
Inf luence of nitrogen status on fermentation performances ofnon-Saccharomyces yeasts: a review
2024-02-15 07:58JinchenLiMengmengYuanNanMengHeheLiJinyuanSunBaoguoSun
食品科学与人类健康(英文) 2024年2期
Jinchen Li,Mengmeng Yuan,Nan Meng,Hehe Li,Jinyuan Sun,Baoguo Sun
Key Laboratory of Brewing Molecular Engineering of China Light Industry, Beijing Key Laboratory of Flavor Chemistry, Beijing Technology &Business University, Beijing 100048, China
Keywords: Non-Saccharomyces yeasts Nitrogen Fermentation kinetics Nitrogen preference Wine aroma
ABSTRACT Nitrogen,one of the most crucial nutrients present in grapes and musts,plays a key role in yeast activities during alcoholic fermentation.Such inf luences are imposed on yeast growth and fermentation performances including the formation of secondary metabolites.Saccharomyces cerevisiae,the main yeast responsible for fermentation,has been studied extensively regarding nitrogen impacts.On the other hand,a similar study for non-Saccharomyces yeasts,whose contributions to winemaking have gradually been acknowledged,remains to be fully explored,with a few studies being reported.This review starts by discussing nitrogen impacts on non-Saccharomyces yeast growth and fermentation kinetics in different case scenarios,then proceeds to summarize the nitrogen preferences of individual yeast strains with regulation mechanisms elucidated by recent studies.Detailed discussions on the influences on the production of volatile compounds and proposed pathways therein are made,followed by future work suggested as the f inal section.In summarizing the nitrogen impacts on non-Saccharomyces yeasts throughout alcoholic fermentation,this review will be helpful in obtaining a more comprehensive view on these non-conventional wine yeasts in terms of nutrient requirements and corresponding volatile production.Research gaps will therefore be elucidated for future research.
1.Introduction
Wine,the product of the successful cooperation between grape juice and yeasts,has long been receiving much attention both from dining tables and laboratory benchtops.Saccharomyces cerevisiae,the major wine yeast responsible for alcoholic fermentation,was previously considered the only responsible microbial participant,and further contributions apart from converting sugar to alcohol,such as producing secondary metabolites including aroma compounds,have gradually been recognized and investigated.Nutritional requirement,as an important factor contributing to yeast performances during fermentation,was specifically studied for their impacts on fermentation kinetics and metabolite production.On the other hand,lengthy process applied to the exploration of the involvement of non-Saccharomycesyeasts in grape juice fermentation.Such yeasts have been treated as spoilage yeasts in the wine industry for long before their positive contributions were finally acknowledged.The vast diversity of non-Saccharomycesyeasts and versatile metabolic abilities render them a useful tool to highlight characteristics of different grapes and/or regions via distinctive aroma production,and to address problems such as wine haze[1].However,unlikeS.cerevisiae,which functions well in wine-like conditions,most non-Saccharomycesyeasts are unable to tolerate osmotic stresses such as high ethanol contents and therefore fail to complete alcoholic fermentation.Co-inoculation and sequential inoculation approaches were applied,of which non-Saccharomycesyeasts are inoculated f irst/together withS.cerevisiaeresulting in mixed populations during fermentation.S.cerevisiaeusually takes over and finishes fermentation as the alcohol content increases.
Countless reports are presently focusing on the contributions of various non-Saccharomycesyeasts from different wine regions to the complexity and distinctiveness of wine aroma profiles.However,inter/intra-specific diversity has also been observed among these yeasts,adding difficulty to characterize the universal features of yeasts for further applications.Increasing interests in such topics have arisen recently,among which,influences of abiotic factors on yeast performances during fermentation are of particular concern.Abiotic factors,including nutrients,play crucial roles in determining yeast growth and potentially metabolic pathways during fermentation.Nitrogen,as one of the major nutrients naturally occurring in grapes,imposes great influence on yeast growth as well as aroma production.And such influence may add complexity to mapping yeast fermentation performance.Metabolic pathways and regulatory mechanisms of nitrogen sources inS.cerevisiaehave been extensively studied and reviewed[2-3],rendering it a rational strategy as nitrogen management for desirable wine character production,such as aromas.Similar work has been initiated in recent years on non-Saccharomycesyeasts for the same reason,with further aims to study nitrogen impacts as well as common metabolic features of the yeasts for more efficient and wider applications.
Therefore,aiming at gaining a clear understanding of how nitrogen impacts wine aroma production by non-Saccharomycesyeasts,this review starts with nitrogen impact on non-Saccharomycesyeasts in terms of their fermentation kinetics,and then proceeds to summarize non-Saccharomycesyeast-related aroma profiles under nitrogen influence.Moreover,potential metabolic mechanisms therein will also be reviewed.Such discussions will be helpful in acknowledging correlations between nitrogen status and aroma production of non-Saccharomycesyeasts,and shed light on potential research gaps for future investigations.
2.Nitrogen influence on fermentation kinetics of non-Saccharomyces yeasts
Based on its involvement in the biosynthesis of primary and secondary metabolites including proteins,amino acids,nucleotides and volatile compounds,nitrogen is recognized as a vital nutrient for yeast growth and fermentation activity.Different nitrogen requirements compared toS.cerevisiaehave been identified for non-Saccharomycesyeasts.And therefore,it is necessary to understand such nutritional needs to guarantee a smooth fermentation.
2.1 Nitrogen influence on yeast growth and fermentation rates
As one of the crucial nutrients in grape must,nitrogen plays a key role in yeast growth and fermentation performance.Higher fermentation rates were generally observed in higher initial Yeast Assimilable Nitrogen (YAN) treatments regardless of yeast species,and yet the same didn’t necessarily apply to the yeast biomass production (Table 1).Higher amounts of aromatic amino acids irrespective of nitrogen level,other than YAN level,imposed more significant impacts on the maximal growth ofMetschnikowia pulcherrimaCECT 13130[4],indicating that certain groups of amino acids were the decisive factors in yeast growth instead of the total nitrogen contents.However,it should be acknowledged that increased fermentation rates corresponding to higher YAN levels in mixed fermentations may as well be due toS.cerevisiaein some cases.For instance,at a high YAN level,shortened fermentation duration and increased maximal fermentation rate were observed in mixed fermentation of a laboratory strainHanseniaspora guilliermondiiwithS.cerevisiaebut no such impact has been observed inH.guilliermondiisingle cultures[5].On the other hand,positive influences on fermentation rates were noted in both mixed and single fermentation ofM.pulcherrimaFlavia®[6].Both examples suggested that the impacts nitrogen imposed on fermentation kinetics and yeast growth are dependent on species/strain,and therefore it is crucial to understand nitrogen requirements including uptake preferences for non-Saccharomycesyeasts.
2.2 Nitrogen uptake preferences
Nitrogen sources present in grapes comprise complex compositions such as ammonium ions,peptides,proteins,and amino acids.Among these,yeasts have been proven to consume YAN sources at different orders,and uptake preferences were identified inS.cerevisiae,of which variations were observed among strains with genetic diversity[10].Yeast uptake preference of nitrogen sources are defined by 1) yeast consumption rate,fermentation kinetic,and yeast growth,2) yeast response based on Nitrogen Catabolite Repression (NCR) system,3) yeast response based on SPS(Ssy1p-Ptr3p-Ssy5p) system[2].On the other hand,yeast nitrogen preference is also studied based on yeast assimilation of nitrogen sources,and the ability of nitrogen to support yeast growth during fermentation.Investigations on yeast nitrogen uptake preference have been conducted in various non-Saccharomycesyeasts recently,by means of the first method mentioned,namely,the phenotypic characteristics.
Inter/intra-specific nitrogen uptake preference patterns were observed among the widely studied non-Saccharomycesyeasts(Table 2).Nevertheless,some nitrogen sources share similarities among those non-Saccharomycesstrains tested,which are glutamine,lysine,arginine,methionine,aspartate,glutamate,alanine,leucine,cysteine,and ammonium.Interestingly,bothS.cerevisiaeand the non-Saccharomycesyeasts were discovered to utilize some of the same amino acids as “preferred” nitrogen sources,which may further indicate genetic homology in terms of nitrogen uptake system in yeasts.On the other hand,differences were also noted.Proline,an amino acid inaccessible forS.cerevisiaeunder anaerobic conditions,was identified as a preferred nitrogen source forLachancea thermotoleransViniflora®ConcertoTM,Candida zemplininaCszB4,H.uvarumHuB10,S.bacillarisFC54,andS.bacillarisMUT5705[11-13].Different nitrogen uptake systems therein,compared toS.cerevisiae,though not yet revealed for most non-Saccharomycesyeasts,are proposed based on such findings.Furthermore,different nitrogen uptake preferences were observed for same strain,M.pulcherrimaFlavia®,andT.delbrueckiiBiodivaTM,in different experimental settings[11,14].Specifically,methionine and lysine were identified as preferred nitrogen sources forT.delbrueckiiBiodivaTM,and lysine and glutamine forM.pulcherrimaFlavia®in the presence ofin these two studies,and yet other different amino acids were observed as preferred nitrogen sources for the two strains (Table 2).Since the experimental design was slightly different,namely,media components,total nitrogen concentrations,and the proportion of organic/inorganic nitrogen,it is thus postulated that complexregulatory mechanisms are involved in the nitrogen uptake and utilization systems.And therefore,investigations of such mechanisms are of great significance.
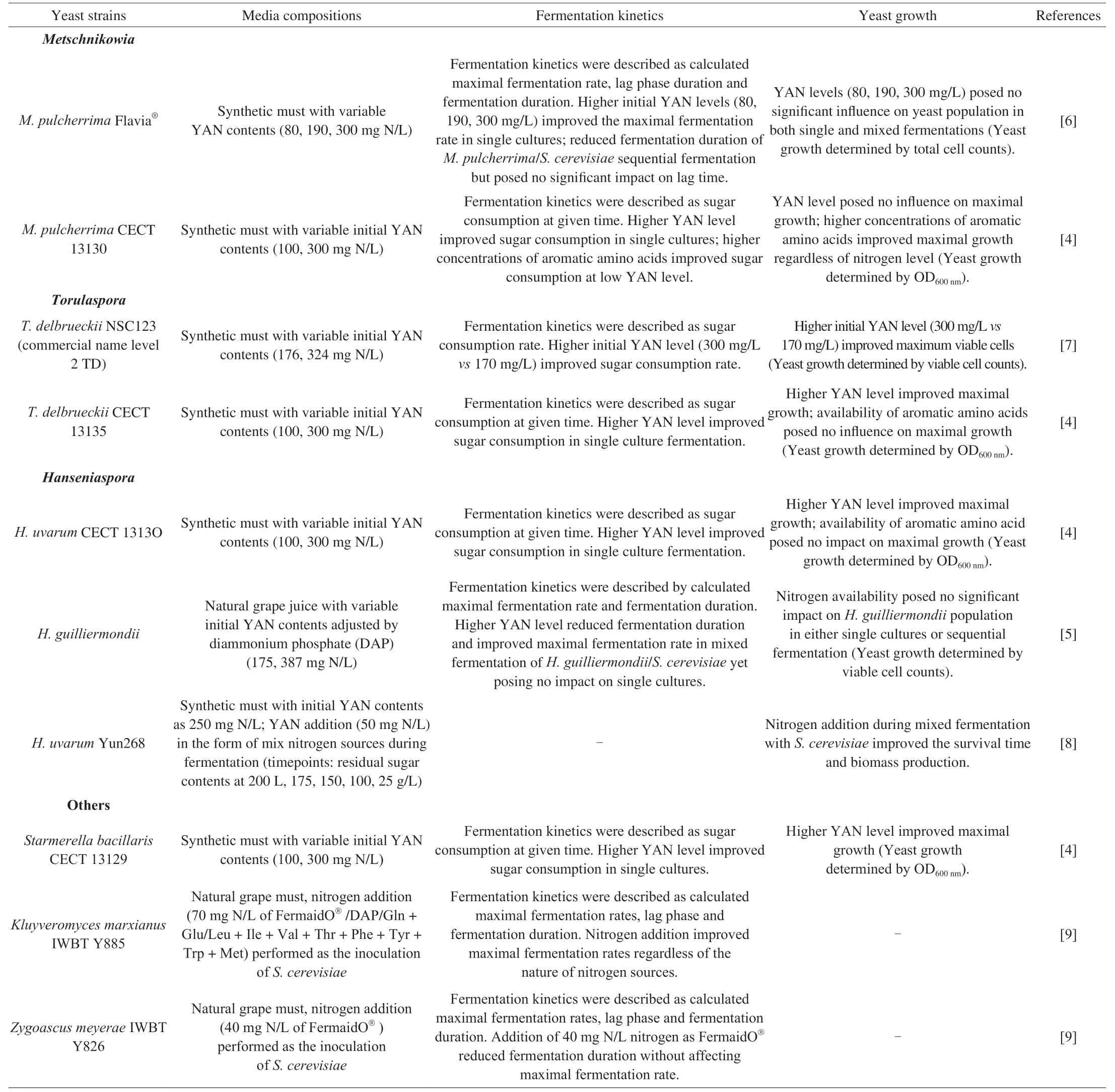
Table 1 Fermentation kinetics and yeast growth of non-Saccharomyces yeasts under different nitrogen conditions.
NCR mechanism,one of the regulatory systems,for amino acid permeases (AAPs) and ammonium permeases,plays a crucial part in nitrogen uptake preference.Under influence of such system,permeases such as GAP1,PUT4 (amino acid permease),MEP1,MEP2 and MEP3 (ammonium permease) are controlled leading to selective utilization of nitrogen sources by yeasts[15].This system has been studied thoroughly in many strains ofS.cerevisiae,elucidating complex mechanisms under nitrogen-deficient conditions[16-19].Such investigations are just launched in non-Saccharomycesyeasts.Lleixà et al.[20]investigated the NCR mechanisms inHanseniaspora vineaeand identified similar regulatory mechanism as that ofS.cerevisiaeQA23.Results indicated that GAP1 (general amino acid permease),MEP2 (ammonium permease)and PUT2 (delta-1-pyrroline-5-carboxylate dehydrogenase) showed evolving expression patterns from repression to activation throughout fermentation as nitrogen sources were gradually depleted,while AGP1 (high-affinity glutamine permease) was repressed as the amino acids were consumed.Similar gene expression patterns were reported inS.cerevisiae[17-18,21-22],suggesting the presence of NCR mechanism inH.vineae.

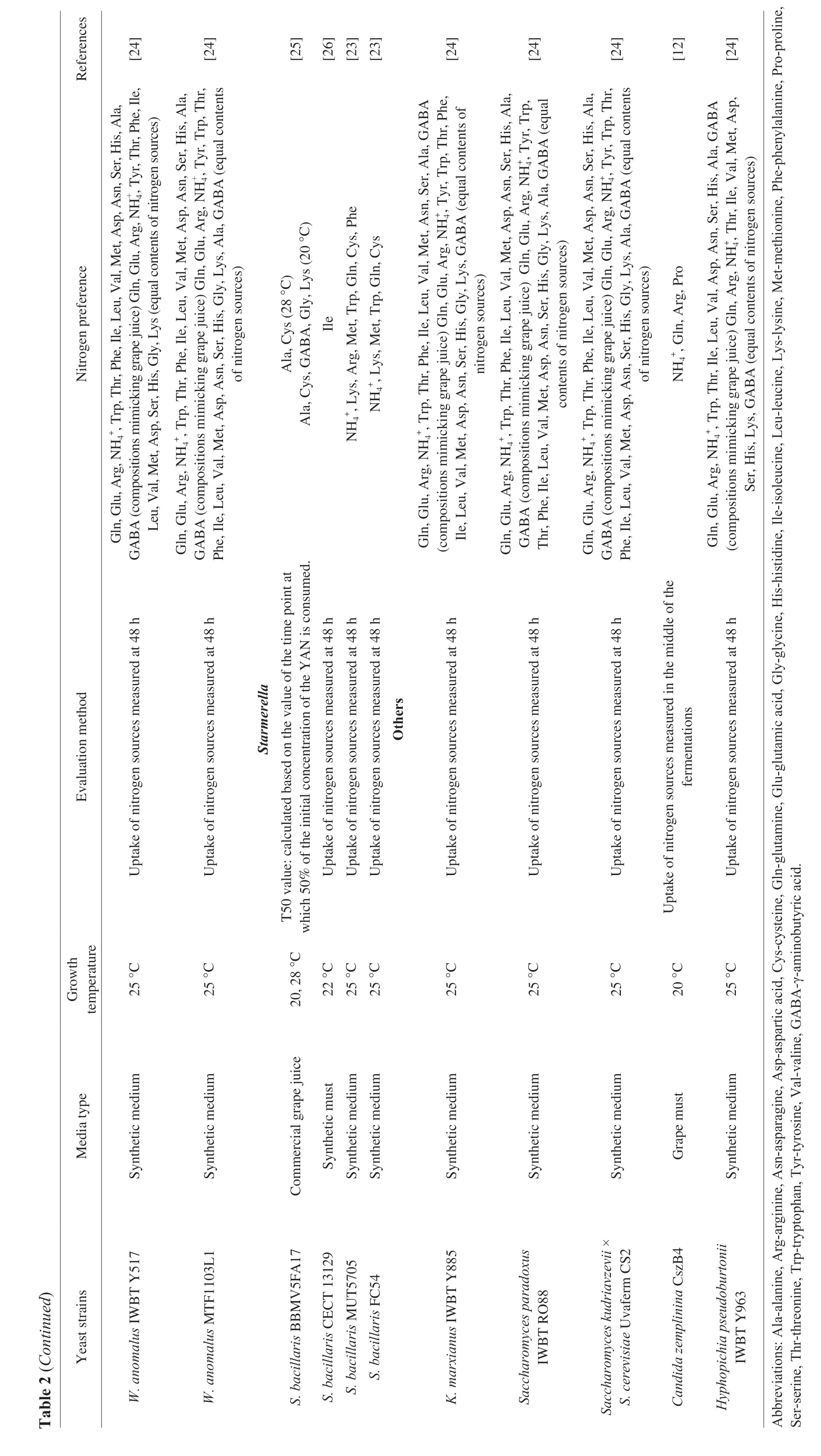
Ssy1p-Ptr3p-Ssy5 (SPS)-mediated system is another nitrogen uptake regulation mechanism identified in yeasts apart from NCR mechanism.The uptake of preferential amino acids is mediated by high-affinity permeases under the control of SPS system[2].Englezos et al.[23]discovered different phenotypes for nitrogen uptake ofS.bacillariscompared toS.cerevisiae,namely,the inability of amino acid uptake in the presence of ammonium.The authors therefore proposed 3 explanations,1) the lower efficiency of amino acid permeases mediated by SPS-system,2) the inhibitory effect of ammonium,3) the presence of another system for ammonium uptake.However,more work is encouraged to elucidate the molecular basis.
Given that nitrogen is one of the most vital nutrients as well as a limiting factor for bothS.cerevisiaeand non-Saccharomycesyeasts during fermentation,it is essential to understand the nitrogen requirements of both yeasts to ensure a smooth fermentation with the production of desirable metabolites.Therefore,further work along these lines is encouraged for a more comprehensive view of nitrogen requirements,uptake preferences and regulatory mechanisms of non-Saccharomycesyeasts.
3.Nitrogen influence on aroma production by non-Saccharomyces yeasts
3.1 Primary aroma production
Known as harboring enzymes responsible for releasing primary aroma compounds,non-Saccharomycesyeasts have long been studied on their volatile-enhancing enzymes,includingβ-glucosidase andβ-lyase.Both enzymes are critical for releasing varietal aromas from their non-odorous precursors that naturally occurred in grape must.
β-Glucosidase hydrolyzes the glycosidic bonds between glucosides and oligosaccharides and therefore releases the glucose glycone[27],namely,terpenes and norisoprenoids,as volatile compounds in wine.Enzyme activity has been widely studied under various winemaking conditions,such as various glucose concentrations,ethanol contents,and pH level,of which significant differences were observed for each factor[27].For example,high levels of glucose generally inhibit the activity ofβ-glucosidase for bothSaccharomycesand non-Saccharomycesyeasts.As another crucial nutrient factor present in grape must,nitrogen is rarely studied on its impact on eitherβ-glucosidase activity or corresponding volatile compound production during fermentation.Since nitrogen plays a vital role in both yeast growth and fermentation,enzyme activity might be influenced directly/indirectly during fermentation by influencing yeast metabolism.And therefore,future research along these lines is encouraged to explore the potential link between nitrogen status andβ-glucosidase activity and its corresponding aroma production.
β-Lyase,on the other hand,has been included in such investigation.With the hydrolytic ability on cysteinylated precursors,this enzyme is known to release varietal thiols,namely,4-mercapto-4-methylpentan-2-one (4MMP),3-mercaptohexanol (3MH),3-mercaptohexyl acetate (3MHA),from their non-odorous precursors.Yeasts,includingS.cerevisiaeand non-Saccharomycesyeasts,were reported with such ability of producing volatile thiols[28-30].Hence,the impact of alcoholic fermentation conditions on enzyme activity and the resulting volatile production is worth studying.Seguinot et al.[31]investigated the influence of initial nitrogen level on varietal thiol production as well asβ-lyase activity ofM.pulcherrimaFlavia®.Results indicated that varietal thiol production increased to different extents according to initial nitrogen levels.Release of 3MH and 4MMP increased up to 200 mg N/L of initial nitrogen level(821.3 ng/L for 3MH,326.2 ng/L for 4MMP),and then decreased with higher nitrogen concentrations,while production of 3MHA was positively correlated with initial nitrogen levels ranging 80-300 mg N/L.Impact onβ-lyase activity was also observed within different nitrogen levels.The highest enzyme activity was recorded throughout fermentation at a medium nitrogen level (190 mg N/L)in sequential fermentation ofM.pulcherrima/S.cerevisiae,whereas lower enzyme activity obtained for both low and high nitrogen levels(80,300 mg N/L).It is thus proposed that nitrogen availability in the grape must plays a key role in determiningβ-lyase activity and the resultant varietal thiol production under sequential inoculation ofM.pulcherrimaFlavia®/S.cerevisiae.
Even though nitrogen status has been proven to influence varietal thiol production by a non-Saccharomycesyeast strain[31],little data is yet available regarding the nitrogen impacts on primary aroma(varietal aroma) production,as well as the enzymatic mechanisms therein for other non-Saccharomycesyeasts.Known as a crucial contributor to the formation of complex aroma profiles during fermentation,such investigations on mechanisms and regulations of primary aroma production of non-Saccharomycesyeasts are in great need for further practical application.
3.2 Secondary aroma production
Secondary aroma,also known as fermentative aroma,is produced by yeasts and bacteria involved in fermentation process.Yeasts impose great contributions to wine aroma and flavor profile mainly during alcoholic fermentation.Such influences are generalized as the following mechanisms: 1) extraction of flavor and aroma compounds from grape musts with the alcohols produced,2) transformation of neutral compound from grapes to flavor/aroma-active compounds,3) direct production of flavor/aroma-active compounds as secondary metabolites,4) yeast autolysis[32].Among these,fermentation-derived aroma-active compounds,which are released viade novobiosynthesis and amino acid catabolism of yeasts (Fig.1),account for the largest proportion of the total volatile composition[33].Secondary aromas mainly comprise of acids,fusel alcohols and esters (Table 3),resulting from yeast metabolic activities during alcoholic fermentation,i.e.,catabolism of carbon and nitrogen sources(Fig.1).Production of such volatile compounds varies with yeast species and strains due to the variations of metabolic abilities within each yeast,and therefore imparting wine with different aroma profiles and styles.Non-Saccharomycesyeasts are known for versatile metabolic activities,thus rendering them a great tool to diversify aroma profiles of the wines produced.
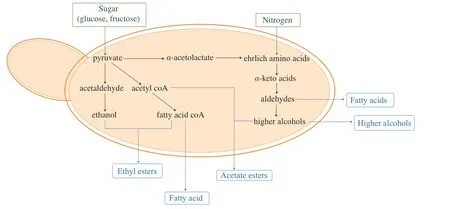
Fig.1 Metabolic pathway of major secondary aroma production.

Table 3 Major secondary aroma compounds and their corresponding amino acids (if available).
It is necessary to investigate nitrogen influences on secondary aroma production of non-Saccharomycesyeasts with the aim of further understanding the metabolic mechanisms therein for potential industrial applications.Several studies,though not in a significant amount,have been conducted to explore relationships between nitrogen contents in grape must and production of secondary aroma compounds.Interestingly,results indicating different correlations were observed (Fig.2).Since wine quality is highly dependent on volatile compositions,therefore the major secondary aroma groups,namely,volatile fatty acids,higher alcohvols,and esters,which received much attention,will be discussed in further details.
3.2.1 Volatile fatty acids
Volatile fatty acid,metabolites of glucose and amino acids,is mostly produced by yeasts during fermentation via biochemical process (Fig.1).Such compounds including isovaleric acid,isobutyric acid,octanoic acid can induce cheesy and buttery smell at certain levels,and deemed sweaty and rancid at excessive amounts.BothS.cerevisiaeand non-Saccharomycesyeasts have been identified with the ability to produce volatile fatty acids.
Complex impacts of nitrogen status were reported on the production of these compounds for non-Saccharomycesyeast strains.Among all the strains tested,despite the differences in experimental settings (i.e.,fermentation must,inoculation strategy),positive correlations were observed for most strains and volatile fatty acids,and yet negative impacts were also noted (Fig.2a).For instance,production of valeric acid is boosted for bothZ.meyeraeIWBT Y826 andK.marxianusIWBT Y885 with nitrogen addition during fermentation,while decreases were recorded for the production of hexanoic acid and decanoic acid by the latter strain with nitrogen addition[9].Complex results were obtained in the same study,in which different nitrogen supplements resulted in opposite effects on the production of volatile fatty acids.Specifically,both increase and decrease of octanoic acid and butyric acid were noticed in the addition of different nitrogen sources forK.marxianusIWBT Y885.The addition of inorganic nitrogen in the form of diammonium phosphate(DAP) improved the production of octanoic acid and butyric acid,while organic nitrogen (Fermaid O®) led to the opposite outcome.However,different inorganic nitrogen sources may lead to different results,which should be included as a further study for elucidation.As a complex nutritious product containing nutrients including nitrogen,vitamins and lipids,complex effects from Fermaid O®were observed,and yet the mechanisms therein remain unsolved.Therefore,further study,in which different types of nitrogen sources are used,together with the comprehensive investigations on yeast growth and metabolite production,are encouraged to elucidate the relationships between nitrogen and metabolic mechanisms of yeasts of interests.
3.2.2 Fusel alcohols
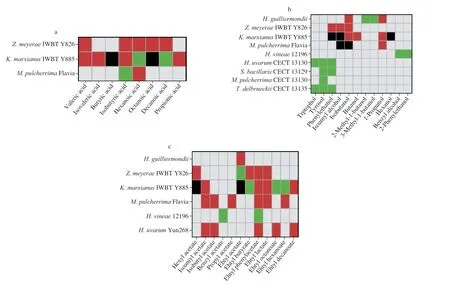
Fig.2.Impacts of nitrogen concentration on secondary aroma production of different non-Saccharomyces yeasts studied.(a) volatile fatty acid production;(b) fusel alcohol production;(c) ester production.Media types/nitrogen conditions/inoculation strategies: Z.meyerae IWBT Y826, K.marxianus KWBT Y885,Sauvignon blanc juice and Shiraz must/nitrogen additions during fermentation/sequential inoculation with S.cerevisiae;M.pulcherrima Flavia,synthetic medium/different initial nitrogen concentrations/sequential inoculation with S.cerevisiae;H.uvarum Yun268,synthetic medium/different initial nitrogen concentrations/simultaneous inoculation with S.cerevisiae;H.vineae 12196,chemically defined grape must/different initial nitrogen concentrations/single culture fermentation;H.uvarum CECT 1313O,S.bacillaris CECT 13129,M.pulcherrima CECT 13130,T.delbureckii CECT 13125,synthetic must/different initial nitrogen concentrations/single culture fermentation.Color interpretations: grey-no report yet;red-positive effect (higher volatile production in the presence of higher nitrogen levels);black-complex effect;green-negative effect (lower volatile production in the presence of higher nitrogen levels)[4,6,8-9,45].
Fusel alcohols are another group of yeast-derived volatile compounds contributing to aroma complexity of wine (Fig.1).Apart from major fusel alcohols such as 1-propanol,and isobutanol,there are some fusel alcohols present in moderate concentrations with high odor intensity,including 3-methyl-1-butanol,2-phenylethanol and isoamyl alcohol[35].Productions of fusel alcohols have been reported in bothS.cerevisiaeand non-Saccharomycesyeasts,with species/strains identified as the major determining factor on the level of production.Among these,either high or low producer of fusel alcohols was identified for non-Saccharomycesyeast strains,compared toS.cerevisiae,such asL.thermotolerans,H.uvarum,C.zemplinina,Saccharomycodes ludwigiibeing recognized as high producers,andCandida stellataandZygosaccharomyces fermentatias low producers[36].As a crucial metabolite of Ehrlich pathway,the production of fusel alcohols is largely dependent on the nitrogen contents and constitutions in the media,thereby making it necessary to investigate fusel alcohol production of non-Saccharomycesyeasts under different nitrogen conditions.Such studies are helpful in elucidating the correlations between potential nitrogen consumption and fusel alcohol production in non-Saccharomycesyeasts.
Up to now,9 non-Saccharomycesstrains have been reported for nitrogen impacts on fusel alcohol production.Specifically,the impacts of nitrogen concentrations,nitrogen constitutions were studied.Unlike those of volatile fatty acids,among which higher initial nitrogen concentration always results in an increased fusel alcohol concentration,large variations were observed for fusel alcohol production (Fig.2b).TakenM.pulcherrimaFlavia®as an example,while a positive impact of nitrogen level on propanol production was noted,quadratic effects on isobutanol and isoamyl alcohol were observed,with maximal production of isobutanol identified at 200 mg N/L.Complex effects similar to this were reported for other strains,such asK.marxianusIWBT Y885[9].Specifically,both positive and negative impacts on production of isoamyl alcohol,phenylethanol and hexanol were recorded depending on fermentation musts and natures of nitrogen sources added,where the addition of FermaidO®led to a decreased production of isoamyl alcohol in Sauvignon blanc fermentation,yet an opposite effect obtained in Shiraz must.
It is widely acknowledged that branched-chain higher alcohols are produced from amino acids via Ehrlich pathway,and therefore nitrogen level,especially amino acid uptake plays a key role in determining the release of these alcohols[36-37].Different impacts on the production of these compounds from nitrogen contents present in the must were encountered in such studies of non-Saccharomycesyeasts.Therefore,detailed investigations are yet to be conducted to fully explain the metabolic network of nitrogen for different non-Saccharomycesyeasts in terms of fusel alcohol production.
3.2.3 Esters
Esters,the most abundant compounds so far identified in wine[38],are formed throughout the winemaking process,including alcoholic fermentation,malolactic fermentation,and wine aging,among which yeast-derived esters are the most important contributing factors to wine aroma.Ethyl esters of fatty acids and acetate esters of fusel alcohols are the two major groups of esters present in wine,with the latter one being a more important aroma contributor.Of all the acetate esters,isobutyl acetate,isoamyl acetate and 2-phenylethyl acetate are the most crucial esters reminiscent of fruity,and flowery characters,with the alcohol precursors identified as isobutanol,isoamyl alcohol and 2-phenylethanol,metabolites of Ehrlich pathway.
Since similar esterification mechanisms were identified for most esters,nitrogen is therefore an important parameter influencing ester production.
Non-Saccharomycesyeasts,known to be good ester producers,have been applied widely for enhanced production of esters.Such species includeM.pulcherrima,H.guilliermondii,H.vineae,andHanseniaspora osmophila,which have been reported to produce pleasant fruity esters such as ethyl octanoate,2-phenylethyl acetate,and isoamyl acetate[29-43].On the other hand,however,some species are known to produce high levels of ethyl acetate,which can impart off aroma at a level of 150-200 mg/L[44].Despite esters being able to bring fruity and flowery characters to wine,such addition to aroma complexity can only be achieved at certain concentrations,and that excessive amounts (above 200 mg/L of total esters) are deemed defective[36].It is therefore critical to explore esterification mechanisms within non-Saccharomycesyeasts as major producers for more rational aroma management.Nitrogen is one of the important parameters to be investigated.
Based on the available studies reported,12 esters have been studied on their production by 7 non-Saccharomycesyeast strains under different nitrogen conditions.Positive correlations between nitrogen level and ester production were spotted,with negative and complex effects also recorded (Fig.2c).Among all the esters studied,productions of ethyl lactate,isoamyl acetate,ethyl decanoate,isobutyl acetate and propyl acetate are improved at higher YAN levels[8-9,31],whereas negative impacts were noted for ethyl phenylacetate and benzyl acetate inH.vineae12196,isolated from a Uruguayan vineyard[45].Species variations in relation to nitrogen impacts on the production of the same ester were also observed.Such an example is present for the production of ethyl acetate,of which production boost with DAP addition was reported for aH.guilliermondiistrain[5],and yet reduction with FermaidO®addition was identified forZ.meyeraeIWBT Y826[9].Furthermore,complex effects were observed inK.marxianusIWBT Y885[9].Specifically,additions of FermaidO®,glutamine+glutamate,and amino acids involved in Ehrlich pathway led to a decline in ethyl acetate release in both Sauvignon blanc and Shiraz musts,whereas the addition of DAP improved production only in Shiraz fermentation.A similar case was encountered for hexyl acetate production by this strain in the same must,where both DAP and glutamate+glutamine imposed a positive impact,yet FermaidO®led to the opposite.
Positive impacts of nitrogen levels were generally observed for acetate esters of fusel alcohols in most non-Saccharomycesyeast strains studied,with exceptions occurring to hexyl acetate and benzyl acetate regardless of the nature of the nitrogen source.Several enzymes responsible for ester formation have been characterized mainly inS.cerevisiae,such as the alcohol acetyl transferases I and II[46-48],and yet diversity in enzyme type and activity is hereby proposed for non-Saccharomycesyeasts based on various ester production abilities.Therefore,such studies in combination with nitrogen impacts with respect to metabolic mechanisms are greatly encouraged to better understand ester production in non-Saccharomycesyeasts.
3.2.4 Other volatile compounds
Apart from volatile fatty acids,fusel alcohols and esters,there are other yeast-derived aroma compounds formed during the fermentation process which play crucial roles in wine aroma complexity.And compounds like this include aldehydes,volatile phenols,and volatile sulfur compounds.
Aldehydes are important aroma compounds reminiscent of apples with low sensory thresholds.Acetaldehyde accounts for 90%of the total aldehyde detected in wine with concentrations ranging from 10 mg/L to 300 mg/L[44].Researchers reported that nitrogen addition in inorganic form (DAP) led to an increase in acetaldehyde production in mixed fermentation ofH.guilliermondii/S.cerevisiae,yet no impacts observed forH.guilliermondiisingle fermentation[5].A detailed investigation is encouraged to explore nitrogen impacts on both aldehyde production and yeast-yeast interactions during fermentations.
Volatile sulfur compounds present in wine,mainly contributing to negative aroma characteristics,such as onion,rubber,and cabbage,can sometimes bring positive attributes such as strawberry,grapefruit and passion fruit[49-51].Yeast metabolisms of sulfur-containing compounds,such as amino acids have been proven to be responsible for the release of these compounds[52].Such amino acids like methionine can be metabolized to produce several off aromas including methionol,4-methylthiobutanol,dimethylsulfide,S-methylthioacetate,3-methylthiopropyl acetate,and H2S.Metabolic mechanisms regarding the production of certain volatile sulfur compounds have been studied extensively inS.cerevisiae(i.e.,sulfate reduction sequence pathway for H2S production).Similar to other mechanisms,genetic regulation plays an important role in determining the amount produced,based on the fact that strain variations forS.cerevisiae,L.thermotolerans,andM.pulcherrima,were observed for H2S production under the same condition[3,53-54].
On the other hand,however,contributions of other fermentationrelated factors are also nonnegligible,among which,nitrogen has long been the top research interest.As a pilot study,Mendes-Ferreira and colleagues recorded no significant differences for the production of H2S under certain nitrogen sources (amino acids,DAP,and(NH4)2SO4) for several non-Saccharomycesstrains,while cysteine was provided as the sole nitrogen source supported the highest sulphide production,as agreed withS.cerevisiae[53].A few studies were conducted to investigate nitrogen influence on the production of other sulfur-containing volatile compounds (Table 4).The nature of nitrogen sources as additions triggered variations in production response.Researchers noticed positive correlations between the production of methionine-derived volatile compounds and Ehrlich pathway-related amino acids/FermaidO®,whereas decreases or unchanged in production were recorded for other types of nitrogen additions[9].Furthermore,fermentation must,potentially containing different key nutrition components also play a key role.The general decline in the release of sulfur-containing volatile compounds was observed for Shiraz musts,compared to the overall increases in Sauvignon blanc[9].More detailed investigations are required to elucidate the mechanisms therein.

Table 4 Nitrogen impacts of production of volatile sulfur compounds in non-Saccharomyces yeasts[5,9].
3.3 Metabolic pathways involved in secondary aroma production
Production of volatile compounds during fermentation has been proven to be related to the consumption of nitrogen sources,i.e.,via Ehrlich pathway.However,detailed metabolic pathways therein remain to be studied.In answering these questions,correlations between nitrogen consumption and volatile production,as well as the underlying mechanisms will be elucidated in non-Saccharomycesyeasts,which can in turn be of practical help to the industry.
Though some studies are available as pilot research to explore the impacts of nitrogen status on volatile production in non-Saccharomycesyeasts,little is known regarding nitrogen metabolism pathways,especially in relation to volatile compound production.Nitrogen uptake and consumption patterns were investigated by a few studies with hypotheses therefore made on the correlations with aroma production.Productions of aromatic alcohols,namely,tyrosol,phenylethanol,and tryptophol were studied inH.uvarumandM.pulcherrimain comparison withS.cerevisiae,in connection with nitrogen consumption as well as yeast growth[4].Such compounds were proposed as potential quorum-sensing molecules,since large amounts of production were observed at the early exponential stage,and secretion of these compounds into the medium was identified at the turning point of the exponential stage to the stationary stage during yeast growth[4].Meanwhile,results indicated that the formation of these potential quorum-sensing compounds was induced by nitrogenlimited conditions inH.uvarumandS.cerevisiae,in agreement with previous studies[55].Links between nitrogen uptake and the production of their respective volatile compounds were also studied.It was proposed that higher nitrogen uptake did not necessarily result in higher production of fermentative aroma compounds,based on the fact that as a good producer of fusel alcohols,Z.meyeraeIWBT Y826 consumed only a few amounts of the corresponding amino acids[24].
Furthermore,a few detailed studies aiming to map the nitrogen metabolic pathway associated with volatile compound production have been conducted,which also supported the hypothesis by Rollero et al.[24]raised.Su and colleagues[14]have identified metabolism of valine and leucine inT.delbrueckiiandM.pulcherrimavia15N-and13C-isotope labelling approach.Different patterns were observed.Specifically,forT.delbrueckii,a larger proportion of amino acids consumed was used forde novosynthesis of proteinogenic amino acids instead of the production of volatile compounds.On the other hand,however,forM.pulcherrima,more amino acids consumed were directed into Ehrlich pathway for the synthesis of fusel alcohols,in line with the observation of lower biomass production.AK.marxianusstrain IWBT Y885 was investigated with a similar approach,using an isotopic enrichment study coupled with gene expression analysis to demonstrate the molecular basis[24].Results indicated that the metabolic flux of which exogenous valine and leucine were catabolized to form isobutanol and isoamyl alcohol,was much lower than that ofS.cerevisiae.Furthermore,expression levels of the genesLEU1/2/4/9,responsible for the production ofα-isocaproate,the precursor of isoamyl alcohol,were also lower than those ofS.cerevisiae.However,higher productions of isovaleric and isobutyric acid were observed inK.marxianus,whereas similar levels of isotopic enrichment were recorded for both strains,indicating a larger proportion ofde novosynthesis via the CCM (Central Carbon Metabolism) pathway.Upregulation of genesALD2/3/4/5/6,responsible for the formation of fusel acids from aldehyde was detected inS.cerevisiae,while PDR12 involved in the acid transport was highly expressed inK.marxianus.On the other hand,the metabolic flux of phenylalanine to form phenylethanol and phenylethyl acetate was diverse forK.marxianuscompared withS.cerevisiae.Higher levels of flux through Ehrlich pathway to produce both compounds were noted forK.marxianus,in accordance with the higher expression levels of the genes involved in the catabolism of aromatic amino acids into the formation ofα-ketoacids(ARO8/9/10) as well as the formation of fusel alcohol from aldehyde (ADH1/2/3/4/5/6andSFA1).Such data indicated the great contribution of exogenous phenylalanine to produce phenylethanol and the corresponding acetate ester in this strain.These studies discussed above not only provides insight into nitrogen metabolism of non-Saccharomycesyeasts,but is of industrial interest as well,in terms of nitrogen management in practical winemaking.
4.Future perspective and conclusion
Nitrogen,naturally present in grapes,is one of the most crucial nutrients during the winemaking process due to its involvement in yeast metabolic activity.It is thus important to elucidate nitrogen metabolism in yeasts,and such mechanisms have been demonstrated extensively inS.cerevisiae.Non-Saccharomycesyeasts have gradually gained research attention due to their versatile catalytic activities,especially the ability to add complexity to wine aroma profiles.Therefore,the influences of nitrogen impacts on non-Saccharomycesyeasts in terms of their fermentation performances and aroma productions are worthy of exploration,which is the major topic of this review.
In this review,nitrogen impacts on yeast growth during fermentation are first discussed.Among the studies reviewed,higher fermentation rates were generally observed at higher initial YAN levels,with exceptions present for some strains.Furthermore,different impacts were also noted in single/mixed fermentation,indicating a complex effect from yeast-yeast interactions.Inter/Intra-specific variations were also identified,and the same is true for YAN preference studies.Despite that most strains studied share a few nitrogen sources as their “preferred” sources,such as glutamine,lysine,arginine,methionine,aspartate,glutamate,alanine,leucine,and ammonium,distinctiveness was also spotted for each strain.NCR mechanism was determined in aH.uvarumstrain,as an attempt to figure out the mechanism behind these phenotypic characteristics.However,more work along this line is required.Since nitrogen metabolism is in tight relation with secondary aroma production,such impacts of non-Saccharomycesyeasts are hence reviewed due to their important roles as crucial tools for aroma improvement during winemaking.The influences of nitrogen status on both primary and secondary aroma formation and their potential mechanisms are discussed based on current studies.Complex effects were obtained,suggesting the hypothetical involvements of many other oenological parameters.More thorough studies focusing on metabolic mechanisms are in great need to decipher the complications.
Future studies with systematic investigations on nitrogen metabolisms in non-Saccharomycesyeasts are highly encouraged.Such study will incorporate multiple factors (such as sugar,lipids,vitamins) as well asS.cerevisiae/non-Saccharomycesinteractions,for a better understanding on nitrogen metabolisms to enable a more efficient nutrient management during fermentation.Furthermore,research on the correlations between nitrogen metabolic pathway and volatile compound production will provide insights into the aroma production of wine.Studies of this category will also be helpful in discovering rational nitrogen management strategies in winemaking for desirable aroma production.
Conflicts of interest
Baoguo Sun is an editorial board member forFood Science and Human Wellnessand was not involved in the editorial review or the decision to publish this article.All authors declare that there are no competing interests.
Acknowledgment
This work was supported by grants from the National Natural Science Foundation of China (32172340).

- 食品科学与人类健康(英文)的其它文章
- Protective effects of oleic acid and polyphenols in extra virgin olive oil on cardiovascular diseases
- Diet and physical activity inf luence the composition of gut microbiota,benef it on Alzheimer’s disease
- Ganoderma lucidum: a comprehensive review of phytochemistry,eff icacy,safety and clinical study
- Resveratrol combats chronic diseases through enhancing mitochondrial quality
- Demonstration of safety characteristics and effects on gut microbiota of Lactobacillus gasseri HMV18
- Cyanidin-3-glucoside protects the photooxidative damage of retinal pigment epithelium cells by regulating sphingolipid signaling and inhibiting MAPK pathway