
多轮次富营养模拟富集对窖泥富集液代谢产物的影响
2023-12-29 04:49叶光斌宗绪岩
酿酒科技 2023年12期
李 栋,叶光斌,2*,宗绪岩,2,邹 伟,2
(1.四川轻化工大学生物工程学院,四川宜宾 644000;2.四川轻化工大学酿酒生物技术及应用四川省重点实验室,四川宜宾 644000)
我国的白酒文化可追溯到四五千年前的“龙山文化”时期,可谓历史悠久[1]。浓香型白酒作为我国的第一大蒸馏酒,其酒体具有绵甜醇厚、窖香浓郁、舒适协调等特点,深受人们喜爱,且市场份额占比大[2]。窖泥作为浓香型白酒生产过程中主要微生物来源之一,其质量对白酒品质改善及特质香气的形成具有重要的影响。泥窖池作为浓香型白酒特有的发酵容器,也是优良酿酒微生物栖息繁衍地,经过长期高酸度和密闭厌氧发酵等特殊酿酒环境的驯化,窖泥中逐渐富集了大量具有产香功能的微生物[3]。
在成熟的纯培养技术下,窖泥中的产香功能微生物也逐渐被发掘出来。己酸乙酯是浓香型白酒的主要风味物质[4],己酸作为己酸乙酯合成的前体物质,其产生菌种类较多。主要有梭菌属中的克氏梭菌(Clostridium kluyveri)、广西梭菌(Clostridium guangxiense)[5-10];芽孢杆菌属中的地衣芽孢杆菌(Bacillus licheniformis)、巨大芽孢杆菌(Bacillus megaterium)[11];颤杆菌科的Caproicibacterium菌属[12];产己酸菌属(Caproiciproducens)[13];瘤胃球菌科CPB6 菌株[14]等。有研究表明,多数梭菌可通过逆β 氧化途径,以乙醇或乳酸为基础,通过乙酰辅酶A,最终缩合成己酸,而其他菌属产己酸的代谢途径却鲜有报道[15-16]。
除己酸外,丁酸、乙酸等酸也对白酒的风味影响颇深。对酪酸梭菌(Clostridium butyricum)[17-18]、毛螺旋菌科(Lachnospiraceae)JN500902T 菌株[19]的研究发现,其可产生多种脂肪酸,如丁酸、乙酸等;在白酒酿造中,丁酸被转化为丁酸乙酯,乙酸被转化为乙酸乙酯,两者皆是白酒中重要的香味成分。也有研究发现丁酸可作为合成己酸的前体物质,同时对于新窖泥生成正丁醇起到至关重要的作用[20-21]。甲烷菌可实现“种间氢转移关系”,与己酸菌能够形成共栖作用,从而更加有利于己酸的生成[22]。
在窖泥环境体系中,微生物十分丰富,种间关系复杂,代谢产物多样。加之实验室条件下可培养的微生物种类有限,因此通过分离培养的方式必然会忽略窖泥中绝大部分具有产酸功能的微生物类型[23]。富集培养是目前有效分离环境样品中难培养微生物的有效方法,例如Wu 等[24]从富集培养物中分离到了许多针对环境污染物的降解菌;Pernicova 等[25]将活性污泥加入到富集培养基(MSM)中培养,分离出了能产聚羟基链烷酸酯(PHA)的芽孢杆菌。对于微生物的分离而言,培养基选择与设计无疑具有重要的作用。窖泥在不同富集处理下发酵产物的变化规律研究对于功能发酵菌液的开发与难培养微生物的分离等都具有重要的意义。如在培养基富集微生物时,进行热激的前处理,就会使得大量非孢子形成的细菌减少,降低微生物的物种丰富度,从而使多数含有芽孢的梭菌、芽孢杆菌等得以生存[26];在培养基组分上加入乙醇作为碳源,富集窖泥后发现代谢产物乙酸的含量会高于己酸的含量,而以葡萄糖作为碳源时,己酸含量则高于乙酸含量[16]。尽管现在富集培养应用领域十分广泛,也帮助学者分离或富集了众多的新种和目的功能菌,但是在白酒上的研究还是较少,限制我们对中国浓香型白酒发酵过程中窖泥微生物驱动原理的了解,所以窖泥中功能微生物的开发、难培养微生物的富集等工作仍是我们现在首要的目标。
本研究设计不同的培养基营养成分和是否使用热激处理等不同的处理组合(具体处理见表1),通过比较不同处理下6 轮次发酵产物确定最佳发酵处理措施。相关研究结果可以为窖泥功能菌液功能导向性富集、富集产酸培养条件优化、窖泥老熟、难培养微生物的分离等提供理论支撑。

表1 培养基成分表
1 材料与方法
1.1 材料、试剂及仪器
1.1.1 窖泥样本
优质老窖泥取自泸州某名优酒厂,用冰盒迅速运回实验室,于4 ℃冰箱保存,用其配置成富集液样品1—20号。
1.1.2 主要仪器与设备
气相色谱-质谱联用仪,美国Agilent 科技有限公司;厌氧工作站,北京八方世纪科技有限公司。
1.1.3 培养基
窖泥用不同培养基进行富集培养,培养基组分见表1。
1.2 试验方法
1.2.1 窖泥的富集
取25 g 窖泥于厌氧工作站中接种至250 mL 已除氧液体培养基(1.1.3)中,所有样品于35 ℃培养7 d 后,以10 %的接种量进行转接,共富集6 轮(详见图1)。将每轮富集液分别保存至4 ℃及-20 ℃冰箱中,用于后续代谢产物研究。
图1 窖泥富集培养接种方法
1.2.2 窖泥富集液代谢产物
取发酵液20 mL,使用2 mol/L 硫酸酸化至pH2,加无水乙醇定容至25 mL,12000 r/min、4 ℃冷冻离心5 min,取上清液,过0.2 μm 微孔滤膜[30]。进样前加10 μL乙酸丁酯(84 mg/mL)作为内标混匀。
气相色谱条件:DB-WAX(60 m×250 μm×0.25 μm)色谱柱;载气为高纯度He,流速1 mL/min,进样口温度230 ℃;程序升温:初始温度为60 ℃保持1 min,8 ℃/min 的升温速率升至180 ℃保持2 min,再以15 ℃/min升至230 ℃,保持5 min。
质谱(MS)条件:电子离子源(EI),70 eV 电子能量,采集模式为全扫描,质量范围20~550 u,离子源温度230 ℃,四级杆温度150 ℃,接口温度230 ℃。
定性分析:质谱图通过与美国Agilent 公司提供的美国国家标准技术研究所(national institute of standards and technology,NIST)标准谱库05a.L 进行比对,选择匹配度均>80的进行定性分析[31]。
半定量分析:以乙酸丁酯作为内标,测定样品中酸类、酯类、醇类及芳香类化合物的峰面积与乙酸丁酯峰面积之比,再根据乙酸丁酯的质量浓度进一步计算其含量[32]。
2 结果与分析
2.1 窖泥富集液代谢产物
窖泥富集液在不同培养基条件下经过六次富集后,其代谢产物数据显示(图2),总酸、乙酸、丁酸和己酸产量范围分别为61.8~11392 mg/100 mL、0~3491 mg/100 mL、0~9695 mg/100 mL、0~1744 mg/100 mL,对于能起到富集酸作用的培养基而言,总酸富集趋势为第一轮富集到第三轮富集呈快速上升,第三轮富集到第四轮富集呈缓慢上升,而在第四轮富集到第六轮富集时则呈现下降,可以看出浓香型白酒的主要挥发性酸积累是在第三轮、第四轮富集时得到最大积累量(最高能到11392 mg/100 mL)。因此我们可以将整个发酵过程分为三个阶段,第一阶段为微生物生长繁殖时期(第一轮、第二轮富集时期),第二阶段为微生物代谢时期(第三轮、第四轮富集时期),第三阶段为微生物衰退消亡时期(第五轮、第六轮富集时期)。
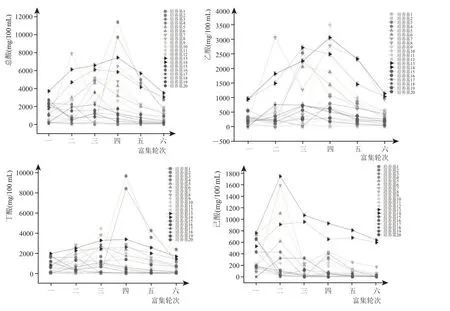
图2 样品富集总酸图
2.2 热激与未热激对同组分培养基间的影响
第三轮、第四轮富集时期,为微生物代谢时期,且在该时期有大量的酸积累。所以,将同培养基条件下,第三轮、第四轮富集时期的总酸积累作为分析基础,对其热激与未热激处理下的样本做显著性分析,如图3 所示,相同培养基组分在不同处理结果下,热激与未热激对酸的富集不呈显著性差异,所以认为热激与未热激的前处理对微生物发酵中期产酸的影响很小。
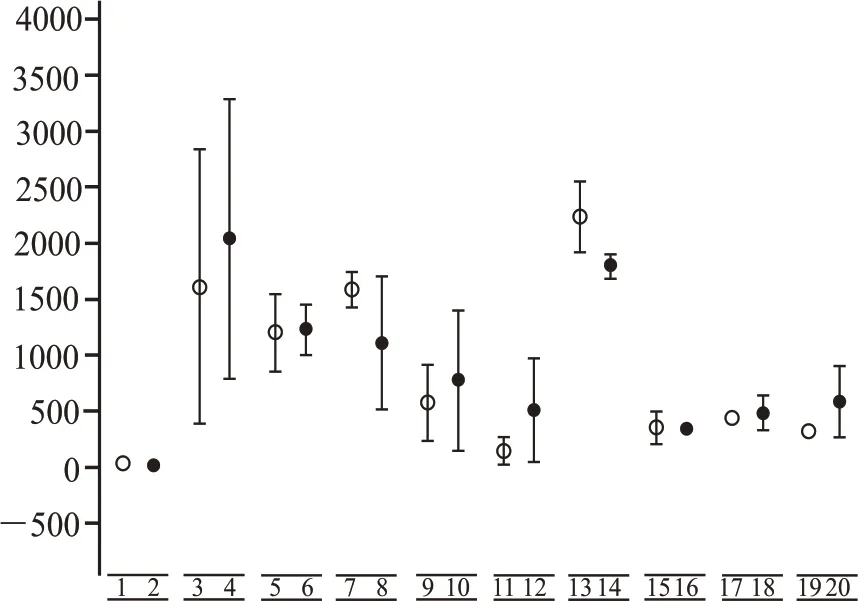
图3 同培养基组分下,热激与未热激处理下微生物的富集与第三轮、第四轮发酵时期总酸积累的显著性关系
2.3 培养基成分对窖泥富集液代谢产酸的影响
虽然在主发酵时期,是否热激处理对同组分富集样本影响较小,但是培养基自身的成分组成对代谢产物的影响极其重要。对此,针对主发酵时期的各个酸代谢量同20 组不同处理的富集样本的营养物质进行相关性冗余分析(RDA 分析),如图4 所示,RDA 分析显示,培养基中的营养物质(硫酸铵、蛋白胨、酵母粉、乙酸钠、丁酸钠、可溶性淀粉)与总酸、乙酸、丁酸、己酸呈正相关关系,乳酸、黄水、乙醇、葡萄糖、巯基乙酸钠、牛肉膏则与上述酸呈负相关关系。
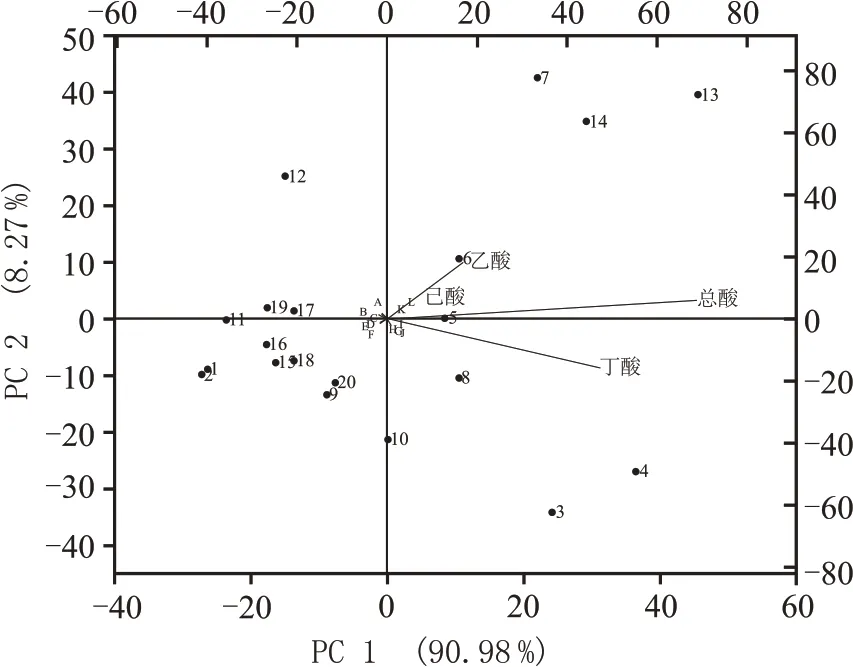
图4 不同处理下,富集培养在第三轮、第四轮富集时培养基的主要组分和总酸、乙酸、丁酸、己酸的冗余分析(RDA分析)
3号、4号、5号、6号、7号、8号、13号、14号处理与总酸、乙酸、丁酸、己酸呈正相关关系,这些酸也与上述处理中的营养物质(3 号、4 号、5 号、6 号、13号、14 号处理共同的营养组分:乙酸钠、硫酸铵、蛋白胨、酵母粉)呈正相关,唯独与7 号、8 号处理中的黄水呈负相关,但这些处理都在富集过程中积累了较好的混合酸(总酸、乙酸、丁酸、己酸)。相关研究发现,在葡萄糖CGM 培养基(本实验5 号处理使用的培养基)中加入丁酸钠和乙酸钠后,能有效提高丁酸、乙酸的产量,进而影响己酸的积累(从654 mg/100 mL 提高至892 mg/100 mL)[16],而3号、4 号、5 号、6 号处理中的丁酸钠和乙酸钠与混合酸呈正相关关系,表明丁酸钠和乙酸钠对混合酸的积累有着促进作用(图2,图4)。其表现为,3号、4 号处理在第四轮富集时,积累了较多的总酸和丁酸(9705.2~11392 mg/100 mL 和8435~9695 mg/100 mL),5 号、6 号处理(独特的营养组分:丁酸钠)在第三轮富集时对总酸、乙酸和丁酸有一定积累(4618~5083 mg/100 mL,2061~2259 mg/100 mL,2351~2687 mg/100 mL)。黄水作为浓香型白酒发酵过程中的产物[33],推测其含有较丰富的促生长因子,所以在加入了黄水的7号、8 号处理中表现出较高的乙酸、丁酸积累。其中7 号处理在第二轮富集时积累了较多总酸、乙酸和己酸(7892 mg/100 mL、3067 mg/100 mL 和1587 mg/100 mL),8 号处理则在第三轮富集时主要对总酸、丁酸有所积累(5159 mg/100 mL,3817 mg/100 mL)。己酸乙酯是浓香型白酒中的主体呈香物质,过多的乳酸会抑制浓香白酒主体香的形成,所以现在的研究推行通过“增己降乳”的方法来提高己酸乙酯的含量[34],1 号、2 号、11 号、12 号处理中富含乳酸,并且与总酸、乙酸、丁酸、己酸呈负相关关系,因此推测过高的酸度对微生物代谢形成了反馈抑制,导致了产酸量的减少。Clostridium kluyveri可以利用乙醇来产己酸,但在窖泥菌群研究中,该菌的相对丰度不高[35],本实验中的9 号、15号、16 号、17 号、18 号处理均加有乙醇,且与总酸、乙酸、丁酸、己酸呈负相关关系,同时产酸(如己酸)较低(平均酸含量<1336.4 mg/100 mL),所以推测乙醇能抑制混菌的生长或能抑制以乙醇为碳源生长的细菌(如Clostridium kluyveri)。有研究表明,巯基乙酸钠作为抑制剂,可以抑制微生物芽孢的形成[36],所以认为15 号、16 号处理中加入的巯基乙酸钠,也会使微生物的含量降低,从而使得产酸很低。10号处理中加入乙醇后,依然在第二轮富集中表现出较高的丁酸含量(4465 mg/100 mL),可能因为该培养基在灭菌后加入乙醇,然后又进行了热激处理,使得乙醇挥发(沸点:78 ℃),不用抑制该培养基的微生物生长。19 号、20 号处理对挥发性酸积累很低(平均酸含量<1336.4 mg/100 mL),可能因为该培养基(MRS 培养基)主要是用来培养乳酸杆菌的,所以使得其它产酸微生物难以生存。13号、14 号处理(独特的营养组分:可溶性淀粉、葡萄糖)能在第三轮、第四轮富集时积累总酸、乙酸、丁酸、己酸(5843~7444.7 mg/100 mL,2254~3055 mg/100 mL,2452~3383 mg/100 mL,652~1067 mg/100 mL),而且混合酸的含量在富集过程中有先增加后下降的趋势,说明富集中期时,有利于这些处理对混合酸的积累。
2.4 热激与否对优势富集样本的各挥发性酸的影响
经过多组处理间的总酸比较,筛选出了13 号、14号处理作为最优的富集培养基组合,为进一步探究热激对其各挥发性酸的影响,我们对其微生物代谢时期的各酸代谢量进行了差异分析,如图5 所示。对比之下,13号、14号处理,在微生物代谢时期(第三轮、第四轮富集)产混合酸差异很小,两轮富集乙酸、丁酸、己酸差距分别在225~801 mg/100 mL、116~155 mg/100 mL、141~301 mg/100 mL 之间,并无显著性差异,且产混合酸能力很高。由于13号、14 号处理在多轮次富集中产酸高且稳定,受热激处理影响小,所以我们认为其可作为窖泥产酸核心菌群研究的出发样本。
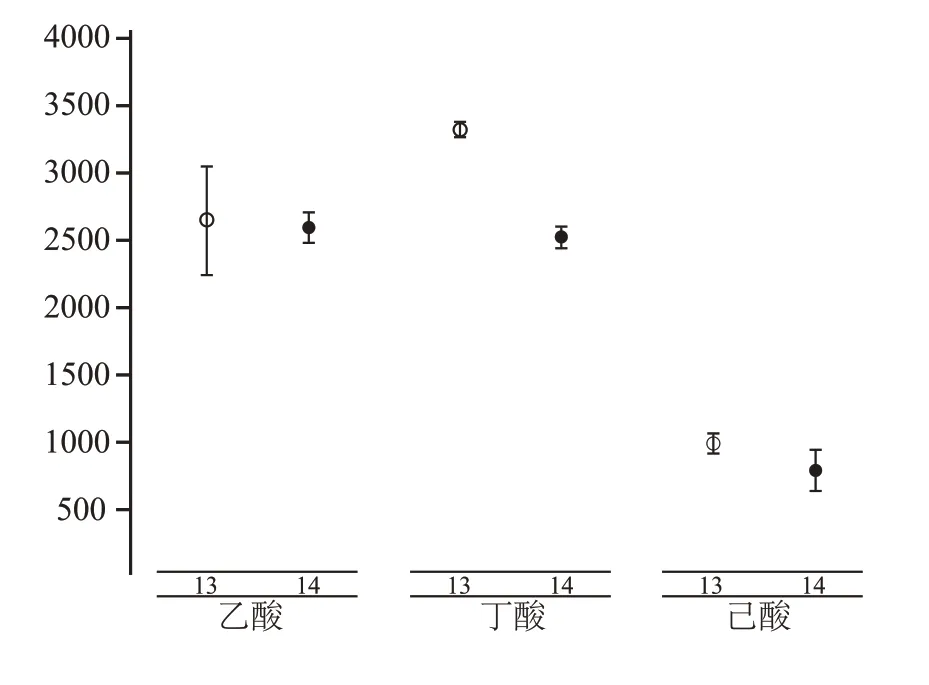
图5 13号、14号处理第三轮、第四轮富集时,富集轮次与酸之间的显著性关系
3 结论
本实验通过不同培养条件对窖泥进行连续富集培养,筛选出了能够高产酸的13 号、14 号处理。在发酵第三轮、第四轮时,13 号处理和14 号处理间无显著性差异,且都能够有效的富集混合酸(乙酸、丁酸、己酸)。在最佳产酸时期,总酸的产值在5000~7500 mg/100 mL之间,乙酸在2254~3055 mg/100 mL之间,丁酸在2452~3383 mg/100 mL之间,己酸在909~1744 mg/100 mL 之间。该项研究成果,后续可作为窖泥产酸核心菌群的研究对象,也可以开展纯菌的分离研究。同时本课题相关研究可以为后续单菌分离、改善窖泥质量、生物强化法生产浓香型白酒等提供理论依据及实践基础。
猜你喜欢
中国酿造(2023年9期)2023-09-28
能源与环境(2022年5期)2023-01-10
山西化工(2022年1期)2022-03-08
中国酿造(2021年12期)2021-03-04
煤炭与化工(2021年1期)2021-02-26
中国当代医药(2019年20期)2019-09-25
农业机械学报(2019年1期)2019-02-15
中国调味品(2018年2期)2018-03-06
军事文摘·科学少年(2016年10期)2016-12-08
中国美容医学(2016年4期)2016-07-25
