
基于FISH和SNP标记分析百农64对其衍生品种的遗传贡献
2023-10-23 07:40张金龙闫林杰贠艳鸽李云香介艺泽陈向东胡喜贵郭冉冉茹振钢
麦类作物学报 2023年11期
张金龙,闫林杰,贠艳鸽,李云香,介艺泽,陈向东,胡喜贵,周 锋,郭冉冉,茹振钢
(1.河南科技学院农学院/河南科技学院小麦中心/河南省杂交小麦重点实验室, 河南新乡453003;2.河南科技学院资源与环境学院,河南新乡 453003)
骨干亲本在农作物品种选育过程中发挥着重要的作用,自新中国成立到2000年止,我国大约一半的小麦(TriticumaestivumL.)品种是由16个骨干亲本衍生出来的[1]。百农64因其高产、抗病、适应性广,在黄淮麦区累计种植533万hm2,成为黄淮麦区骨干亲本之一,获得了河南省科技进步二等奖。截止2021年,以百农64为亲本,选育出的优良一代衍生品种达30多个,其中子一代衍生品种百农207累计种植面积约430万hm2,获得了河南省科技进步一等奖。
单核苷酸多态性(single nucleotide polymorphism,SNP)标记在各个物种中分布广、易获得且通量高,已越来越多的用于评价骨干亲本对其衍生后代遗传贡献。Jiang等[2]利用小麦90K SNP芯片,解析宁麦9号对其17个衍生品种(系)的遗传贡献,结果表明,除杨辐麦4号外,16个衍生后代与宁麦9号遗传相似系数均超过了0.7。李玉刚等[3]利用SSR和SNP标记分析鲁麦14对其衍生品种青农2号的遗传贡献,SNP芯片结果显示鲁麦14与青农2号的一致性达72.55%,高于另一亲本烟农15,且遗传贡献率大于50%的染色体有18对。白彦明等[4]利用660K SNP芯片分析了4份蚂蚱麦、1份小白麦及其136份衍生品种(系)的遗传多样性,结果表明,在小麦三个基因组中,多态性最高的为B基因组,在7个同源群中,多态性位点数最多的是第3同源群,供试材料间遗传多样性较低,育种家应继续努力丰富小麦遗传资源。高 艳等[5]利用小麦55K SNP芯片,分析周麦22及其80个衍生品种(系)的遗传多样性,结果表明周麦22对其衍生后代的遗传贡献平均为0.658,且衍生系间遗传多样性存在差异。
近年来,已经开发了许多特异单链寡核苷酸(single strand oligonucleotide,SSON)探针,这种探针可由公司合成、分辨率高、成本低,通过一次荧光原位杂交(fluorescenceinsituhybridization,FISH)能够清晰地区分小麦21对染色体[6-9]。杜海梅等[10]利用5个单链寡核苷酸探针,通过非变性荧光原位杂交(ND-FISH)技术,鉴定了小麦-簇毛麦6VS/6AL易位系在9个绵麦37衍生品种中的遗传传递情况,结果显示其传递率为100%。Wu等[11]利用单链寡核苷酸探针套-荧光原位杂交技术(ONMP-FISH),分析了6个6VS/6AL易位系材料及其32个衍生后代品种(系),发现其中27个衍生后代含6VS/6AL易位系。Du等[6]开发了可以清晰地区分小麦21对染色体的SSON探针套,Huang等[8]对该探针套略作修改,利用FISH技术,在373份中国栽培小麦品种中鉴定出14种染色体结构变异,167种染色体多态(block)类型,表明SSON探针在小麦及其近缘物种染色体多样性和结构变异鉴定研究中具有高效性。然而,目前分析骨干亲本对其衍生品种遗传贡献的研究主要集中在农艺性状、品质性状和分子标记水平上,鲜有将SNP芯片和FISH技术相结合,同时从分子和染色体水平揭示骨干亲本对其衍生后代遗传贡献的研究。
本研究旨在通过将FISH和小麦55K SNP芯片技术相结合,探讨百农64对其衍生后代的遗传贡献,揭示生产上选育新品种过程中对染色体片段选择的偏好性,以期为小麦遗传研究和新品种选育过程中的亲本选配提供理论支撑,辅助育种家选育出综合性状好的优良品种。
1 材料与方法
1.1 试验材料
本研究选用百农64(BN64)及其4个子一代衍生品种百农207(BN207)、百农160(BN160)、华育198(HY198)和04中36(04Z36),衍生品种的亲本材料周麦11(ZM11)、周麦16(ZM16)、温麦6号(WM6)共8份小麦材料为研究对象。百农207来源于杂交组合周麦16/百农64,而华育198是从其反交组合百农64/周麦16中选育而成;04中36来源于百农64/周麦11杂交组合;百农160来源于多抗893/温麦6号//百农64/温麦6号。所用试验材料均由河南科技学院小麦中心提供。
1.2 根尖细胞有丝分裂中期染色体制片
小麦种子发根程序参照王丹蕊等[7],根尖分生区细胞用纤维素酶和果胶酶酶解,制成悬浮液滴于载玻片上,有丝分裂中期染色体制片程序参照Zhao等[12]和Komuro等[13]。
1.3 寡核苷酸探针和荧光原位杂交
本研究利用的SSON探针和序列参见Huang等[8],分别为TAMRA(6-carboxytetramethylrhodamine,红色)修饰的pAs1-1、pAs1-3、pAs1-4、pAs1-6、AFA-3和AFA-4,FAM(6-carboxyfluorescein,绿色)修饰的pSc119.2-1和(GAA)10,共8个SSON探针。探针由南京擎科新业生物技术有限公司(TSINGKE biological technology,南京)合成。每张制片的探针用量和荧光原位杂交程序见Huang等[8]。
FISH分析后的根尖有丝分裂中期染色体制片在BX51(Olympus,日本)荧光显微镜下进行镜检,通过Coolcube 1摄像系统及Isis核型分析系统(MetaSystems,德国)等摄取染色体FISH核型高清图像,并进行染色体排队,利用Adobe Photoshop CS6对各个品种染色体FISH核型进行剪切和排列。每个试验材料至少选取6粒种子的根尖细胞进行观察,摄取6~12个染色体形态较好的分裂相,从中选取1个染色体完整且形态良好的分裂相进行核型排队,少数试验材料由于部分染色体重叠,选取了两个分裂相进行染色体排队。
1.4 DNA提取和芯片检测
取各个品种幼叶,每个品种20株混合取样,采用CTAB法[14]提取叶片基因组DNA,将浓度及纯度达标的DNA用于SNP芯片标记检测。利用中国农业科学院作物科学研究所小麦基因组与基因资源研究团队开发的小麦55K SNP芯片,对供试小麦叶片全基因组DNA进行扫描,SNP芯片检测在中玉金标记(北京)生物技术股份有限公司进行。
1.5 数据分析
SNP芯片数据初步过滤由公司完成,设置阈值DQC≥0.82 &&CR≥95进行SNP位点质控,获得基因型原始数据。筛选阈值Best and Recommended=1 &Best Probeset=1 &Conversion Type=Poly High Resolution &call rate≥97的标记,得到初步过滤后的基因型数据;同时删除没有匹配到参考基因组物理位置或匹配到多个物理位置的SNP标记,初步获得32 498个分布于小麦21条染色体上有特异位置的SNP位点。
利用Tassel V5.2.81软件[15](http://www.maizegenetics.net/),去除最小等位基因频率小于1%的SNP标记,共获得23 323个多态性SNP标记用于后续分析。利用Tassel V5.2.81软件,设置检测窗口500 bp,步长100 bp,计算核苷酸多样性指数(nucleotide diversity,Pi)。利用Power marker V3.0 软件[16]计算SNP位点的多态性信息含量(polymorphic information content,PIC)、基因多样性(gene diversity,D)、主要等位基因频率(major allele frequency,MAF),同时计算供试材料之间Nei1983遗传距离,基于该遗传距离构建UPGMA聚类树,使用MEGA V5.02绘制聚类树。遗传相似系数(genetic similarity,GS)采用NTSYSpc V2.10e 软件计算。
2 结果与分析
2.1 百农64对其衍生后代染色体结构变异和多态性的影响
本研究构建了百农64、4个衍生品种及其亲本的SSON探针FISH核型(图1)。与Huang等[8]检测到的小麦染色体结构变异相比,百农64含臂间倒位perInv6B(图1箭头所示),在其衍生品种中仅百农207检测到了此结构变异,且百农207和华育198的6B染色体短臂端部红色信号强,百农64与4个衍生后代间6B染色体FISH核型均有差异。除百农207之外,其他3个衍生品种都含有小麦-黑麦T1RS/1BL易位,且在亲本材料周麦11和周麦16中均鉴定到了此易位系(图1箭头所示)。

绿色信号为FAM修饰的寡核苷酸探针pSc119.2-1和(GAA)10,红色信号为TAMRA修饰的寡核苷酸探针pAs1-1、pAs1-3、pAs1-4、pAs1-6、AFA-3和AFA-4,背景为蓝色(DAPI);箭头所指分别为T1RS/1BL易位系和perInv6B染色体。
分析所研究材料的染色体多态性(图2),共鉴定了48种多态类型(block),B基因组染色体block类型最多,为20种,其次是A基因组有18种,D基因组最少10种(图2)。不同染色体间block类型也有差异,变异范围为1~4;1A和6B染色体的block类型最多,有4种;3A、3D、5D、6D和7D最少,只有1种。其衍生后代中,5A染色体中仅百农160,5B染色体中仅华育198,1B、7A染色体中仅百农207与百农64染色体核型相似;1A染色体仅04中36,2D、4B、4D、7B染色体仅百农160,2A、6A染色体仅百农207与百农64不同;百农160和04中36的3B染色体与百农64不同。

探针颜色同图1。
用“1”和“0”分别表示每个小麦品种21对染色体“是”和“否”为某一种染色体block类型,利用Power marker V3.0 软件计算了供试材料间48种染色体block类型的MAF、D和PIC的值(表1),MAF变异范围为0.500~1.000,平均值为0.797;D值变异范围为0.000~0.500,平均值为0.289;PIC变异范围在0.000~0.375 之间,平均值为0.237。

表1 供试材料间的遗传差异参数比较Table 1 Comparison of genetic diversity parameters among the test varieties
2.2 供试材料多态性SNP位点分布及其多态性
利用Tassel V5.2.81软件筛选后,共获得均匀分布于小麦21条染色体上的23 323个多态性SNP标记用于后续分析。在每条染色体上多态性SNP分布变幅为429~1780,最少的是4D,最多的是2A,平均为1 110个(图3a),不同染色体上SNP分布差异明显。多态性SNP标记在7个染色体同源群中分布变幅为2 569(第6同源群)~3 950(第2同源群)个,按照SNP从多到少分布顺序依次是2>5>7>3>1>4>6(图3b)。在小麦三个基因组中,A和B基因组多态性SNP标记较多,分别为8 504和9 726个,D基因组最少,为5 093个(图3c)。
基于23 323个多态性SNP位点,利用Tassel V5.2.81软件计算得到供试材料核苷酸多样性指数(Pi)变异范围为0.259~0.512,平均值为0.419。利用Power marker V3.0 软件计算了23 323个多态性SNP位点的MAF、D和PIC值(表1),MAF变异范围为0.500~0.938,平均值为0.725;D值变异范围为0.117~0.500,平均值为0.370;PIC范围在0.110~0.375 之间,平均值为0.295。
2.3 百农64 特异SNP位点在其衍生品种基因组上的分布
比较百农64与衍生品种的另外3个亲本周麦11、周麦16和温麦6号间的差异SNP标记,获得3 646个百农64特异SNP位点,其中A、B和D基因组分别为1 341、1 108和1 197个(表2),用于分析百农64特异性SNP位点在其衍生后代染色体上的分布。分别比较4个衍生品种与百农64的相同SNP位点比例(SSLR),结果显示,同一衍生品种不同染色体之间的SSLR差异明显,比如04中36,3D染色体仅为1.6%,而4B染色体高达100.0%;同一染色体不同品种之间的SSLR差异也明显,以6B染色体为例,华育198为10.0%,而百农207高达92.7%。4个衍生后代与百农64在21对染色体上SSLR平均值变幅为21.5%~85.4%,其中2B,3D,6D低于30.0%;在30.1%~50.0%之间的最多,有12对染色体;超过50.0%的有6对染色体,其中6A染色体达到了85.4%。A、B和D三个基因组之间SSLR平均值也有差异,分别为54.2%、46.9%和36.5%。除百农160外,其它3个衍生品种的21对染色体SSLR平均值均超过了50.0%。
2.4 百农64及其衍生后代的遗传相似性分析
将供试材料FISH鉴定的48种染色体block类型作为细胞学标记,结合23 323个多态性SNP位点,共23 371个标记,利用NTSYSpc V2.10e软件计算了供试材料间的GS值(表3)。百农207、华育198和04中36三个衍生品种与百农64的GS都超过了0.700,华育198最高为0.762,这表明百农64对其后代遗传贡献率高。百农160与百农64的GS值为0.564,而温麦6号对百农160贡献较高,两者之间GS为0.613。四个衍生系之间的GS变幅为0.464~0.652,平均值为0.544,华育198与百农207之间GS最高,为0.652,华育198与百农160之间GS最低,为0.464。

表3 供试材料间遗传相似系数Table 3 Genetic similarity of the test varieties
2.5 聚类分析
基于供试材料染色体多态性block类型和多态性SNP共23 371个标记,利用Power marker V3.0软件,计算了Nei1983遗传距离,并根据遗传距离构建了UPGMA聚类树,在Nei1983遗传距离0.20处,可分为三大类群(图4)。百农64及其4个衍生后代均属于类群Ⅰ,华育198与百农64遗传距离最近。温麦6号也聚在了类群Ⅰ,且百农160与其遗传距离较近,这与其系谱及遗传相似系数相吻合。周麦11和周麦16各为一类,与其它材料遗传距离相对较远。Nei1983遗传距离与遗传相似系数结果一致。
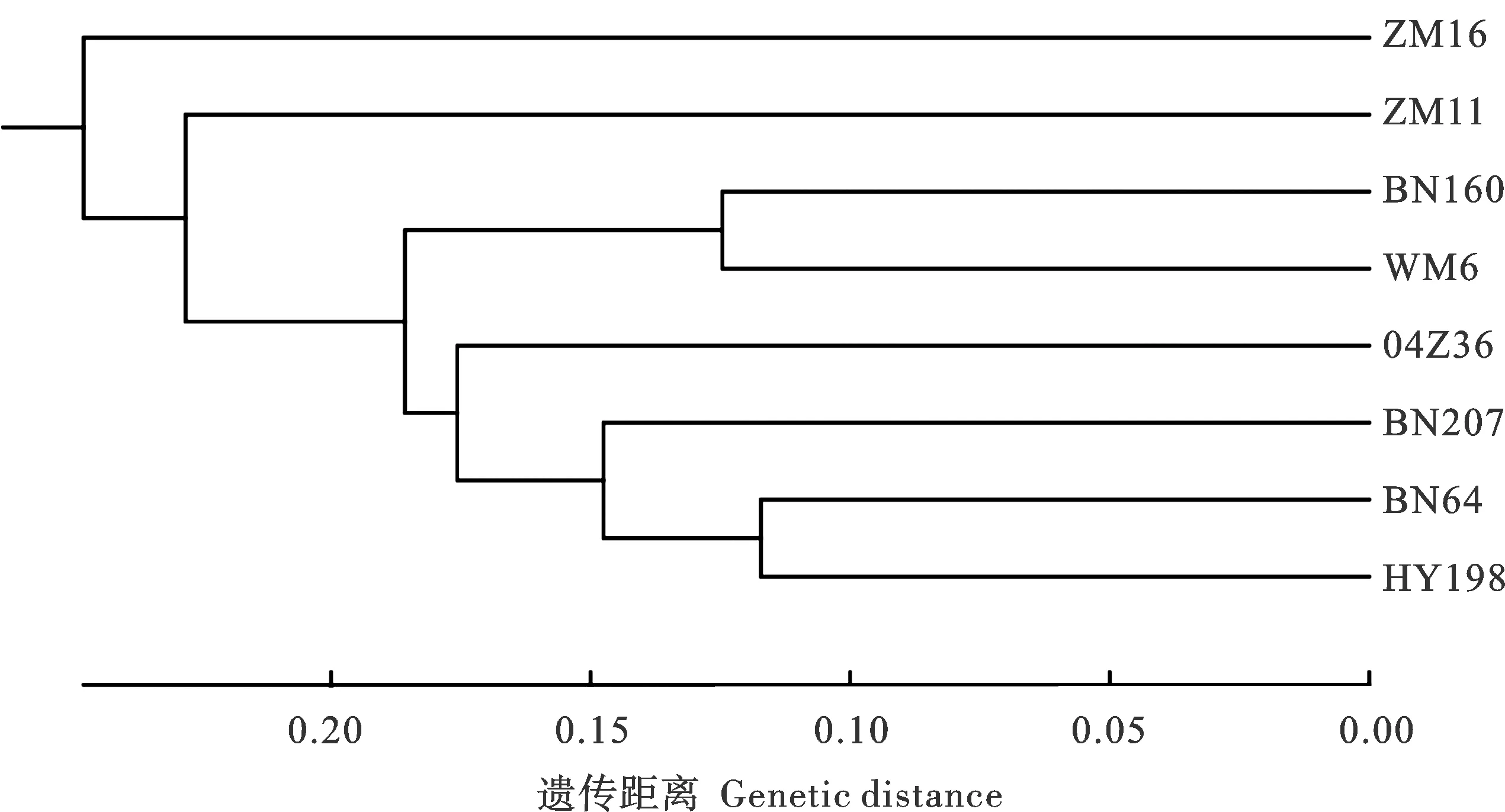
图4 供试材料基于48种染色体多态性block和23 323个多态性SNP标记的聚类分析
3 讨论
依据重复序列探针在不同小麦品种的丰度和分布不同,利用FISH技术可以清晰地检测染色体结构变异和多样性,对解析小麦基因组结构和组成具有重要参考价值[7-8]。利用pSc119.2和pAs1两个重复DNA质粒探针,对21个小麦品种进行FISH分析,以中国春(CS)核型作为参考,鉴定到7对B组染色体均存在染色体多态性[17]。Huang等[8]对CS和373份中国栽培小麦品种进行FISH分析,从中鉴定出167种染色体多态(block)类型,其中A组和B组较多,分别为78种和64种,而D组只有25种。Carvalho等[18]利用(AAC)5作为探针,对葡萄牙5个面包小麦和5个硬粒小麦进行FISH核型分析,发现5个面包小麦的1B和6B染色体具有多态性。本研究与前人研究结果相似,A和B基因组染色体多态Block类型差异不大,D基因组最少,这表明D基因组序列比较保守。本研究在百农64中鉴定出了臂间倒位perInv6B,且遗传给了其子代品种百农207,在百农64及其衍生品种间6B染色体多态性较丰富。同时,本研究在亲本材料周麦11、周麦16和3个衍生品种中均鉴定出了小麦-黑麦T1RS/1BL易位,表明华育198和04中36的T1RS/1BL易位分别来自其亲本周麦16和周麦11。百农160含T1RS/1BL易位,而其亲本百农64和温麦6号都不含,其易位可能来自另一亲本多抗893。小麦育种过程中,小麦-黑麦T1RS/1BL易位系被育种家广泛选择。本研究表明FISH核型可以追踪小麦品种系谱。
本研究利用小麦55K SNP芯片,对百农64及其衍生品种进行全基因组扫描,结果显示在3个基因组中,B基因组多态性SNP标记最多,A基因组与B基因组差异不大,D基因组最少,且D与A和B两个基因组差异较大,这与本研究的FISH染色体多态性block类型结果一致,同时与之前大多数研究结果一致[4,19-20]。这与D基因组来源单一,序列比较保守有一定关系[19],且在六倍体小麦进化过程中,A和B基因组组装到一起形成四倍体的时间较早,而AABB与D基因组结合形成六倍体栽培小麦的时间较短[21]。Jia等[21]通过对小麦D基因组供体材料节节麦(AegilopstauschiiL.)进行全基因组测序分析,发现其基因家族与抗病性、非生物胁迫及谷物品质相关,而育种家在品种选育过程中,偏向于选择具有这些优良性状的品系,这种长期的定向选择导致了其遗传多样性比A、B基因组低。白彦明等[4]、曹廷杰等[19]和刘易科等[20]研究均表明,小麦第四同源群多态性SNP标记最少,且4D染色体上的多态性SNP标记最少,而本研究结果显示第四和第六同源群多态性标记数均少,多态性标记最少的染色体同样是4D染色体。已有报道显示4D染色体上携带控制育种目标性状的基因较多,多年来育种家的定向选择导致了4D染色体遗传多样性降低[19]。
FISH技术可以从细胞学上鉴定染色体结构变异和多态性,而染色体结构变异和多态性在品种环境适应性、抗病性、抗逆性、驯化和优良农艺性状形成等过程中发挥的作用比SNP大;SNP标记可在单核苷酸水平上检测物种遗传差异,且在各个物种全基因组水平上均有广泛分布。将这两项技术结合起来,可以用于构建重组自交系群体遗传图谱[12],也可进行染色体结构变异和农艺性状的关联分析[22],但鲜有将两种技术结合起来研究骨干亲本遗传贡献的报道。本研究将FISH和SNP芯片技术相结合,从细胞学和单核苷酸等不同水平分析百农64与其衍生一代品种之间的遗传相似性。除百农160与百农64遗传相似系数小于0.6外,其它的品种都超过了0.7,且百农64特异性SNP位点在百农207、04中36和华育198染色体上分布的平均比例均超过了50%,这与核型分析结果一致,这3个品种分别有16、14、17对染色体核型与百农64一致。百农64及其4个衍生品种聚为一类,聚类分析结果与遗传相似系数结果一致。SNP标记和核型分析结果均表明百农64对这些衍生品种的遗传贡献较高,生产上育成新品种的遗传构成不是理论上的双亲平均值,而是偏向于综合性状好的骨干亲本,骨干亲本许多优良性状被育种家定向选择,遗传给了子代[23-24]。对百农160遗传贡献率最高的亲本是温麦6号,两者之间遗传相似系数为0.613,聚类分析结果显示百农160与温麦6号遗传距离最近,这是因为百农160是以温麦6号为共同亲本与另外两个亲本进行三亲本双交选育而成的,这些研究结果与其系谱信息相吻合。
猜你喜欢
作物学报(2022年9期)2022-07-18
耕作与栽培(2022年1期)2022-04-07
长江蔬菜(2021年12期)2021-04-04
中国瓜菜(2019年4期)2019-07-03
中国果业信息(2019年11期)2019-01-05
现代检验医学杂志(2016年3期)2016-11-15
西南农业学报(2016年5期)2016-05-17
三峡大学学报(自然科学版)(2016年6期)2016-04-16
物理实验(2015年9期)2015-02-28
新疆农垦科技(2014年10期)2014-02-28
