
重组枯草芽孢杆菌产角蛋白酶的发酵培养基及发酵条件优化
2023-10-17 07:17利刚慧刘俊杰彭帅英梁颖茵鄢陆琪李昆太
中国饲料 2023年19期
利刚慧, 刘俊杰, 彭帅英 , 梁颖茵, 鄢陆琪, 李昆太
(1.广东海洋大学食品科技学院,广东湛江 524088;2.江西农业大学生物科学与工程学院,江西南昌 330045)
角蛋白是动物毛发和甲角的主要成分, 其是自然界中继纤维素类和甲壳素类之后产量第三大的蛋白质类有机聚合物(Bealer 等,2020)。角蛋白富含各类氨基酸,除高达10% ~14%的半胱氨酸外,还含有精氨酸、丝氨酸、脯氨酸、缬氨酸、亮氨酸、苏氨酸、谷氨酸、甘氨酸和天冬氨酸等必需氨基酸(Hassan 等,2020)。 国外将来自家禽加工厂和屠宰场的羽毛、毛发和其他角化副产品归类为第三类动物副产品,这意味着它们属于对人体和动物健康风险程度较低的原材料(Verma,2017)。 因此,角蛋白被视为潜在的优良蛋白质或氨基酸资源,可资源化应用于饲料、肥料、食品、医药等行业。
由于角蛋白分子富含相互交联的二硫键、氢键以及疏水氨基酸,致使其结构极为致密,不仅不溶于水、弱酸、有机溶剂,而且不易被胰蛋白酶或胃蛋白酶等常规蛋白水解酶水解, 给其资源化利用带来极大困难(De Oliveira 等,2020)。目前工业降解角蛋白的方式主要有高温高压水解法、 酸碱水解法和物理膨化法,但这些方法存在能耗高、氨基酸营养损失大和环境二次污染等诸多缺点(Lazarus 等,2021)。 角蛋白酶系一种可特异性降解角蛋白的酶类,不仅绿色环保、经济实用,而且不会破坏氨基酸的天然结构, 在角蛋白降解领域展现出巨大的应用潜力和生态价值(Srivastava等,2020)。 角蛋白酶产生菌的来源与分布十分广泛,在细菌、放线菌和真菌中均发现有角蛋白酶的产生(Gupta 等,2006)。 目前发现的角蛋白酶大多产于细菌,其中以芽孢杆菌属(Bacillus sp.)为主,如地衣芽孢杆菌(B.licheniformis)、枯草芽孢杆菌(B.subtilis)、短小芽孢杆菌(B.pumilus)、解淀粉芽胞杆菌(B.amyloliquefaciens)和蜡样芽孢杆菌(B.cereus)等(Su 等,2020;Li 等,2019)。
然而,产量低、活性差、性质弱一直是制约角蛋白酶工业化生产和应用的问题 (蒋少龙等,2019)。 尤其是野生型产角蛋白酶菌株的酶表达量不高(胡洪,2013),难以满足工业化所需。近年来, 随着分子生物学和基因工程技术的迅速发展,研究学者们对角蛋白酶的分子研究主要集中在角蛋白酶的异源表达和角蛋白酶分子改造两个方面,极大促进了角蛋白酶稳定性和产量的提高(Su 等,2020)。 目前已报道的用于角蛋白酶异源表达的宿主菌主要有大肠杆菌、芽孢杆菌和毕赤酵母等。 其中,枯草芽孢杆菌因其具有完整的蛋白质折叠分泌机制,且在表达来源于芽孢杆菌属的角蛋白酶基因时不存在密码子偏好性问题,在异源表达角蛋白酶方面有着得天独厚的优势(Liu 等,2014)。 例如,Peng 等(2020)将来源于地衣芽孢杆菌BBE11-1 (B.licheniformis BBE11-1)的角蛋白酶基因KerZ1 在枯草芽孢杆菌WB600中分泌表达, 并通过优化工程菌在15-L 罐上发酵产酶的温度、pH、溶氧、补料等过程参数,使得重组角蛋白酶KerZ1 的酶活达到426.60 KU/mL,较原始菌株的角蛋白酶酶活(120.1 U/mL)提高了3552 倍。 Yang 等(2015)将来源于解淀粉芽孢杆菌K11(B.amyloliquefaciens K11)的角蛋白酶基因KerK 在不能降解羽毛的枯草芽孢杆菌SCK6 中进行表达, 重组表达角蛋白酶基因的枯草芽孢杆菌能在24 h 内完全降解羽毛。 Gong 等(2020) 采用aprE 启动子将角蛋白酶基因kerBv在枯草芽孢杆菌中进行异源表达,其酶活性提高了15 倍左右。 本研究以自主构建的角蛋白酶kerJY-23 分泌表达工程菌枯草芽孢杆菌WB600为研究对象,对其产酶的发酵培养基和发酵条件进行优化,旨在为该重组角蛋白酶工业化降解角蛋白提供基础。
1 材料与方法
1.1.1 菌株 重组角蛋白酶工程菌枯草芽孢杆菌WB600-pMA5-kerJY-23,由本实验室构建并保藏。
1.1.2 主要试剂和仪器 5%可溶性角蛋白购自东京化成工业株式会社;LB 肉汤购自北京陆桥技术有限公司; 酵母浸膏购自广东环凯生物科技有限公司; 福林酚试剂购自上海源叶生物科技有限公司;卡那霉素购自北京索莱宝科技有限公司;葡萄糖、蔗糖、氯化钠等化学试剂均购自上海麦克林生化科技有限公司。
立式压力蒸汽灭菌锅(YXQ-100A):上海云泰仪器仪表有限公司;电热恒温水槽(SSW-420-2S):上海博迅实业有限公司医疗设备厂;上海雷磁pH 计(PHS-3C):上海仪电科学仪器股份有限公司;恒温振荡器(HZQ-X500C):上海一恒科技有限公司;紫外可见分光光度计(UV-5500PC):上海元析仪器有限公司;高速冷冻离心机(KDC-160HR):安徽中科中佳科学仪器有限公司。
1.1.3 培养基 种子培养基:胰蛋白胨10 g,酵母粉5 g,氯化钠10 g,蒸馏水定容至1 L,pH(7.0±0.2)。
初始发酵培养基:葡萄糖50 g,蛋白胨20 g,K2HPO41.5 g,NaCl 0.3 g,MgSO4·7H2O 0.025 g,CaCl20.025 g,FeSO4·7H2O 0.015 g,蒸馏水定容至1 L,pH 7.4 ~7.6,121 ℃湿热灭菌20 min。
1.2 试验方法
然而,用哺乳动物瘦素处理银大马哈鱼(Oncorhynchus kisutch)[39]、鲶(Ictalurus punctatus)[40]和绿海鲂(Lepomis cyanellus)[27],却不改变它们的摄食行为或能量代谢。
1.2.1 培养方法 种子制备: 取出于甘油中-80 ℃冻存的枯草芽孢杆菌WB600-pMA5-kerJY-23 菌液,先后于-20 ℃和4 ℃解冻,吸取200 μL 接种于50 mL LB 液体培养基 (含50 μg/mL 卡那霉素)中,37 ℃、200 r/min 培养12 h 获得种子液。
初始发酵条件: 以2% 接种量接种重组枯草芽孢杆菌WB600-pMA5-kerJY-23 种子液于装量为50 mL 初始发酵培养基 (含50 μg/mL 卡那霉素) 的250 mL 三角瓶中,37 ℃、200 r/min 振荡培养48 h。
1.2.2 角蛋白酶活性的测定 参考Yamamura 等(2002)的测定方法并进行适当修改,使用1% 的可溶性角蛋白 (在50 mmol pH 8.0 的Tris-HCl中稀释)作为底物来测定角蛋白酶活性。向1 mL底物中加入1 mL 适当稀释的粗酶液, 在60 ℃下孵育20 min。 加入2 mL 0.4 mol/L 的三氯乙酸终止反应,以12000 r/min 离心5 min,收集上清液1 mL 至试管中,加入1 mL 福林酚试剂和5 mL 0.4 mol/L Na2CO3,混匀后置于60 ℃水浴中孵育20 min,测定660 nm 处的吸光度。 对于空白对照的测定,1 mL 菌液先加入2 mL 0.4 mol/L TCA使酶失活,再加入1 mL 1%可溶性角蛋白。 试验结果为多组试验的平均值。 酶活定义:在上述反应条件下,660 nm 下吸光度值每增加0.01 定义为一个酶活力单位(U/mL)。
1.2.3 单因素试验优化产酶条件 培养基组分的单因素优化: 通过改变发酵基础培养基中的单一组分, 研究不同组分对菌株发酵产角蛋白酶的影响。其中,碳源种类的优化分别采用浓度为50 g/L的葡萄糖、蔗糖、葡萄糖:蔗糖(25:25 g/L)、麦芽糖、可溶性淀粉;有机氮源种类的优化分别采用浓度为20 g/L 的酵母浸膏、牛肉膏、蛋白胨、玉米浆粉、 黄豆饼粉; 硫酸铵添加浓度的优化分别采用0、2.5、5、7.5、10、12.5 g/L;金属离子的优化分别采用添加浓度为0.05 g/L 的K+、Mn2+、Cu2+、Fe3+、Zn2+。
发酵条件的单因素优化: 分别考察不同发酵温度 (29、33、37、41、45 ℃)、 培养基初始pH(6、6.5、7、7.5、8 和8.5)、装液量(250 mL 三角瓶中分别装有40、45、50、55、60、65 mL 发酵培养基)、接种量(2%、4%、6%、8%和10%)、摇床转速(140、160、180、200、220 r/min)、发酵周期(12、24、36、48 h)对菌株发酵产角蛋白的影响。
1.2.4 二次通用旋转组合设计试验 以发酵培养基单因素试验结果为基础,对葡萄糖:蔗糖(X1)、酵母浸膏(X2)、硫酸铵(X3)、MnCl2(X4)四个因素进行4 因素5 水平的二次通用旋转组合设计试验, 用以优化菌株产角蛋白酶的最佳发酵培养基配方。 试验因素水平及编码如表1 所示。

表1 二次通用旋转组合的试验设计因素与水平 g/L
1.2.5 数据分析 试验数据采用Excel 软件进行处理,Origin 2021 软件进行作图分析, 二次通用旋转组合设计的试验结果分析均在DPS 数据处理软件上运行, 试验中各产酶条件的优化均为3次平行试验, 结果以3 次试验的 “平均值±标准差”表示。
2 结果与分析
2.1 发酵培养基组分的单因素优化结果
2.1.1 碳源对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图1 可知,当葡萄糖和蔗糖复配时对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶促进作用最大。 根据不同碳源的酶活力差异性分析, 对其大小进行排序为葡萄糖:蔗糖>葡萄糖>可溶性淀粉>蔗糖>麦芽糖。 单一的蔗糖和麦芽糖作碳源时产酶酶活相对较小,不适合作培养基中的碳源,因而选择葡萄糖和蔗糖复配作为该菌发酵产角蛋白酶的碳源。

图1 碳源种类对枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.1.2 有机氮源对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图2 可见, 酵母浸膏作为氮源时, 重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶酶活最高。根据不同氮源的酶活力差异性分析, 对其大小进行排序为酵母浸膏>玉米浆粉>蛋白胨>牛肉膏>黄豆饼粉。 其中,黄豆饼粉作为氮源时角蛋白酶的表达量最低, 这可能是由于黄豆饼粉为迟效氮源,导致菌株生长代谢较缓慢,从而使得产酶较低。 因而, 选取酵母浸膏作为重组枯草芽孢杆菌WB600 发酵产角蛋白酶的有机氮源。

图2 有机氮源种类对枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.1.3 硫酸铵添加量对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图3 可见,随着硫酸铵添加量的提高,发酵所产的角蛋白酶酶活逐渐增加,当硫酸铵添加量为10 g/L时,角蛋白酶产酶量最大。 当硫酸铵添加量进一步增大至12.5 g/L,角蛋白酶的酶活出现明显下降。因此,重组枯草芽孢杆菌WB600-pMA5-kerJY-23 发酵产角蛋白酶的最佳硫酸铵添加量为10 g/L。

图3 硫酸铵添加量对枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.1.4 金属离子对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图4 可见,当培养基添加氯化锰时,发酵所产的角蛋白酶酶活最高。 根据不同金属离子的酶活力差异性分析,对其大小进行排序为Mn2+>K+>Fe3+>Zn2+>Cu2+,而添加硫酸铜则会对菌株产酶有抑制作用。 金属离子一般作为辅酶成分或充当辅酶功能(丁莉莉,2017),添加氯化锰显著提高了重组枯草芽孢杆菌WB600 发酵产角蛋白酶的酶活,但是额外添加硫酸锌会抑制角蛋白酶产酶, 这与Arokiyaraj 等(2019)的报道相似。

图4 金属离子对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2 发酵条件优化结果
2.2.1 接种量对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 图5 表明,接种量为2% ~6%时,菌株所产的角蛋白酶与接种量呈正相关, 继续增大接种量则导致角蛋白酶产量明显下降。 当接种量为6%时,重组角蛋白酶的酶活达到最大值。 接种量太少,细胞生长延滞,酶的合成受到影响;而接种量过高时,前期细胞生长迅速,营养物质消耗过快,后期供需不足从而会影响酶的合成(Lu 等,2020)。因此,重组枯草芽孢杆菌WB600-pMA5-kerJY-23 发酵产角蛋白酶的最佳接种量确定为6%。

图5 接种量对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2.2 装液量对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图6 可知,在发酵培养基装液量为45 mL/250 mL 时,重组角蛋白酶的酶活达到最高值。装液量会影响到摇瓶发酵的供氧状况, 在重组枯草芽孢杆菌WB600-pMA5-kerJY-23 发酵产角蛋白酶的过程中, 随着装液量不断增大,通气和溶氧随之减小,从而导致角蛋白酶的合成因氧供应不足而受到抑制。

图6 装液量对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2.3 发酵时间对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图7 可见, 随着发酵时间的延长, 重组枯草芽孢杆菌WB600 所产角蛋白酶的酶活先升高而后降低,在第36 h 时所产酶活最高。 发酵时间过长,由于发酵液中营养物质耗竭,导致菌体产生自溶并且累积有害代谢产物,反而使得角蛋白酶酶活有所下降。

图7 发酵时间对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2.4 发酵温度对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图8 可知,温度在37 ℃以下时,随温度上升酶活逐渐提高,在37 ℃时酶活达到最大。 但是当温度继续升高时,由于超过了重组枯草芽孢杆菌WB600 的最适生长温度,菌株生长代谢受阻,导致角蛋白酶的酶活力迅速下降。 因此, 重组枯草芽孢杆菌WB600-pMA5-kerJY-23 发酵产角蛋白酶的最适温度为37 ℃。

图8 发酵温度对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2.5 初始pH 对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图9 可知,pH 从6 ~8 的过程中,酶活随着pH 的升高而逐渐增大,pH 继续升高, 菌株产酶活力有所下降。 在pH 8 时,菌株产酶量达到最高酶活。 大多数细菌产角蛋白酶的最适pH 在中性或者略偏碱性,这与之前报道的Pseudomonas sp.LM19 在pH 8 时产酶最高相似(Mohamad 等,2017)。
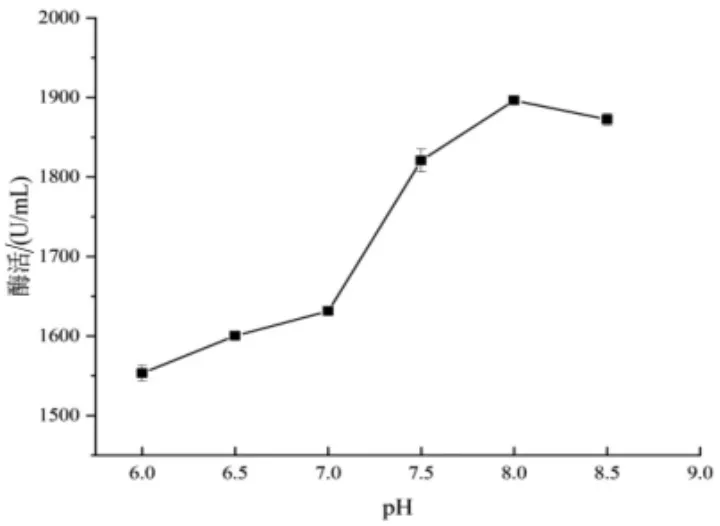
图9 初始pH 对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.2.6 摇床转速对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响 由图10 可知,在摇床转速为140 ~200 r/min 时,重组枯草芽孢杆菌WB600 产酶酶活随着摇床转速的提高而增大。 在200 r/min 的摇床转速下,角蛋白酶的酶活达到最高,继续加大转速至220 r/min,酶活反而有所下降。在发酵过程中,转速会影响物质和溶解氧的交换, 在转速为140 ~200 r/min 时,转速不断增大使发酵溶氧和物质交换增加, 菌株产酶能力增强;当转速增大至220 r/min 后,剪切力也随转速的增加而增大, 制约了菌株生长和产酶(刘婕,2013)。 因此,重组枯草芽孢杆菌WB600-pMA5-kerJY-23 摇瓶发酵产角蛋白酶的最佳转速为200 r/min。

图10 摇床转速对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 产角蛋白酶的影响
2.3 发酵培养基的二次通用旋转正交组合优化与验证 重组枯草芽孢杆菌WB600 发酵培养基中葡萄糖:蔗糖(X1)、酵母浸膏(X2)、硫酸铵(X3)、MnCl2(X4)等组分的二次通用旋转组合设计试验及其结果如表2 所示,方差分析结果如表3 所示。

表2 二次通用旋转组合设计试验方案及结果

表3 试验结果方差分析表
根据表2 所示的试验结果, 以角蛋白酶活为响应值进行多元回归拟合, 得到回归方程:Y=1392.51+73.42X1-124.81X2+86.59X3+100.92X4-141X12+12.21X22+115.80X32+12.15X42+203.96X1X2-174.46X1X3-36.63X1X4。
回归方程中Y 表示角蛋白酶活(U/mL),X1、X2、X3、X4分别表示葡萄糖:蔗糖、酵母浸膏、硫酸铵、MnCl2在培养基中的浓度(g/L)。
根据试验结果进行方差分析可知, 回归方程失拟性检验F1=3.20<F0.05(5,3)=5.41,差异不显著,而显著性检验F2=14.74>F0.05(11,8)=2.95,回归方程显著。 这说明该模型拟合度较好; 在α=0.10 显著水平剔除不显著项后, 简化后的回归方程为:Y=1392.51+73.42X1-124.81X2+86.59X3+100.92X4-141X12+115.80X32+203.96X1X2-174.46X1X3。
2.4 数学模型的应用分析
2.4.1 单因素分析 根据表4 和图11 对单因子试验分析,碳源对酶活的影响最大,其次为硫酸铵添加量、酵母浸膏,氯化锰添加量影响最小。 随着葡萄糖和蔗糖复配添加量的增加, 酶活呈现先增加后下降的趋势。

图11 各单因子与角蛋白酶酶活关系图
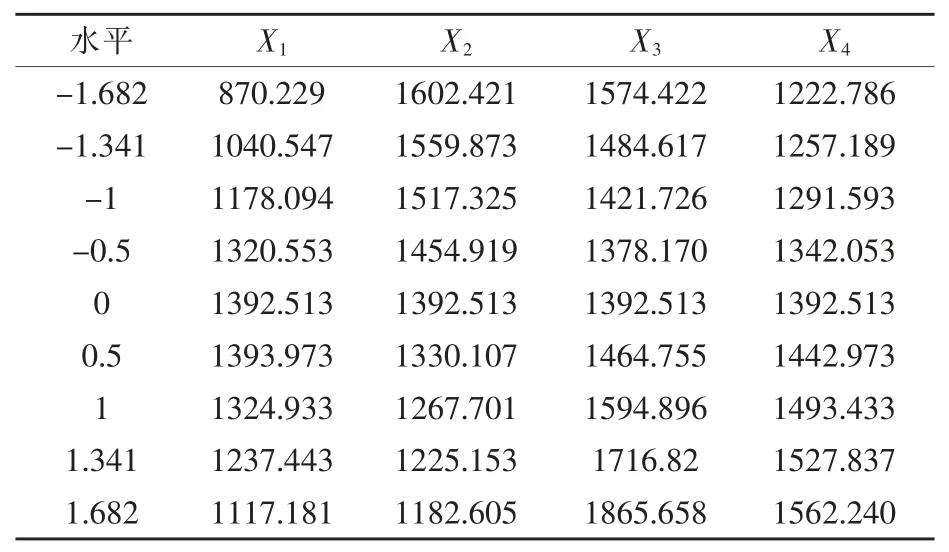
表4 单因子效应分析
2.4.2 产酶优化与验证 根据上述分析,在95%置信区间的优化方案为: 葡萄糖24.95 ~26.15 g/L,蔗糖24.95 ~26.15 g/L, 酵母浸膏18.3 ~20.35 g/L,硫酸铵9.78 ~10.53 g/L, 氯化锰0.05 ~0.06 g/L(表5)。 考虑到实际操作及试剂成本的情况下,将优化方案定为葡萄糖25 g/L,蔗糖25 g/L,酵母浸膏20 g/L,氯化锰0.05 g/L。 对此方案进行试验验证,得到酶活为(2110.33±15.011)U/mL,较优化前提高了5.10 倍。

表5 各变量取值频率分布
3 结论
本研究对重组枯草芽孢杆菌WB600-pMA5-kerJY-23 摇瓶发酵产角蛋白酶的培养基组分和发酵条件进行了优化。 在优化的发酵培养基配方(葡萄糖25 g/L,蔗糖25 g/L,酵母浸膏20 g/L,硫酸铵10 g/L,磷酸氢二钾1.5 g/L,氯化钠0.3 g/L,氯化钙0.025 g/L, 硫酸镁0.025 g/L, 硫酸亚铁0.015 g/L,氯化锰0.05 g/L)和发酵条件(培养基初始pH 8, 装液量45 mL/250 mL 三角瓶, 接种量6%,培养温度37 ℃,摇床转速200 r/min,发酵周期36 h)下,重组枯草芽孢杆菌WB600 所产的角蛋白酶活性达到(2110.33±15.011)U/mL,较优化前提高了5.10 倍。 本研究通过培养基组分和发酵条件的优化,提高了重组枯草芽孢杆菌WB600 发酵产角蛋白酶的酶活, 为该酶后续应用于角蛋白资源的高值化利用提供了一定的理论依据和研究基础。
猜你喜欢
湖南饲料(2021年4期)2021-10-13
工业微生物(2020年3期)2020-06-30
扬子江(2019年3期)2019-05-24
天然产物研究与开发(2019年1期)2019-03-01
天然产物研究与开发(2018年1期)2018-02-02
中成药(2017年5期)2017-06-13
大科技(2016年29期)2016-07-14
中华皮肤科杂志(2014年4期)2014-12-19
中国药业(2014年21期)2014-05-26
中国药业(2014年4期)2014-05-09
