均相同。试验处理期间每天浇水,保证土壤相对含水量在田间持水量的80%。
1.4 植株形态及光合作用参数测定方法
1.4.1 植株形态指标测定
植株高温处理2 周后(块茎形成期),每个处理选取10 株进行形态指标测量。株高采用直尺测量植株基部到顶端生长点的长度(cm);茎粗采用游标卡尺测量植株倒4 叶与倒5 叶间主茎的直径(mm);叶面积采用长宽系数法,用直尺测定植株倒4 叶顶小叶的最长和最宽,并采用公式计算:叶长×叶宽×0.76(cm2);叶宽长比=叶宽/叶长;叶全长采用直尺测定植株倒4 叶的叶柄基部到叶尖的长度(cm);叶夹角用量角器测定植株倒4 叶的叶柄与主茎之间的角度(°);叶垂角用量角器测定植株倒4 叶的叶尖到叶柄基部的直线与主茎之间的角度(°);披垂角=叶垂角-叶夹角(°);节间长用直尺测定每两个节之间的间距,并计算平均值(cm);小叶数统计植株倒4 叶顶小叶和侧小叶数量;叶片数为马铃薯植株绿色复叶的数量。
1.4.2 光合作用参数测定
在开展植株形态指标测定的同时,使用Li-6400XT 光合-荧光记录仪(Li-Cor,USA)对叶片气体交换参数和叶绿素荧光参数进行测定,所有参数均选择植株倒4 叶的顶小叶进行测定。净光合速率、光响应曲线、CO2响应曲线、光诱导曲线和叶绿素荧光参数的测定均参照Li-6400XT 光合-荧光记录仪的操作说明,测定完成后,参照双升普等[14]的方法进行相关光合特征参数计算。
1.4.3 产量测定
当对照组处于块茎成熟期时,收获各处理组植株薯块,用分析天平称量马铃薯的单株产量(g)、统计单株结薯数,计算平均单个薯重=单株产量/单株结薯数(g),每个处理组选取9 个植株进行数据统计。
1.4.4 可塑性分析
在高温和适温处理测定指标的所有生物学重复中找出最大值和最小值,并用公式计算:可塑性指数=(最大值-最小值)/最大值。
1.5 数据处理
使用Excel 2019 进行数据处理,SPSS 26.0 进行数据统计分析,用Origin 2020b 进行主成分分析,用Sigmaplot 14.0 和GraphPad Prism 8 拟合曲线及作图。
2 结果与分析
2.1 ‘青薯9 号’和‘滇薯187’在高温胁迫下的结薯特征
‘青薯9 号’(QS9)在20 ℃正常结薯,单株结薯重量为13.05 g、单株结薯数目为1.89 个、单个薯平均重量为8.65 g;而QS9 在30 ℃仅形成少量匍匐茎,不能结薯(图1,图2)。‘滇187’(D187)在20 ℃下,单株结薯重量为22.89 g、单株结薯数目为2.89 个、单个薯平均重量8.6 g;与QS9 不同,D187 在30 ℃下,依然可以结薯,但结薯性状各项指标均显著(P<0.01)下降(图2)。根据上述结果,将QS9 归为热敏感型种质,而将D187 归为耐热型种质,本研究将利用这2个品种(系)开展后续研究。

图1 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的结薯表现Fig.1 Tuberization of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
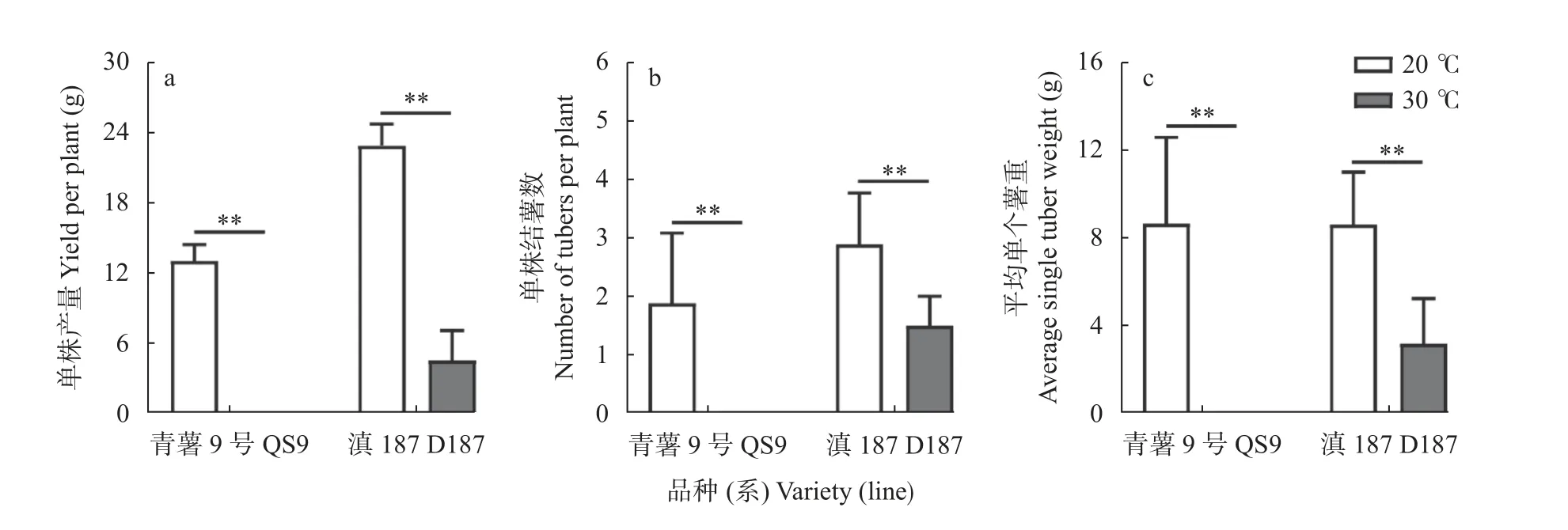
图2 ‘青薯9 号’和‘滇187’马铃薯在正常温度(20 ℃)和高温(30 ℃)下的产量性状Fig.2 Yield characters of potato materials QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
2.2 耐热型和热敏感型马铃薯在高温胁迫下的植株形态变化
与20 ℃相比,在30 ℃下生长的QS9 和D187植株株高显著(P<0.01)增加,增长比例分别为49.05%和49.33% (图3,图4a),但节数没有显著变化(图4b),但节间显著伸长(图4c)。与20 ℃相比,在30 ℃下QS9 和D187 的茎粗没有显著改变(图4d),QS9 的叶片数目显著(P<0.05)减少,而D187 没有显著改变(图4e)。马铃薯的叶面积在高温下显著(P<0.01)缩小,QS9 和D187 的叶面积分别显著减小48.59%和54.36%,叶全长分别显著减小8.39%和11.41%,而叶片宽长比和小叶数并无显著变化(图4i-l)。高温下,马铃薯植株叶片更为直立,展现出向上性生长(图3),QS9 和D187 叶片夹角分别显著(P<0.01)减少39.29%和45.72% (图4f),叶垂角分别显著(P<0.01)减少43.67%和31.47% (图4g),QS9 的披垂角显著(P<0.01)减少54.45%,但D187 无显著变化(图4h)。由以上分析可知,高温胁迫对不同耐热型马铃薯的植株形态影响是相似的,但在高温环境下耐热型的D187 的叶片数目和披垂角性状更为稳定。

图3 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下植株株型和叶片性状Fig.3 Plant architecture and leaf morphology of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures

图4 ‘青薯9 号’和‘滇187’马铃薯在正常温度(20 ℃)和高温(30 ℃)下植株株型和叶片形态指标Fig.4 Morphological indexes of plant architecture and leaf of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃)temperatures
2.3 耐热型和热敏感型马铃薯在高温下的光合特征
在高温胁迫下,D187 的净光合速率(Pn)和气孔导度(Gs)均呈上升趋势(未达显著水平);而QS9的Pn和Gs均呈下降趋势,其中Pn显著下降26.90%(P<0.01)。QS9 和D187 的胞间CO2浓度(Ci)、水分利用效率(WUE)均表现出下降趋势,且WUE 下降水平显著(P<0.01),蒸腾速率(Tr)均表现为显著上升(P<0.05)。与QS9 相比,高温胁迫下D187 有较高的Pn和WUE,说明高温胁迫下耐热型马铃薯的光合能力更强(表1)。

表1 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的净光合速率及相关参数Table 1 Net photosynthetic rate and related parameters of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃)temperatures
2.4 耐热型和热敏感型马铃薯在高温下净光合速率响应光照强度、CO2 浓度及相关参数的变化特征
2.4.1 光响应特征及相关参数
光响应相关参数能有效反映植物对光强度的利用范围,高温处理下2 个不同耐热型马铃薯的最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd)均升高,其中D187的Pnmax显著升高60.18% (P<0.01),QS9 的LSP、LCP 分别显著升高41.37% (P<0.01)、40.54% (P<0.05)。D187 的表观量子效率(AQY)在高温处理下无显著变化,而QS9 的AQY 则显著降低25.00% (P<0.01) (表2)。由此说明,高温下马铃薯对强光的利用能力增强,对弱光的利用能力减弱,呼吸消耗增加。高温胁迫下,D187 比QS9 具有更高的Pnmax和AQY,更低的LCP和Rd,这说明耐热型马铃薯在高温胁迫下具有更强的光能利用能力和更低的呼吸消耗。
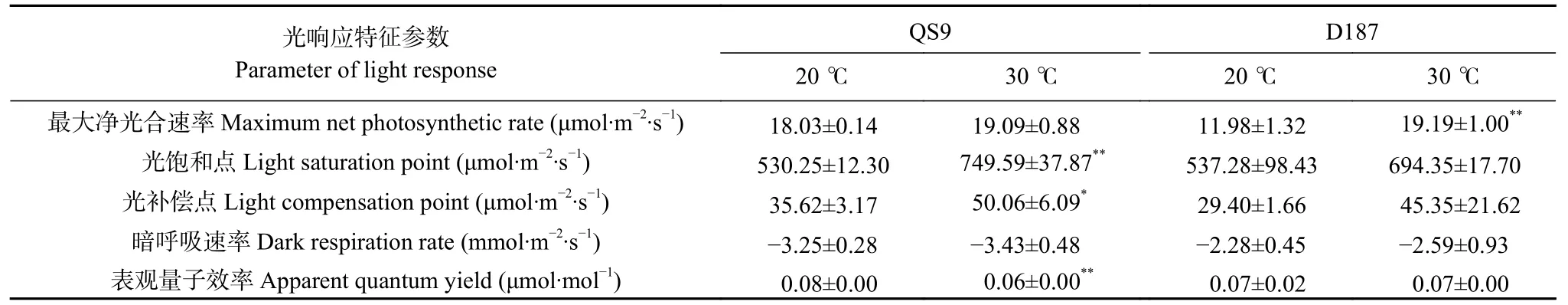
表2 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的光响应特征参数Table 2 Parameters of light response of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
2.4.2 CO2 响应特征及相关参数的影响
与20 ℃相比,在30 ℃下QS9 和D187 的光呼吸速率(RL)降低,CO2补偿点(CCP)升高,最大电子传递速率(Jmax)及Jmax/Vcmax均降低,其中QS9的RL显著降低26.21% (P<0.05),CCP 显著升高24.77%(P<0.01),Jmax/Vcmax显著降低了11.22% (P<0.05),但羧化效率(CE)无变化(表3)。此外,高温处理对QS9和D187的Vcmax影响均不显著。由此可知,伴随着温度的升高,马铃薯对高浓度CO2的利用能力增强,对低浓度CO2的利用能力减弱,光下的呼吸消耗增强。D187 在高温下比QS9 具有更高的CE、Vcmax、Jmax和Jmax/Vcmax,说明在面对高温胁迫时耐热型马铃薯具有更高的碳同化能力和RuBP 的再生潜力。
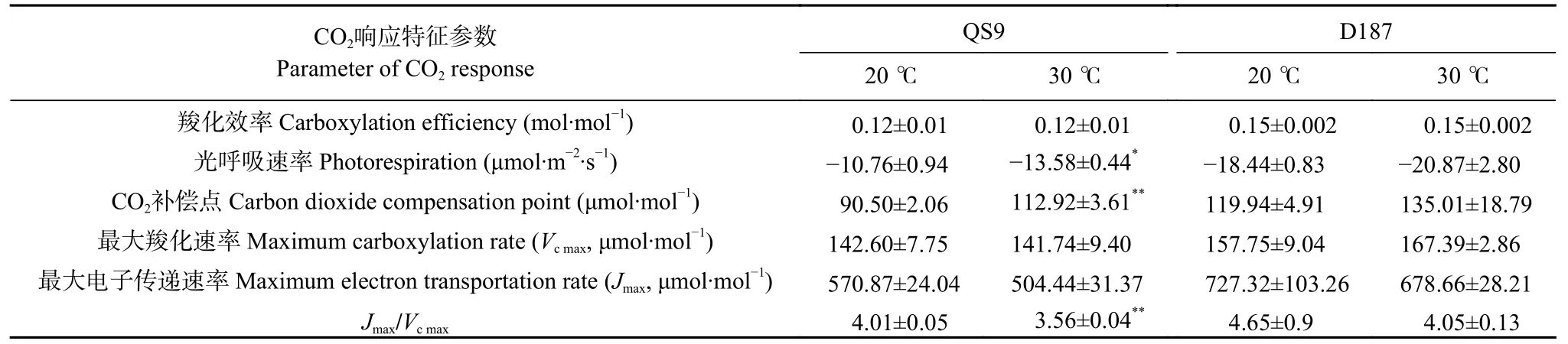
表3 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的CO2 响应特征参数Table 3 Parameters of CO2 response of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
2.5 高温胁迫对不同耐热型马铃薯光合诱导特征及相关参数的影响
由表4 可知,高温对QS9 和D187 光合诱导相关参数的影响一致。30 ℃处理下,2 个不同耐热型马铃薯暗适应后达到最大净光合速率30%所需的时间(T30%P)、暗适应后达到最大净光合速率60%所需的时间(T60%P)、暗适应后达到最大净光合速率90%所需的时间(T90%P)显著减少32.92%~52.24% (QS9P<0.01;D187P<0.05),暗适应后60 s 所达到的最大净光合速率百分比(IS60s) (P<0.01)、暗适应后300 s所达到的最大净光合速率百分比(IS300s) (QS9P<0.01;D187P<0.05)、暗适应后600 s 所达到的最大净光合速率百分比(IS600s)显著(QS9P<0.01;D187P<0.05)升高28.69%~234.86%。说明马铃薯的Pn在高温胁迫下能更快达到最大值。

表4 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的光合诱导特征参数Table 4 Parameters of photosynthetic induction of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃)temperatures
2.6 不同耐热型马铃薯在高温下叶绿素荧光参数及其在光响应和光诱导过程中的变化
2.6.1 对黑暗下叶绿素荧光参数的影响
对叶片进行叶绿素荧光检测可评价高温胁迫对叶绿体的损伤程度。对表5 分析发现,高温胁迫下,2 个不同耐热型马铃薯的叶绿素荧光参数均降低,其中黑暗下最大荧光强度(Fm)、光系统Ⅱ(PSⅡ)潜在光化学量子效率(Fv/Fm)和PSⅡ潜在光化学活性(Fv/F0)显著降低(P<0.05,P<0.01)。当Fv/Fm<0.75 时,表明植物受到不可逆的严重胁迫损伤[15]。在30 ℃高温胁迫下,QS9 和D187的Fv/Fm均高于0.75,说明30 ℃的高温胁迫没有造成马铃薯叶片中叶绿体的不可逆损伤。

表5 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下的叶绿素荧光参数Table 5 Chlorophyll fluorescence parameters of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
2.6.2 对光下叶绿素荧光参数的影响
由图5 可知,不同温度处理下,2 个耐热型马铃薯的荧光特征参数在光响应过程中的变化一致,并且在大于300 μmol·m-2·s-1的光照强度下,高温处理与对照才表现出明显差异。随着光照强度增强,电子传递速率(ETR)先快速增加后达到稳定(图5a);最大光化学量子效率(Fv′/Fm')、实际光化学量子效率(ΔF/Fm')和光化学猝灭系数(qP)均降低(图5c,e,g);非光化学猝灭系数(NPQ)升高(图5i)。高温处理使QS9 和D187 的ETR、Fv'/Fm'、ΔF/Fm'和qP升高,但两者间并无显著差异。在1000 μmol·m-2·s-1的光照强度下,不同温度处理下Fv'/Fm'趋于一致,但ΔF/Fm'在300 μmol·m-2·s-1的光照强度后始终保持着显著(P<0.05)差异。高温下,两个马铃薯品种的NPQ 均降低,但QS9 下降趋势更明显。在1000 μmol·m-2·s-1的光照强度后,2 个品种的NPQ 又趋于一致。
光诱导过程中,不同温度处理的QS9 和D187 的荧光特征参数变化一致。随着诱导时间延长,ETR、ΔF/Fm'、qP和NPQ 快速升高后逐渐趋于平稳(图5b,f,h,j),Fv'/Fm'迅速下降后逐渐趋于平稳(图5d)。高温处理使2 个不同耐热型马铃薯的ETR、ΔF/Fm'、qP和NPQ 升高,但处理间和材料间的差异均不显著。NPQ 在光照下诱导180 s 后又缓慢降低,且在高温处理下的QS9 降低到与对照相同。Fv'/Fm'在高温处理下降低,但2 种基因型马铃薯的变化差异均不显著。
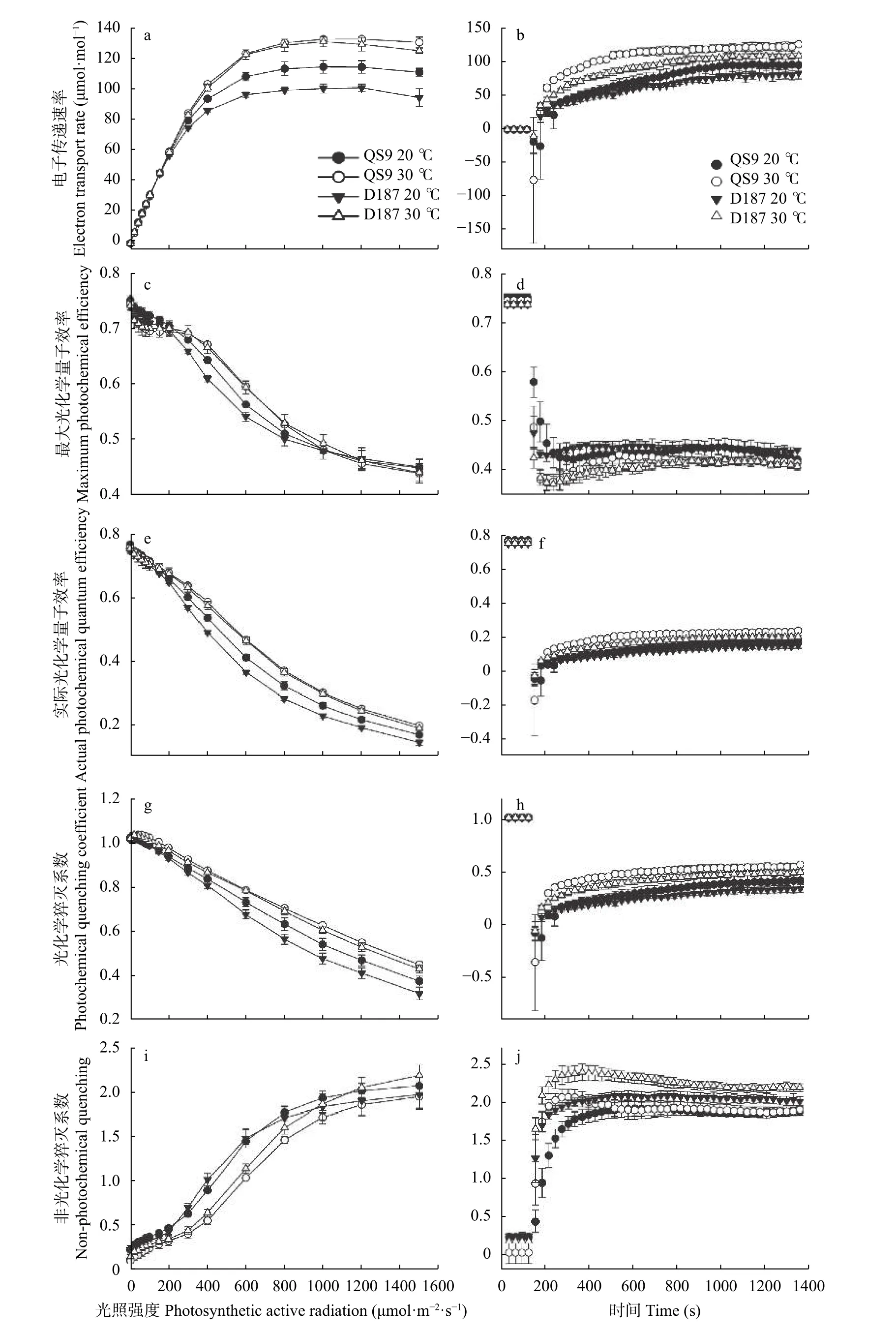
图5 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下光响应和光诱导过程中荧光特征参数的变化Fig.5 Changes of fluorescence characteristic parameters of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃)temperatures during light response and photosynthetic induction
2.7 耐热型和热敏感型马铃薯光能分配在光响应和光诱导过程中的变化
马铃薯叶片捕获的光能分配到不同耗散途径的比例在光响应和光诱导过程中表现出不同变化趋势(图6)。随着光照增强,QS9 和D187 在不同温度下的光化学耗散比例(ΦPSⅡ)逐渐减小,而非光化学猝灭耗散比例(ΦNPQ)和荧光耗散比例(Φf,d)逐渐增加。高温处理下,2 个不同耐热型马铃薯的ΦNPQ较对照显著减少(P<0.05),ΦPSⅡ显著增加(P<0.05),Φf,d无显著变化,且试验材料间光能分配到不同耗散途径的比例无显著差异。随着光诱导时间的延长,2 个不同耐热型马铃薯的ΦNPQ和Φf,d快速升高后缓慢降低,而ΦPSⅡ迅速降低后缓慢增加。高温处理下,QS9 和D187的ΦNPQ和Φf,d较对照均显著减少(P<0.05),ΦPSⅡ显著增加(P<0.05),且QS9的ΦPSⅡ比D187 显著增加(P<0.05),ΦPSⅡ显著减少(P<0.05),Φf,d无显著变化。由此表明,高温改变了马铃薯的光能分配,且热敏感型马铃薯比耐热型的分配比例变化更大。
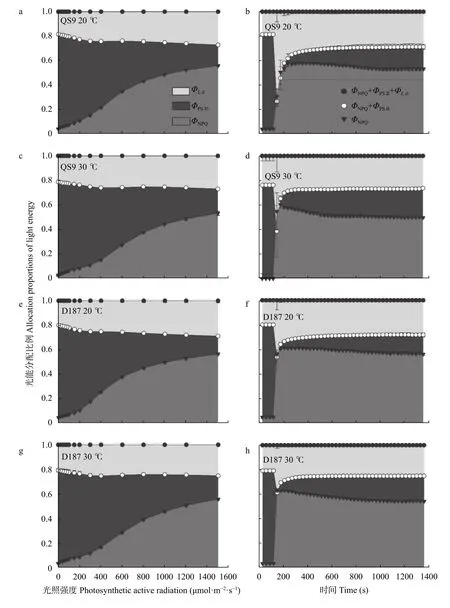
图6 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯在正常温度(20 ℃)和高温(30 ℃)下光响应和光诱导过程中的光能分配特征Fig.6 Characteristics of light energy distribution of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures during light response and photosynthetic induction
2.8 高温下不同耐热型马铃薯植株形态、光合特征和产量性状的可塑性
由图7 可知,在形态指标中,可塑性指数大于0.5 的参数,QS9 有4 个,占比为33.33%;D187 有6 个,占比为50%。在光合特征中,QS9 可塑性指数大于0.5 的指标有6 个,占比为23.08%;而D187 有10 个,占比为38.46%。总体上,QS9 的平均可塑性指数为0.418,D187 的平均可塑性指数为0.448,在高温胁迫下D187 在形态和光合作用方面可塑性高于QS9。这说明D187 能够更好地通过调节植株形态和光合作用来适应高温环境。
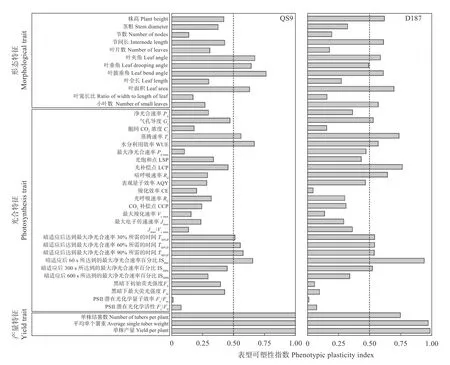
图7 ‘青薯9 号’(QS9)和‘滇187’(D187)马铃薯高温(30 ℃)下形态、光合和产量特征等各参数的表型可塑性指数Fig.7 Phenotypic plasticity indexes of parameters of morphology,photosynthesis,and yield of potato materials of QS9 and D187 at high (30 ℃) temperatures
2.9 耐热型和热敏感型马铃薯响应高温的主成分分析
对QS9 和D187 的41 个形态特征、光合特征和产量特征参数进行主成分分析,得到各主成分对马铃薯响应高温的特征值和贡献率(表6)。从表中可以看出,QS9 在PC1 和PC2 的累计贡献度为83.58%,D187 为80.90%,且特征值都大于1,因此这两个主成分都能有效反映参数特征,并能够充分解释马铃薯对高温的响应特征。QS9 和D187 的点在PC1/PC2平面上完全聚集为2 簇,同时PC1 明显将高温处理和对照分开,表明本研究所设置的高温胁迫对2 个马铃薯品种(系)的生长和抗性生理指标均有影响(图8)。
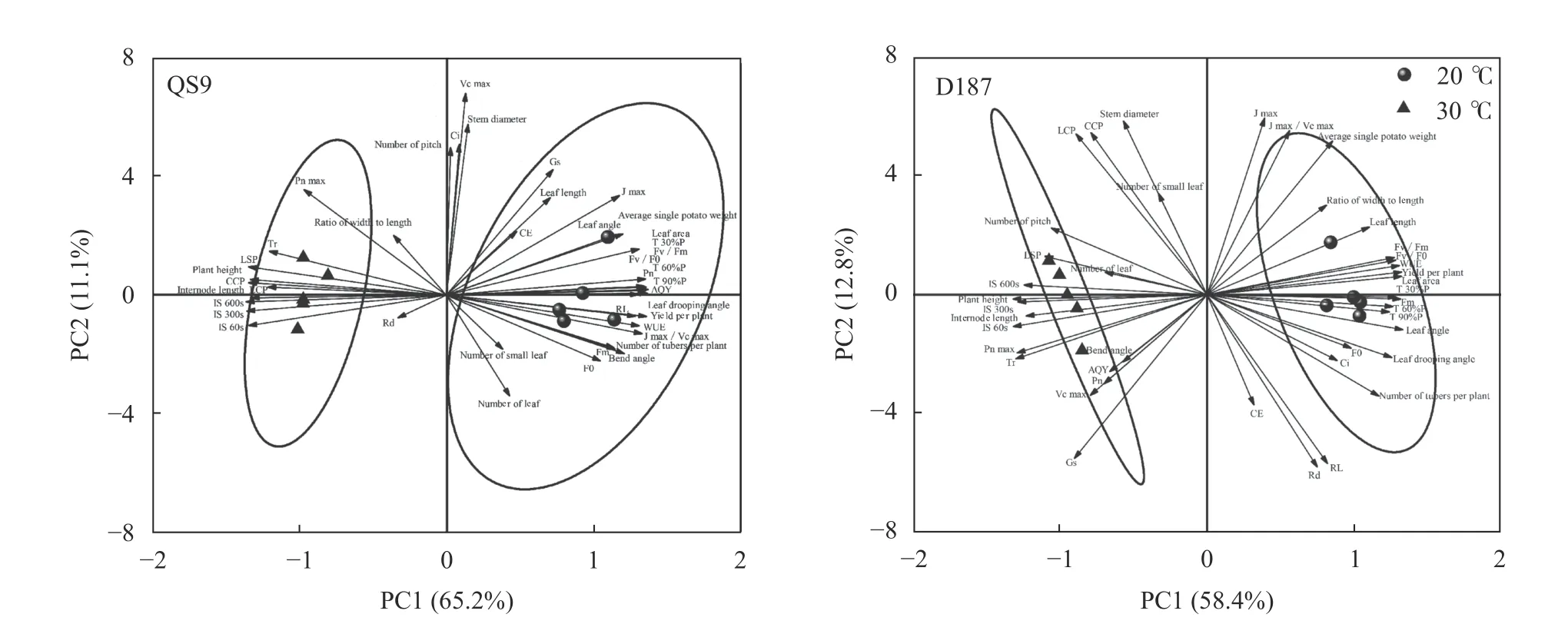
图8 ‘青薯9 号’(QS9)和‘滇187’(D187)在正常温度和高温下的形态、光合及产量指标的主成分分析Fig.8 Principal component analysis of morphology,photosynthesis,and yield indexes of potato materials of QS9 and D187 at normal (20 ℃) and high (30 ℃) temperatures
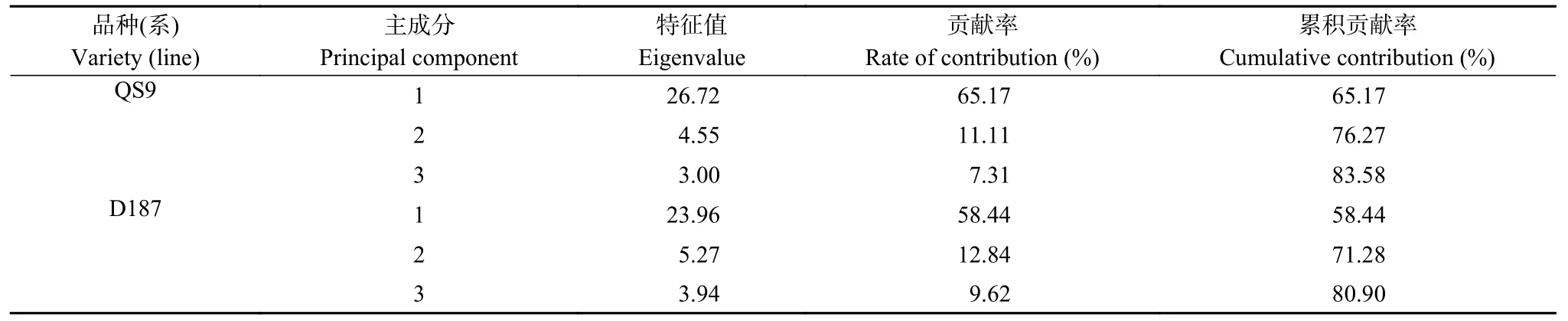
表6 ‘青薯9 号’(QS9)和‘滇187’(D187)形态特征、光合特征和产量特征参数初始特征值及累积贡献率Table 6 Initial eigenvalues and accumulated variance contribution of each component of morphology,photosynthesis,and yield of potato materials of QS9 and D187
QS9 的6 个形态指标参数(株高、节间长、叶面积、叶夹角、叶垂角和披垂角)、20 个光合特征参数(Pn、Tr、LSP、CCP、LCP、WUE、AQY、RL、Jmax/Vcmax、IS60s、IS300s、IS600s、T30%P、T60%P、T90%P、F0、Fm、Fv/Fm和Fv/F0)和3 个产量特征参数(单株产量、单株结薯数和平均单个薯重)在PC1 中的权重系数和载荷系数较大,对PC1 有较大贡献,其中株高、节间长、Tr、LSP、CCP、LCP、IS60s、IS300s和IS600s与PC1 呈负相关关系,其他特征参数呈正相关。D187的7 个形态特征参数(株高、节间长、节数、叶全长、叶面积、叶夹角和叶垂角)、13 个光合特征参数(Pnmax、Tr、LSP、WUE、IS60s、IS300s、IS600s、T30%P、T60%P、T90%P、Fm、Fv/Fm和Fv/F0)和2 个产量特征参数(单株产量和单株结薯数)在PC1 中的权重系数和载荷系数较大,对PC1 有较大贡献,其中株高、节间长、节数、Tr、LSP、Pnmax、IS60s、IS300s和IS600s与PC1 呈负相关关系,其他特征各参数呈正相关。
总体而言,高温胁迫下QS9 光合作用和产量性状相关参数对PC1 的贡献率高于D187,这说明,由高温引起的光合作用参数变化在D187 和QS9 两个品种(系)之间存在明显差异,更多的光合作用相关参数被用于解释QS9 在高温与适宜生长环境下的区别,高温对耐热型马铃薯D187 光合作用和产量性状相关参数的影响小于热敏感型。
3 讨论
3.1 高温胁迫下不同耐热型马铃薯的结薯特征和株型差异
马铃薯喜性冷凉,其块茎发育对高温尤为敏感,夜间温度超过28 ℃表现为不结薯,且块茎作为马铃薯最主要的经济器官,因此常用在28 ℃的条件下是否结薯来评价马铃薯的耐热性[16]。试验中,耐热型马铃薯在30 ℃下虽然产量降低,但还能结薯,而热敏感型则不结薯(图1)。
较多研究都已证明高温会使马铃薯的株高显著(P<0.01)增加[17-19],本研究表明高温显著(P<0.01)增加了马铃薯的株高和节间长,但未显著改变节数(图4a-c)。由此证明,马铃薯株高增加是由于高温引起节间增长、而不是增加节的数量。叶片角度决定了植株的株型[20],高温胁迫显著(P<0.01)减小了QS9 和D187 的叶夹角(图4f)和叶垂角(图4g),使马铃薯株型由平展型变为紧凑型[21],并缩短了叶片长度(图4i),使马铃薯植株横向生长减弱。较大的叶面积和较多的叶片能够保证植株具有更大的光合面积。试验中,高温使马铃薯的叶面积显著(P<0.01)减小(图4j),削弱马铃薯光合能力。此外,QS9 的叶片数显著(P<0.05)减少,但D187 却增加(图4e),且结合图3 发现,QS9 植株下部叶衰老脱落,而D187 则没有明显的叶片脱落迹象。由此推断,高温下热敏感型马铃薯叶片数减少主要是因为高温加速老叶衰老脱落,而耐热型则是加速新叶长出并减缓老叶衰老来抵御高温伤害。
3.2 高温胁迫对马铃薯植株碳同化能力的影响
马铃薯处于高温环境中时,植株会通过加强水分散失来降低体温[22],因此Tr会显著(P<0.05)升高。本研究发现 2 个不同耐热型马铃薯在高温胁迫下的Tr均显著(P<0.05)升高,且D187 比QS9 具有更高的Tr,但因D187 具有更高的Pn,所以其WUE 也更高,进而保证光合机构合成更多的同化产物。试验中,2个不同耐热型马铃薯的Ci在高温胁迫下均降低,说明高温减弱了马铃薯对CO2的吸收能力[23]。高温胁迫下,QS9因Gs和Ci降低导致Pn降低,而D187 通过增大Gs,加强与外界的气体交换,进而提高Pn,但其Ci却降低,说明耐热型马铃薯在高温胁迫下的净光合速率升高是由气孔因素和非气孔因素共同作用导致,而热敏感型则是由于气孔因素导致净光合速率下降[24]。
AQY 越高、LCP 越低,表示其利用弱光的能力越强;Pnmax和LSP 越高,说明其具有较高的强光利用能力[25]。本试验发现(表2),高温胁迫使2 个不同耐热型马铃薯的Pnmax和LSP 均提高,说明高温胁迫提高了马铃薯对强光的利用能力。高温胁迫下,QS9的LCP 提高,同时AQY 降低;但D187 仅LCP 升高,AQY 不变,说明相较于耐热型,高温胁迫减弱了热敏感型马铃薯对弱光的利用能力,而且LSP 的提高比例远大于LCP,由此表明高温胁迫使马铃薯的光能利用范围扩大。植物为抵御逆境带来的伤害,其Rd会显著(P<0.05)提升[26]。本研究中,2 个不同耐热型马铃薯的Rd均升高,说明马铃薯需要消耗更多的光合产物来减轻高温的伤害。
CE和Vcmax的大小与核酮糖-1,5-二磷酸羧化酶(Rubisco)丰度和活性呈正比关系[27]。本研究表明,高温胁迫对2 个不同耐热型马铃薯的CE 无影响,Vcmax的变化也未达到显著水平。说明抑制块茎形成的高温对马铃薯的Rubisco 活性(或蛋白丰度)及碳同化能力无显著影响。试验中,QS9 和D187 的CCP在高温胁迫下都升高,说明高温胁迫减弱了马铃薯叶片对低浓度CO2的利用能力。RL是植物生命活动能顺利进行的重要保证[28],试验中2 个不同耐热型马铃薯的RL在高温胁迫下都升高,且QS9 升高显著(P<0.05),说明高温增强了马铃薯在光下的呼吸消耗,且热敏感型消耗更多。Jmax体现了RuBP 的再生潜力[29],高温胁迫使QS9 和D187的Jmax降低,表明在高温胁迫下,马铃薯的RuBP 的再生能力受到抑制。前人研究表明,在适宜条件下,植物的Jmax/Vcmax大于4 时,光合电子传递与Rubiso 催化能力间达到动态平衡[30]。本试验中,发现高温胁迫使2 个不同耐热型马铃薯的Jmax/Vcmax值降低,且QS9 下降(4 以下)显著(P<0.01),而D187 则保持在4 以上。表明在高温胁迫下,耐热型马铃薯的电子传递能够满足羧化的需求,但热敏感型不能满足。
3.3 高温胁迫提高了马铃薯对短暂强光的利用能力但打破了光能耗散平衡
光合诱导可用于检验植物对有限光能的利用能力[31-32]。在本研究中(表4),高温胁迫下2 个不同耐热型马铃薯达到最大净光合速率所用的时间均显著(P<0.05)缩短,并且QS9 比D187 所用时间更短。这说明高温提高了马铃薯短暂强光的利用能力,并且热敏感型马铃薯对短暂强光利用率更高。植物接收的光能主要通过3 个途径耗散:光化学反应耗散、非光化学热耗散和荧光耗散,三者为竞争关系,此消彼长[33]。在光响应和光诱导过程中,高温处理使2 个不同耐热型马铃薯的Φf,d较对照无显著变化,但ΦNPQ显著(P<0.05)减少,ΦPSⅡ显著(P<0.05)增加(图6)。说明在高温胁迫下,马铃薯光能耗散失衡,捕获的光能更多用于光化学反应还原力的形成,抵御高温伤害。
3.4 抑制马铃薯块茎形成的高温未造成PSⅡ反应中心受到严重破坏
有研究表明,高温胁迫会造成植物的F0、Fm、Fv/Fm和Fv/F0下降,进而降低净光合速率[34-36]。当PSⅡ的电子传递受到抑制时,Fm会显著(P<0.05)降低,本试验中QS9 和D187的Fm和Fv/Fm在高温胁迫下均显著(P<0.05)下降,说明高温胁迫抑制了马铃薯PSⅡ反应中心的电子传递和光能转换效率[37]。植物PSⅡ反应中心受到逆境破坏时,会引起F0升高,同时Fv/F0显著(P<0.05)降低[38]。试验中,高温胁迫使2 个不同耐热型马铃薯的Fv/F0显著(P<0.01)降低,但F0并未升高,表明马铃薯的PSⅡ反应中心受到影响,但未被抑制块茎形成的高温破坏[39]。
Fv'/Fm'和ΔF/Fm'分别反映了开放的PSⅡ反应中心对激发能捕获效率和实际电子传递效率[40],但ΔF/Fm'比Fv'/Fm'能更准确地反映PSⅡ反应中心的实际情况[41]。本试验中,QS9 和D187 在高温下的Fv'/Fm'在光响应中有部分降低,在光诱导中完全降低(图5c-d),但ΔF/Fm'表现为升高(图5e-f),由此说明马铃薯植株通过提高PSⅡ反应中心的光化学反应效率来适应高温环境。qP表示PSⅡ中开放的反应中心占比及光合电子流动强弱[42],高温处理使马铃薯的qP(图5g-h)和ETR (图5a-b)升高,表明高温提高了马铃薯PSⅡ反应中心的开放程度和电子传递,保证有充足的能量和还原力供给碳同化反应。NPQ 反映热耗散的大小[43],其值变小(图5i-j),表明高温抑制了马铃薯对过剩光能的热耗散。
3.5 高温胁迫下耐热型马铃薯有更高的可塑性
生命体通过某些特征的可塑性响应以改变表型,从而适应新环境。研究表明,可塑性指数越大,植物对环境变化的适应能力越强[44]。试验中,2 个不同耐热型马铃薯的产量特征参数相较于形态特征参数和光合特征参数具有较高可塑性指数和主成分贡献率(图7-图8),这是因为块茎形成对温度变化最敏感,而形态及光合敏感度较弱。对于耐热性不同的马铃薯而言,热敏感型比耐热型在高温下的产量、形态及光合方面受到的影响更大,以避免不适应高温环境而死亡。
4 结论
高温胁迫显著增加了马铃薯植株节间长和株高,但并未显著改变节数和茎粗;此外高温胁迫还显著减小叶片面积、长度和角度,但叶宽长比和小叶数并未显著改变,而且耐热型马铃薯在高温下具有更稳定的叶片数和披垂角。高温胁迫会使马铃薯的Tr加强,WUE 降低,Ci降低和CCP 提高,Rd和RL升高,Jmax降低,黑暗下的叶绿素荧光参数(F0、Fm、Fv/Fm和Fv/F0)降低,而光下的叶绿素荧光参数(Fv'/Fm'、ΔF/Fm'、qP和ETR)升高,达到最大净光合速率所用的时间均显著缩短,ΦPSⅡ增加。此外,热敏感型马铃薯在高温下的Pn和Gs下降,LCP 升高,Vcmax降低,Jmax/Vcmax小于4,而耐热型则表现相反。可塑性和主成分分析表明,高温显著影响马铃薯的形态、光合及产量,且耐热型马铃薯比热敏感型受到的影响较小。
猜你喜欢
青海农林科技(2022年3期)2022-09-26
中国马铃薯(2022年3期)2022-09-13
青海农技推广(2022年1期)2022-05-05
中国马铃薯(2020年3期)2020-07-17
江苏农业科学(2019年11期)2019-07-22
作物研究(2019年2期)2019-04-02
新课程·下旬(2017年7期)2017-08-14
中国马铃薯(2015年3期)2015-04-16
西北林学院学报(2015年5期)2015-01-03
中国马铃薯(2014年3期)2014-02-13
