
Mechanism of action of cordycepin in the treatment of hepatocellular carcinoma via regulation of the Hippo signaling pathway
2024-02-15 07:59XiaominLiQingLiuSongyuXieXiaopingWuJunshengFu
食品科学与人类健康(英文) 2024年2期
Xiaomin Li,Qing Liu,Songyu Xie,Xiaoping Wu,Junsheng Fu
College of Life Sciences, Mycological Research Center, Fujian Agriculture and Forestry University, Fuzhou 350002, China
Keywords: Cordycepin Hepatocellular carcinoma Hippo signaling pathway GBP3 ETV5
ABSTRACT Hepatocellular carcinoma (HCC) is one of the common most malignant tumors.This study aimed to determine the in vitro and in vivo anticancer activity of cordycepin and elucidate its mechanism of action.The results of in vitro and in vivo studies revealed that cordycepin inhibited proliferation and migration in HepG-2 cells and inhibited the growth of HepG-2 xenograft-bearing nude mice by inducing apoptosis.Transcriptome sequencing analysis revealed a total of 403 differential genes,which revealed that cordycepin may play an anti-HCC role by regulating Hippo signaling pathway.The regulatory effects of cordycepin on the Hippo signaling pathway was further investigated using a YAP1 inhibitor.The results demonstrated that cordycepin upregulated the expression of MST1 and LAST1,and subsequently inhibited YAP1,which activated the Hippo signaling pathway.This in turn downregulated the expression of GBP3 and ETV5,and subsequently inhibited cell proliferation and migration.Additionally,YAP1 regulated the expression of Bax and Bcl-2,regulated the mitochondrial apoptotic pathway,and induced apoptosis by upregulating the expression of the caspase-3 protein.In summary,this study reveals that cordycepin exerts its anti-hepatocarcinoma effect through regulating Hippo signaling pathway,and GBP3 and ETV5 may be potential therapeutic targets for hepatocarcinoma.
1.Introduction
Hepatocellular carcinoma (HCC) is a malignant tumor of liver tissues that is characterized by a high recurrence rate and aggressiveness[1].The two major issues in the treatment of HCC are the difficulty of early diagnosis and poor prognosis,and the lack of specific therapeutic targets[2].Cancers of the liver primarily include HCC,intrahepatic cholangiocarcinoma,mixed hepatocellular cholangiocarcinoma,and fibrous lamellar HCC.The current clinical treatments for HCC include surgery,chemotherapy,and immunotherapy,among others.Surgical resection is the main treatment modality for patients with HCC,but the rate of recurrence is high after surgery.Therefore,the identif ication of highly effective yet less toxic antitumor drugs for HCC and elucidation of their mechanism of action are of utmost importance.
More and more studies have shown that natural products are the main source of bioactive substances,especially those with low toxicity and few side effects.CordycepsmilitarisLink is a fungus that is consumed as food and medicine,and has been considered to have natural medicinal properties,including anti-angiogenic,antitumor,anti-inflammatory,and hypoglycemic activities[3].Cordycepin is a potent active substance extracted fromC.militarisand has been reported to have various biological activities,including anticancer,antiviral,and immunomodulatory properties[4].Although the cytotoxic effects of cordycepin on HepG-2 cells have been demonstrated,the specif ic mechanism of action has not been fully elucidated,and there is a scarcity of studies on the mechanism by which cordycepin inhibits HepG-2 cells via the Hippo signaling pathway.The Hippo signaling pathway regulates cell survival,proliferation,and differentiation,and inhibits tumor growth.Therefore,abnormalities in the Hippo signaling pathway may lead to abnormal tumor growth[5].The core of the Hippo signaling pathway comprises a series of proteins,including MST1/2,SAV1,MOB1,and LAST1/2[6].When Hippo signaling is inactivated,unphosphorylated YAP enters the nucleus and binds to the transcriptional enhancement binding domain (TEAD) of transcription factors,which promotes cell proliferation and inhibits apoptosis[7].The anticancer effects of the YAP inhibitor,verteporfin(VP),are mediated via the inhibition of the YAP-TEAD complex,and VP has been used in the treatment of HCC[8].The role of the Hippo signaling pathway in the anti-HCC effects of cordycepin remains to be elucidated.
RNA sequencing (RNA-seq) is a technique for sequencing and analyzing the entirety or a portion of the mRNAs,small RNAs,and non-coding RNAs in cells and tissues using highthroughput sequencing technologies[9].In recent years,transcriptome sequencing has been widely used in several studies for mining novel genes,annotating gene function,and determining differential gene expression.Transcriptome sequencing can also be used as an auxiliary means to explore the potential mechanism of action of anticancer drugs.In this study,transcriptome analysis showed that cordycepin may play an anti-liver cancer role by regulating Hippo signaling pathway.Using a transcriptome sequencing technology and bioinformatics tools,this study aimed to identify the differentially expressed genes (DEGs) in HCC before and after cordycepin treatment.We additionally performed gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses for screening the potential key genes related to the Hippo signaling pathway and determined the alterations in gene expression for obtaining insights into the possible mechanism underlying the inhibitory effect of cordycepin on HCC.
2.Materials and methods
2.1 Experimental materials and reagents
The HepG-2 human HCC cell line was purchased from iCell Bioscience Inc.RPMI 1640 culture medium and fetal bovine serum(FBS) were procured from Gibco,and cordycepin was purchased from Sigma.Penicillin-Streptomycin Liquid,5% BSA (Blocking Buffer),and 0.1% aqueous solution of crystal violet were purchased from Beijing Solarbio Science &Technology Co.,Ltd.Xylene,anhydrous ethanol,30% H2O2,and other chemicals were purchased from Sinopharm Chemical Reagent Co.,Ltd.VP was procured from Beijing Bioss Biotechnology Co.,Ltd.,while the monoclonal antibodies against YAP1,caspase-3,Bcl-2,MST1,GBP3,ETV5,TGF-β1 and E-cadherin were purchased from Proteintech Group,Inc.FITC-labeled goat anti-secondary antibody was purchased from ABClonal Biotechnology Co.,Ltd.
Animals: BALB/C nude mice,weighing 14-15 g,3-5 weeks old,male,SPF class,were purchased from Wu Lab Animals Online (http://www.wssydw.com/m/index.asp).
2.2 Cell culture
HepG-2 cells were recovered from liquid nitrogen,inoculated in RPMI 1640 medium supplemented with 10% FBS and 1% Penicillin-Streptomycin Liquid,placed in a 5% CO2incubator at 37°C,and allowed to grow to a confluence of 80%-90%.The cells were digested with trypsin for culture passage.
2.3 Cell viability assay
Cells at the logarithmic stage of growth were selected,diluted to a concentration of 5 × 104cells/well,and inoculated in 96-well plates.After the cells had grown to a confluence of 80%-90%,the control,cordycepin-treated,and zeroing wells were set up separately.The control group contained the complete medium with cells,the cordycepin group contained cordycepin at concentration gradients of 25,50,100,200,and 400 µmol/L,and the zeroing wells were devoid of cells.A total of 5 replicate wells were set up for each group and at least 3 biological replicates were performed.After 48 h of treatment,the medium was discarded and 10 µL of 5 mg/mL MTT and 90 µL of complete medium were added to each well,incubated for 4 h,and discarded.Int formazan were then dissolved in DMSO,shaken well,and placed in an enzyme marker for measuring the absorbance at 490 nm (OD490nm) and determining cell viability.Cell survival was determined using the following formula:
All the results obtained from the MTT assay were confirmed by at least 3 independent replicates and 5 replicate wells for each group.
2.4 Colony formation assay
The cells at the logarithmic phase of growth were inoculated with 500 cells/well in 6-well plates,and treated with cordycepin for 7-14 days.Colony formation was observed daily under a microscope,and the culture was ceased when the number of clones was more than 50.The culture medium was discarded,following which the cells were fixed with anhydrous methanol and subsequently stained with 0.1% crystal violet solution.The cells were then rinsed with PBS until no obvious purple background was visible,and finally photographed and counted using ImageJ software.
2.5 Wound healing assay
The cells were diluted to a concentration of 1 × 105cells/well and seeded in 6-well plates.After the cells reached a confluence of 80%-90%,the cultures in the 6-well plates were scratched with a 10 µL spear head,washed with PBS for removing the cells that had dropped,and subsequently treated with cordycepin.Scratch healing was recorded at 0 h and 48 h for each group and analyzed using ImageJ.
2.6 Hoechst 33 258 staining
Clean coverslips were taken and placed in a 6-well plate.The coverslips were washed with 75% ethanol and PBS,following which HepG-2 cells were inoculated and cultured in the 6-well plate,and the culture was terminated after 48 h of cordycepin treatment.The cells were fixed and stained according to the instructions in the Hoechst 33258 kit,and subsequently sealed and observed under a fluorescence microscope for determining cellular apoptosis.
2.7 RNA extraction and cDNA library construction
HepG-2 cells were inoculated in RPMI 1640 medium containing 10% FBS and 1% Penicillin-Streptomycin Liquid,incubated in a 5%CO2incubator at 37 °C,and allowed to grow to a confluence of 90%or higher for passaging.The cells were inoculated in a 6-well plate at a concentration of 1 × 105cells/well.The control and cordycepintreated groups were set up when the cells reached a confluence of 80%-90% after attaching to the wall,and three replicate wells were set up for each group.The replicate wells included Con-1,Con-2,and Con-3 in the control group,and Cor-1,Cor-2,and Cor-3 in the cordycepin-treated group.After 24 h of treatment,the cells were washed twice with PBS and collected.
RNA-seq transcriptome sequencing was performed according to the method described by Zhou et al.[10],with slight modifications.The total RNA from cordycepin-treated HepG-2 cells was extracted using TRlzol lysate,and a Nanodrop,Agilent 2100 Bioanalyzer was used for analyzing the purity,concentration,and integrity of the RNA.The polyA tails of eukaryotic mRNAs were combined using magnetic beads with Oligo,following which the mRNAs were fragmented into short fragments using a fragmentation buffer,and used as templates for the generation of double-stranded cDNA.End repair,addition of polyA tails,and ligation of sequencing junctions were performed for the construction of double-stranded cDNA,followed by fragment size selection using AMPure XP beads.The final cDNA library was enriched by PCR experiments.The cDNA libraries were then added to each lane of the flow-through tank and sequenced using an Illumina NovaSeq sequencer.The quality score was determined using the relevant software.For library construction,1 µg of RNA from each sample was used,and three biological replicates were performed for each group.
2.8 RNA quantification and qualification
The concentration and purity of the extracted RNA were measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific,Wilmington,DE,USA).The integrity of the RNA was assessed using an RNA Nano 6000 Assay Kit of an Agilent 2100 Bioanalyzer system(Agilent Technologies,CA,USA).
The raw data (raw reads) in FASTQ format were first processed using in-house Perl scripts.In this step,clean data (clean reads) were obtained by removing the reads containing adapter sequences,reads containing ploy-N,and low quality reads from raw data.At the same time,the Q20 and Q30 scores,GC content,and sequence duplication level of the clean data were calculated.The downstream analyses were performed using clean data of high quality.The correlation of the gene expression levels between the samples was determined using Pearson’s correlation coefficient.
2.9 DEG screening and GO and KEGG enrichment analyses of DEGs
In order to identify the DEGs with altered expression levels in the cordycepin-treated group relative to those of the control group,differential expression analysis of the two groups was performed using the EBSeq package in R.The resulting false discovery rate (FDR)was adjusted using the posterior probability of being differentially expressed (PPDE).FDR < 0.05 and |log2foldchange (FC)| ≥ 1 were selected as the thresholds for determining significantly differential expression.
GO enrichment analysis of the DEGs was performed using the cluster Profiler package in R.Enrichment analysis is based on hypergeometric testing for determining the GO entries that are significantly enriched in comparison to the entire genome background.Gene Set Enrichment Analysis (GSEA) can also be performed using the cluster Profiler package in R.The GO terms related to the biological process category withP< 0.01 were arranged and represented in a bar chart according to the size of thePvalue.The terms related to cell proliferation,migration,and apoptosis were indicated with different colors.
The KEGG database by Kanehisa et al.[11]enables the exploration of the high-level functions and utilities of biological systems,including cells,organisms,and ecosystems,from molecular information,especially large-scale molecular datasets generated by genome sequencing and other high-throughput experimental technologies (http://www.genome.jp/kegg/).The differential genes screened from the transcriptome were mapped to the genes related to the Hippo signaling pathway,and the intersecting genes were subjected to heat map clustering and volcano plot analyses.
2.10 Effect of VP on HepG-2 cells
YAP1 is a key downstream effector molecule of the Hippo signaling pathway.In order to investigate whether cordycepin inhibits HCC through the Hippo signaling pathway,HepG-2 cells were treated with VP,a YAP inhibitor,and the control group (CK),cordycepin group (COR),verteporfin group (VP),and pre-treated with VP followed by cordycepin incubation 48 h (COR+VP)groups.The expression of important genes of the Hippo signaling pathway and apoptosis-related genes were detected by fluorescencebased quantitative PCR following treatment with cordycepin and VP(VP+COR),and the expression of important proteins was detected by Western blot.
The MTT method was used for detecting the activity of VP on HepG-2 cells and for observing the changes in cellular morphology in the different treatment groups.
2.11 Detection of the expression of genes related to the Hippo signaling pathway and apoptosis-related genes by q-PCR
Cells from different treatment groups were collected,and RNA was extracted using the Trizol method.The purity and concentration of the RNA was tested,and reverse transcription was performed using a TransScript All-in-one First-Strand cDNA Synthesis SuperMix for qPCR (One-step g DNA Removal) kit.The concentration of the sample was diluted to 500 ng/µL for single-strand cDNA synthesis,and the cDNA was stored at -20 °C as backup.The primer sequences(Table 1) were synthesized by Yihe Biotechnology Co.,Ltd.and the cDNA was diluted 5 times before quantification.The following reaction conditions were used for qPCR: pre-denaturation at 94 °C for 2.5 min,denaturation at 94 °C for 5 s,annealing at 58 °C for 30 s,and extension at 60 °C for 5 s,for a total of 40 cycles.The expression of the genes of the Hippo signaling pathway and apoptosis-related genes was detected,and three biological replicates of each sample were performed for relative quantification according to the 2-△△CTmethod.

Table 1 Primer sequence of Q-PCR.
2.12 Immunofluorescence assay
Clean coverslips were placed in 6-well plates,and cells were seeded in 6-well plates.After incubation with cordycepin for 0,24,48 h,the cells were fixed with 4% paraformaldehyde,permeabilized with 0.1% Triton X-100 for 15 min,washed with PBS for 3 times,and blocked with PBS containing 1% BSA,cells were incubated with YAP1 primary antibody (1:400) at 4 °C overnight.FITClabeled goat anti-mouse secondary antibody (1:200) was added and incubated for 1 h,then the nuclei were stained with DAPI,and finally the anti-fluorescence tablets were placed in an inverted Fluorescence microscope for observation and photography.
2.13 Detection of the expression of YAP and caspase-3 proteins with western blotting
The cells were collected and the total cellular protein was extracted by adding RIPA lysate.Some of the proteins in the supernatant were selected for determining the protein concentration using a BCA protein concentration assay kit.Each well was sampled with 40 µg of protein,following which the proteins were separated by SDS-polyacrylamide gel electrophoresis (PAGE) and transferred to polyvinylidene difluoride (PVDF) membrane.The membrane was blocked with 5% BSA for 1 h at room temperature,and subsequently incubated overnight with the specific primary antibody at 4 °C.The membrane was then incubated with horseradish peroxidase (HRP)-labeled goat anti-mouse or goat anti-rabbit secondary antibody at room temperature.The membrane was finally placed in an ECL solution for allowing the reaction to occur and color development.
2.14 Xenograft studies
HepG-2 cells at logarithmic growth stage were taken at a concentration of 1 × 107cells/mL,and the cells were mixed with matrigel at 1:1.HepG-2 xenograft-bearing mice model was established by subcutaneous inoculation with HepG-2 cells into the skin of nude mice.After the xenograft tumors grew to about 100 mm3,they were randomly divided into control and cordycepin groups.The control group was given sterile water by gavage every day,and the cordycepin group (COR group) was given 20 mg/kg cordycepin by gavage every day,and the administration was stopped when the xenograft tumor grew to about 1 000 mm3.The mice were execution,the tumors were excised,weighed,and then some tissues were fixed with 4% paraformaldehyde and paraffin-embedded for further immunohistochemical analysis.
2.15 Immunohistochemistry studies
Sections of tumor tissue treated with or without cordycepin were prepared and left at room temperature for 60 min,dewaxed with xylenes I,II,and III,rehydrated with gradient ethanol,washed with PBS,cupped with 3% H2O2for blocking endogenous peroxidases,boiled for antigen repair,closed,incubated overnight with the primary antibody,and subsequently incubated with the secondary antibody.SABC was added to the tissue sections in a dropwise manner,following which DAB working solution was added for staining.Following hematoxylin staining,differentiation,blue return,dehydration,blocking,and other steps,the tissues sections were finally observed under a microscope and photographed.The expression of YAP1,caspase-3,and Bcl-2 in the tumor tissues of the untreated and cordycepin-treated groups was measured.
2.16 Statistical analysis
The data are presented as the mean ± standard deviation (SD).One-way ANOVA was performed for analyzing the variance in this study using SPSS 25.0.P< 0.05 was considered to be statistically significant andP< 0.01 was considered to be highly statistically significant.
3.Results
3.1 Cordycepin inhibits the proliferation of HepG-2 cells
The cytotoxicity of cordycepin on HepG-2 cells following treatment with cordycepin for 48 h at different doses is depicted in Fig.1B.The results demonstrated that the cell survival rate gradually decreased in the dose range 25-400 µmol/L,and cordycepin had a dose-dependent effect on the survival rate of HepG-2 cells(Fig.1B).Based on the cytotoxicity curve,the IC50value of cordycepin on HepG-2 cells was determined to be 149.892 µmol/L.The concentration of cordycepin selected in subsequent studies was 150 µmol/L.To investigate the effect of cordycepin on the proliferation ability of HepG-2 cells,we performed colony formation assay (Fig.1A).The results demonstrated that cordycepin significantly inhibited the proliferation of HepG-2 cells at concentrations 50-400 µmol/L.The clone formation rates were (52.497 ± 1.990)%and (1.325 ± 0.177)% when the concentrations of cordycepin were 200 and 400 µmol/L,respectively (Fig.1C).Altogether,the results demonstrated that cordycepin had cytotoxic effects on HepG-2 cells and significantly inhibited cell proliferation.
3.2 Cordycepin induces apoptosis in HepG-2 cells
Hoechst 33258 is a convenient and popular nuclear counterstain that emits blue fluorescence under fluorescence microscopy when bound to DNA.In this study,Hoechst 33258 was used for studying the effect of cordycepin on the apoptosis of HepG-2 cells following treatment at different concentrations.As depicted in Figs.2A and B,the number of apoptotic cells following treatment with cordycepin was significantly higher than that of the control group.The nuclei of the stained apoptotic cells showed the presence of dense,brightly fluorescent chromatin,whereas the nuclei of normal cells appeared oval or round and emitted a uniform blue fluorescence.In apoptosisrelated proteins,cordycepin up-regulated the expression of Bax and caspase-3 and down-regulated the expression of Bcl-2 protein in a dose-dependent manner (Figs.2C and D).The results demonstrated that cordycepin promoted apoptosis in HepG-2 cells in a concentration gradient-dependent manner.
3.3 Cordycepin inhibits the migration of HepG-2 cells

Fig.1 Cordycepin inhibits proliferation in HepG-2 cells.(A) Colony formation assay to detect the effect of cordycepin on the proliferation of HepG-2 cells.(B) The MTT assay was used for determining the effect of cordycepin on cell viability,and it was observed that cordycepin significantly inhibited the viability of HepG-2 cells.(C) The plate cloning results were analyzed using ImageJ software and showed that the number of cell clones of HepG-2 decreased with increasing concentration.**P < 0.01 vs control group.
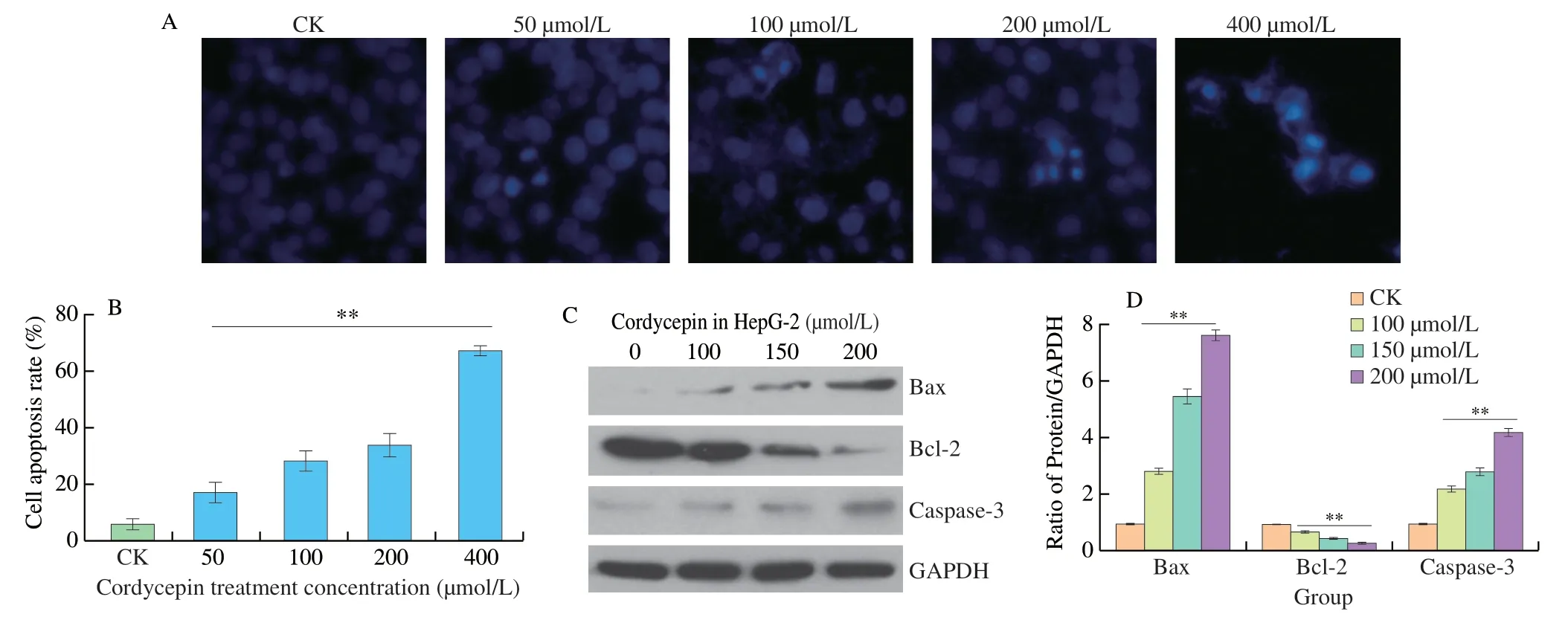
Fig.2 Cordycepin induces apoptosis in HepG-2 cells.(A) The effect of cordycepin on apoptosis of HepG-2 cells was detected by Hoechst 33258 dye.(B) Hoechst 33258 dye could permeate the cell membrane and release blue fluorescence from the embedded double-stranded DNA.The membrane permeability of apoptotic cells was enhanced and dense blue fluorescence was visible in the nuclei of apoptotic cells,and the number of apoptotic cells increased with increasing concentration.(C) Western blot detected the expression of Bax,Bcl-2 and caspase-3 proteins in HepG-2 cells treated with different concentrations of cordycepin for 48 h.(D) The relative expression levels of Bax,Bcl-2,and caspase-3 proteins were analyzed by optical density.Cordycepin can upregulate the expression levels of Bax and caspase-3 proteins and downward the expression levels of Bcl-2 protein.**P < 0.01 vs.control group.
HCC have potent invasive migration ability,and we tested the effect of cordycepin on the migration of HepG-2 cells using the scratch healing assay.The results of the scratch healing assay showed that the migration of HepG-2 cells was inhibited in all the treated groups,compared to that of the control group,and the differences between the scratched areas of the treated and control groups were highly significant (P< 0.01) (Fig.3).The migration rate of HepG-2 cells in the control group was 2.64-fold higher than that of the group treated with 200 µmol/L cordycepin.The findings revealed that cordycepin could inhibit the motility of HepG-2 cells and significantly inhibited cell migration ability.
3.4 Reliability of transcriptome sequencing data
Transcriptome sequencing was performed using two cell samples in the control and treatment groups,with three biological replicates per group.After transcriptome sequencing of the samples,the clean reads obtained from the samples were aligned to the designated reference genome for comparison,and the alignment efficiency reached 97.02%-97.70%.The matching efficiency is the most direct measure of transcriptome data utilization,and the results revealed that the selected reference genome assembly met the requirements of information analysis.Analyses of the base composition and quality revealed that the percentage of Q30 bases was 94.98% or higher,and the GC content of each sample ranged from 50.18% to 52.04%(Table 2,indicating that the quality of transcriptome sequencing of the control and cordycepin-treated HepG-2 cells was good.A total of 22 328 genes were detected by analyzing the data obtained by transcriptome sequencing,of which 2 375 were determined to be novel genes.
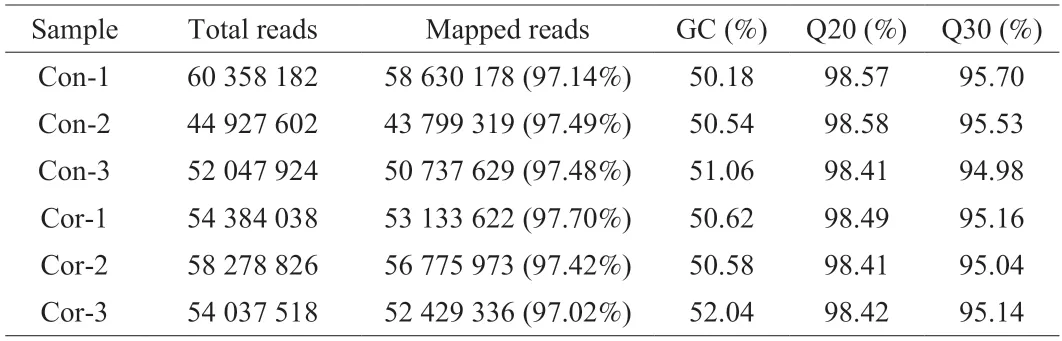
Table 2 Quality control analysis of sequencing data.
3.5 Identification and functional enrichment analysis of DEGs
The differences in gene expression between the control and cordycepin-treated groups were determined.A total of 403 differential genes were determined by analysis using screening conditions of FDR < 0.05 and |log2FC| > 1,of which 228 genes were upregulated and 175 were downregulated in the cordycepin-treated group,compared to the gene expression profile of the control group (Fig.4A).
GO enrichment based functional analysis of the 403 DEGs revealed that the DEGs were enriched in GO terms classified into biological processes,cellular components,and analytical functions.A total of 53 GO biological processes were analyzed usingP< 0.01 as the screening condition,and the bar graphs were arranged according to the lg(P-value) values.The biological processes associated with apoptosis,cell proliferation,and cell migration were labeled with different colors,where red was associated with apoptosis,blue indicated cell proliferation,and black represented cell migration(Fig.4B).The results of GO analyses were consistent with those of our previous study,indicating that cordycepin can regulate the processes of apoptosis,proliferation,and migration in HepG-2 cells.

Fig.3 Cordycepin inhibits the motility of HepG-2 cells.A sterile pipette tip was used for making a straight scratch,simulating a wound.Cells were recorded by microscopy at 0 h and after 48 h of cordycepin incubation.Wound healing assay indicated that cordycepin can suppressed HepG-2 cell migration.**P < 0.01 vs.control group.
3.6 Identification of DEGs involved in the Hippo signaling pathway
KEGG pathway enrichment analysis of the DEGs was separately performed using the clusterProfiler package in R,and 19 KEGG pathways were identified usingP< 0.01 as the threshold.The DEGs were mainly enriched in the Hippo signaling pathway,Steroid biosynthesis,Transcriptional misregulation in cancer,TNF signaling pathway,and other signaling pathways (Fig.4C).Of these,the Hippo signaling pathway had the smallestP-value,indicating that this pathway may play an important role in the anti-HCC effect of cordycepin.Enrichment analysis of all the differential genes related to the Hippo signaling pathway led to the identification of a total of 46 intersecting genes.TheYAP1gene is an important gene of the Hippo signaling pathway.In order to investigate the interactions between the 46 intersecting genes andYAP1,a protein-protein interaction(PPI) network map was constructed using Cytoscape.As depicted in Fig.5A,the results demonstrated thatYAP1and the 46 intersecting genes closely interacted with one other.Details of the 46 genes are in Appendix 1.
Hierarchical clustering analysis was performed based on the expression levels of the intersecting genes among the DEGs that were related to the Hippo signaling pathway in the control and cordycepin-treated groups (Fig.4D).A change in color from green to red during cluster analyses indicated increased gene expression.Heat map analysis revealed that 26 genes were downregulated while 20 genes were upregulated among the intersecting DEGs that were related to the Hippo signaling pathway in the control and cordycepintreated groups.The 3 samples in the control and cordycepin-treated groups showed a similar clustering pattern,in which genes with the same function or closely related genes formed distinct gene clusters.The control and cordycepin-treated groups had one gene cluster with high expression and one gene cluster with low expression.The transcription of some genes in the control and cordycepin-treated groups was markedly reversed,such as the transformation of the low expression similar cluster in the control group into high expression clusters in the cordycepin-treated group.
3.7 Clinical significance of key genes in the Hippo signaling pathway
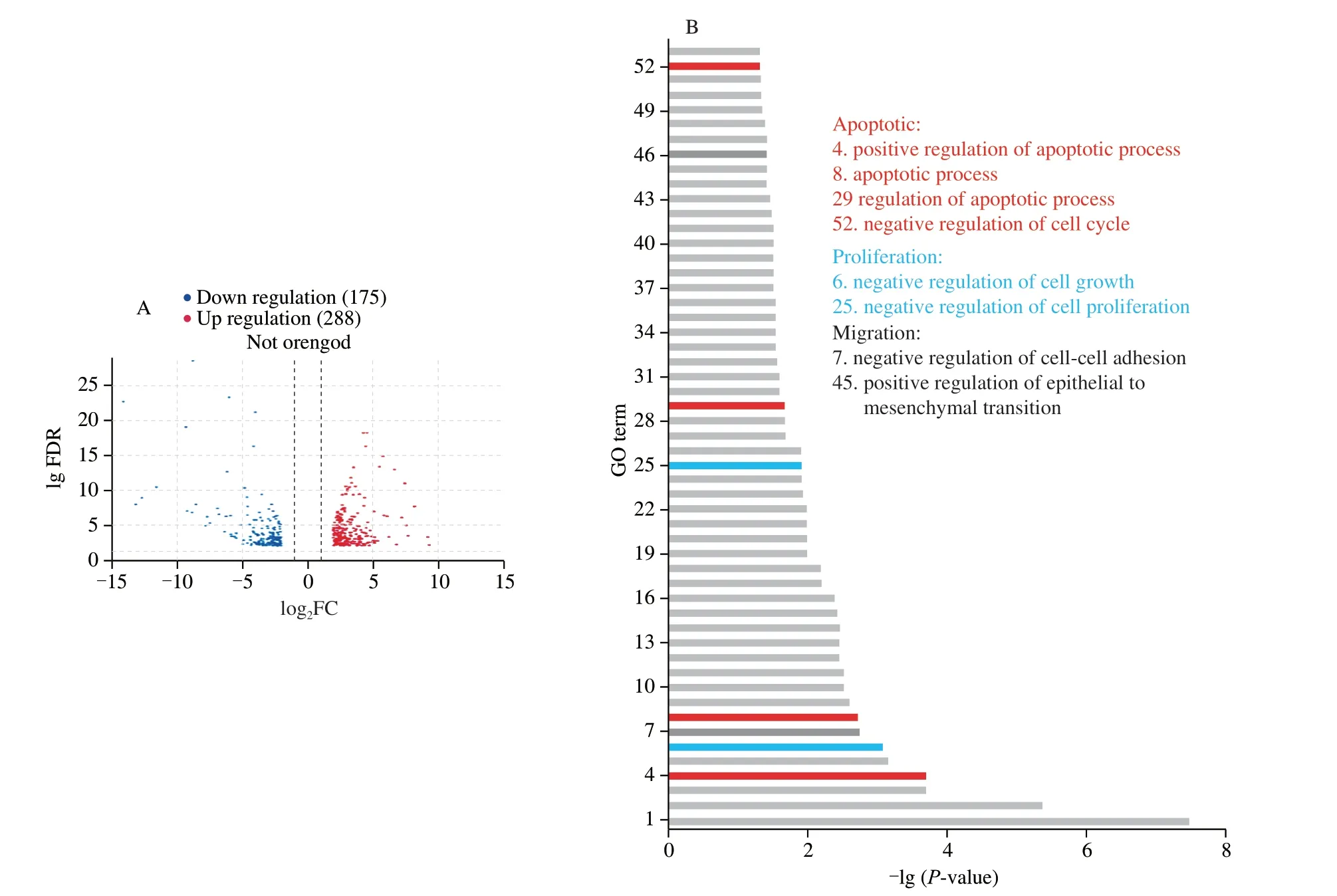
Fig.4 Elucidation of the role of the Hippo signaling pathway in the anti-HCC effect of cordycepin using transcriptomics analyses.(A) Differential genes between the control and cordycepin-treated groups,and volcano plot analysis of the DEGs.The upregulated and downregulated genes are indicated in red and blue colors,respectively.(B) GO enrichment analysis of the differential genes;biological processes with P < 0.01 were ranked by the size of the P-value and plotted as bars.Biological processes associated with apoptosis,proliferation,and migration were labeled in red,blue,and black,respectively.(C) KEGG enrichment analysis of the differential genes.The screened pathways with P < 0.01 were graphically represented using bubble plots,which reveal that the differential genes are enriched in the Hippo signaling pathway.The color represents -lg(P-value) and the size of the bubbles indicate the number of enriched genes.(D) The intersection genes of differential genes and Hippo signaling pathway-related genes were taken,and a total of 46 genes were obtained and heat map clustering analysis was performed.
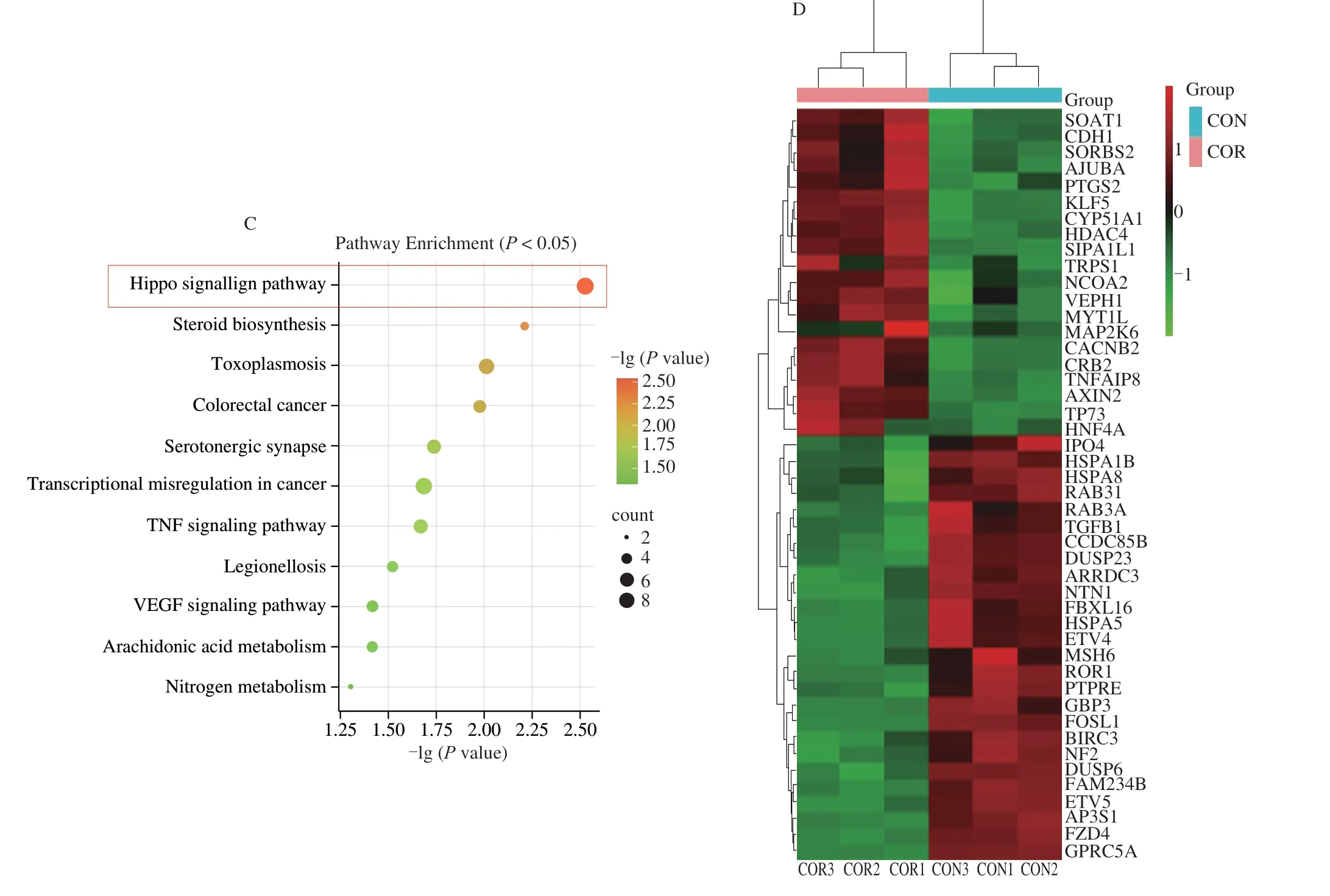
Fig.4 (Continued)
In order to further investigate the clinical significance of the genes related to the Hippo signaling pathway,we analyzed the expression ofYAP1,MST1,andGBP3in normal and liver cancer tissues using data retrieved from the GEPIA 2 database.The results demonstrated thatYAP1is highly expressed in liver cancer tissues,while the expression ofMST1is low in liver cancer tissues and significantly different from that of normal liver tissues.Compared to that of normal liver tissues,the expression ofGBP3was high in tumor tissues,and differed significantly from that of normal tissues (Fig.5B).We mapped the intersecting DEGs related to the Hippo signaling pathway to a volcano map,and identified thatETV5,GBP3,GPRC5A,andFOSL1had high fold changes in expression (Fig.5D).The results suggested that these genes may play an important role in the anti-HCC effect of cordycepin.We further investigated the correlation between the expression ofMST1andETV5in the Kaplan-Meier plotter database and the overall survival (OS) of patients with HCC.The results demonstrated that patients with HCC having high expression ofMST1had significantly longer OS than patients with HCC having low expression ofMST1.Additionally,patients with HCC having low expression ofETV5had better prognosis (Fig.5C).The correlation among the expression ofGBP3,ETV5,GPRC5A,FOSL1,andYAP1was analyzed by Pearson correlation coefficient analysis (Fig.5E),and the graph showed that the expression ofGBP3andETV5was more positively correlated to the expression ofYAP1,indicating thatGBP3andETV5could be important effector genes in the regulation of the Hippo signaling pathway by cordycepin.
3.8 Cordycepin inhibited YAP1 transcription and expression of Hippo signaling pathway components in HepG-2 cells
The transcriptome analysis indicated that YAP1 may play an important role in the anti-hepatocarcinoma effect of cordycepin.We further investigated the effects of cordycepin on YAP1 and Hippo signaling pathways.We found that cordycepin time-dependently(Figs.6A and B) and dose-dependently (Figs.6C and D) inhibited the expression of transcription factor YAP1 in HepG-2 cells.Furthermore,the time-dependent translocation of YAP1 to the nucleus of HepG-2 cells was inhibited by cordycepin (Fig.6E),which indicated that cordycepin could inhibit the transcriptional activity of YAP1 in HepG-2 cells.To further verify the accuracy of the transcriptome,we used western blot assay to detect the expression of some intersecting gene-related proteins between differential genes and Hippo signaling pathway (Fig.6F).As the concentration of cordycepin increased,cordycepin significantly up-regulated the expression of MST1,E-cadherin,and other proteins;and down-regulated the expression levels of TGF-β1,GBP3,ETV5 in a dose-dependent manner(Fig.6G).These results suggest that cordycepin affects the translocation and transcriptional activity of YAP1 and regulates the expression of Hippo signaling pathway-related proteins.
3.9 Inhibition of YAP1 enhances the inhibitory effect of cordycepin on HepG-2 cell viability
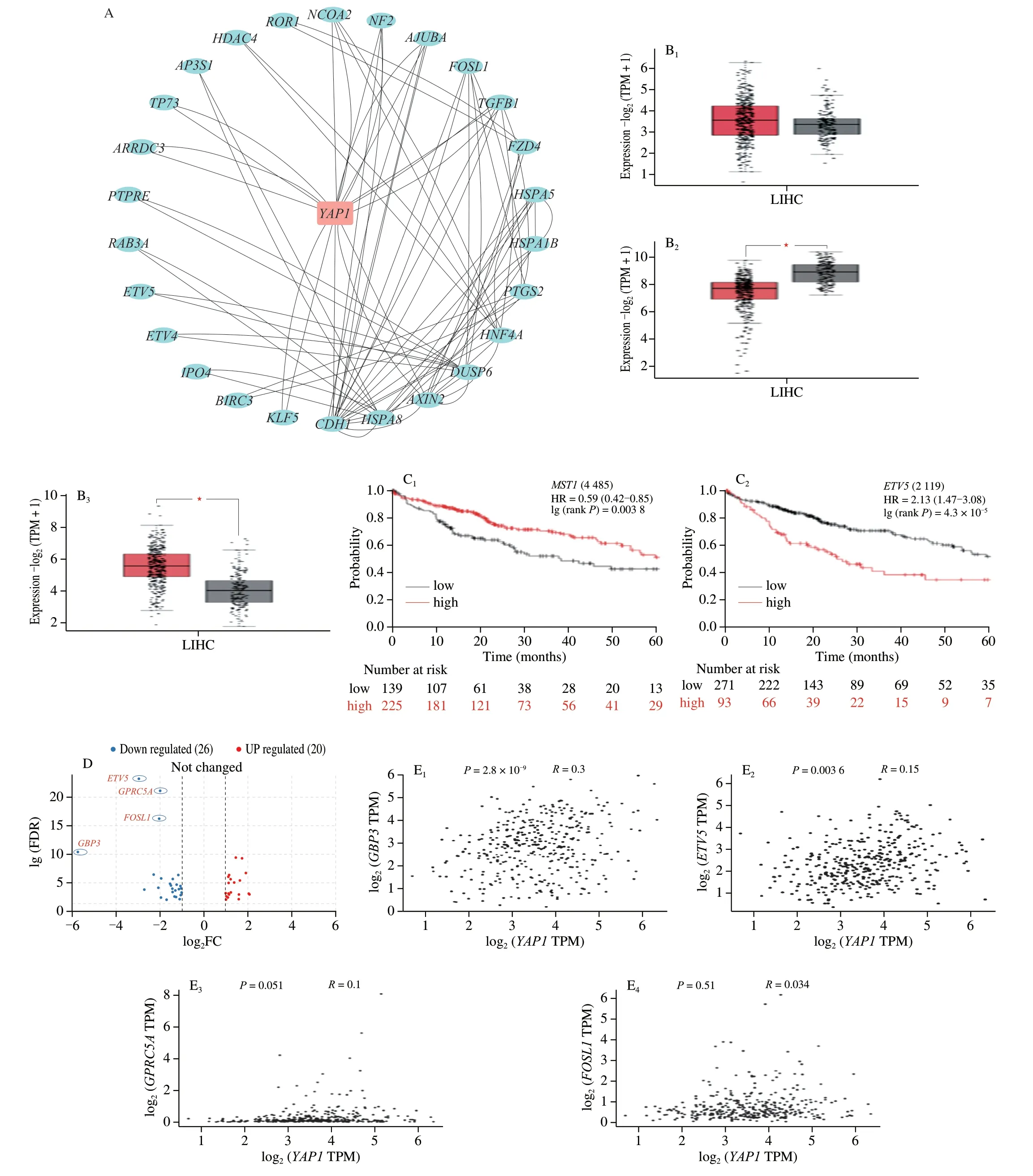
Fig.5 Key gene correlation analysis of cordycepin against hepatocellular carcinoma.(A) The interactions between YAP1 gene and 46 intersecting genes were analyzed in String database,and the PPI interactions network was plotted by Cytoscape software,from which it can be seen that YAP1 and intersecting genes closely interacted with each other.(B) The expression of key genes of Hippo signaling pathway in normal liver tissues and hepatocellular carcinoma tissues were analyzed in GEPIA 2 database (B1-B3,YAP1,MST1,GBP3).(C) Prognostic analysis of MST1 and ETV5 in patients with liver cancer in Kaplan-Meier plotter database.(D) Volcano plotting of 46 intersecting genes,GBP3,ETV5,GPRC5A and FOSL1 may be important genes in cordycepin regulation of Hippo signaling pathway against hepatocellular carcinoma.(E) Correlation of GBP3,ETV5,GPRC5A,FOSL1 and YAP1 was analyzed in the GEPIA 2 database.
Since YAP1 is the core component of the Hippo signaling pathway,and we used the YAP inhibitor VP to investigate the mechanism of action of the Hippo signaling pathway in cordycepin against liver cancer.The effect of VP on the proliferation of HepG-2 cells was detected using the MTT assay,and the results are depicted in Fig.7C.VP inhibited the proliferation of HepG-2 cells in a concentration gradient-dependent manner.The cell survival rate in the group of COR+VP was lower than that following treatment with cordycepin alone (Figs.7A,D).
3.10 Regulation of the Hippo signaling pathway in HepG-2 cells by cordycepin
In order to investigate whether the Hippo signaling pathway plays a key role in the inhibitory effect of cordycepin on HepG-2 cells,VP was used to detect the changes of cordycepin mediated Hippo signaling pathway related genes and protein expression by pretreatment with or without VP.As depicted in Fig.7H,the expression of YAP1 was lower in the group pre-treated with VP than that of the group treated with cordycepin alone.The expression levels ofMST1,LAST1,GBP3andETV5mRNAs were subsequently examined,and the results are provided in Fig.7F.Compared to those of the control group,the cordycepin-treated group increased the mRNA expression levels ofMST1andLAST1,the expression ofLAST1mRNA was significantly upregulated in the COR +VP group compared to that of the group treated with cordycepin alone.Treatment with cordycepin or VP alone downregulated the expression ofGBP3andETV5mRNAs;however,the COR+VP group highly significantly downregulated the expression ofGBP3andETV5mRNAs (P< 0.01),compared to that of the group treated with cordycepin alone.In conclusion,the results demonstrated that the anticancer effects of cordycepin are mediated via regulation of the expression of genes related to the Hippo signaling pathway which leads to the inhibition of cell proliferation and migration.
3.11 Activation of the Hippo signaling pathway enhances cordycepin-induced apoptosis
In order to determine whether cordycepin-induced apoptosis in HepG-2 cells was associated with the Hippo signaling pathway,the YAP inhibitor (VP),was used in the study for investigating whether pretreatment with VP affected cordycepin-induced apoptosis in HepG-2 cells.The expression of apoptosis-related genes and proteins was subsequently detected,and the results are depicted in Fig.7G.The findings revealed that compared with those of the group treated with cordycepin alone,COR+VP group significantly upregulated the expression levels ofCASP3,Bax,andCytcmRNAs,and significantly decreased the expression levels ofBcl-2andPARP1mRNAs.The morphological changes in the cellular nuclei were detected using Hoechst 33258 dye,and the results revealed that the number of nuclei that stained bright blue increased after VP pretreatment followed by cordycepin (Figs.7B,E),indicating that the nuclei of HepG-2 cells exhibited chromatin condensation and nuclear rupture,and cordycepin could promote VP-induced apoptosis.These results indicated that cordycepin induced apoptosis by activating the Hippo signaling pathway in HepG-2 cells.

Fig.7 Effects of cordycepin on the expression of genes related to the Hippo signaling pathway,and the effect of the Hippo signaling pathway on cordycepininduced apoptosis.(A) Alterations in the morphology of HepG-2 cells in the control,COR (cordycepin treated),VP (VP treated),and pre-treated with VP followed by cordycepin incubation for another 48 h (COR+VP) groups.Cordycepin enhanced the activity of VP (10 × 20).(B and E) Hoechst 33258 dye was used to detect the effect of different treatment groups on the apoptosis of HepG-2 cells (10 × 20).(C) Effects on the survival rate of HepG-2 cells treated with different concentrations of verteporfin,which inhibited the viability of HepG-2 cells.(D) Cell survival of HepG-2 cells in control,COR,verteporfin,and COR+VP,respectively.(F) Expression analysis of important genes of Hippo signaling pathway in different treatment groups using qPCR.(G) The expression of key genes of mitochondrial apoptosis pathway was detected by qPCR.(H) Expression of YAP1 and caspase-3 proteins in different treatment groups.*P < 0.05;**P < 0.01 vs.control group;#P < 0.05,##P < 0.01 vs.COR group;@P < 0.05,@@P < 0.01 vs.VP group.
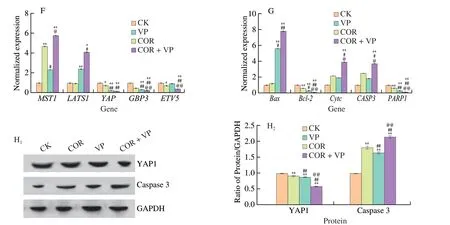
Fig.7 (Continued)
3.12 Cordycepin inhibits the growth of HepG-2 xenografts in Balb/c nude mice
In order to investigate the clinical significance of cordycepin in HCC,we further investigated its antitumor effect on mice bearing HepG-2 tumors.Mice were intraperitoneally injected with 20 mg/kg cordycepin every day for 16 days.The tumor size was observed on alternate days,and the administration of cordycepin was terminated when the xenograft tumor grew to a volume of 1 000 cm3.The animals were sacrificed and the tumors were photographed and weighed.The results demonstrated that tumor volumes in the cordycepin-treated groups were highly significantly reduced compared to that of the control group (P< 0.01) (Fig.8A).In conclusion,cordycepin can inhibit the growth of HepG-2 xenografts in nude mice.
3.13 Cordycepin inhibits tumor growth and induces apoptosis by activating the Hippo signaling pathway and caspase-3
We next assessed the expression of YAP1,Bcl-2,and caspase-3 in the tumor tissues by immunohistochemical staining.As depicted in Fig.8,the results demonstrated that the positive expression rates of YAP1 and Bcl-2 in the tumor tissues of the cordycepin-treated group were lower than those of the control group,and the positive expression rate of caspase-3 was higher than that of the control group (Fig.8E).The results indicated that cordycepin upregulated the expression of caspase-3 and downregulated the expression of YAP1 and Bcl-2 in tumor tissues (Figs.8B-D).The findings also suggested that cordycepin may induce apoptosis in HCC xenograft tumors by regulating the Hippo signaling pathway,thereby inhibiting their growth.
4.Discussion
Liver cancer is a common malignancy with high recurrence and mortality rates.As chemotherapy usually affects both normal cells and tumor cells,treatment with natural anticancer drugs can reduce the damage to normal cells.Cordyceps millitarisis a mushroom that is used as food and medicine and is a rich source of several active constituents,including cordycepin.The anticancer effects of cordycepin have been widely reported,and the molecular mechanism underlying the antitumor effect of cordycepin is complex.This study aimed to investigate the anticancer activity of cordycepin on HepG-2 cells,and perform in-depth investigations into the mechanism underlying the effect of cordycepin against liver cancer by transcriptome sequencing.

Fig.8 Cordycepin regulates the Hippo signaling pathway for inhibiting the growth of HepG-2 xenograft tumors.(A) Cordycepin can significantly inhibit the volume of xenograft tumors in nude mice.(B-D) The results found that there were more caspase-3 protein-positive cells and fewer YAP1 and Bcl-2 protein-positive cells in the cordycepin group than in the control group,there were significant differences between the two groups (P < 0.01).(E) Immunohistochemical detection of YAP1,Bcl-2 and caspase-3 expression in cordycepin-treated tumor tissues and control tumor tissues (10 × 40).**P < 0.01 vs.control group.
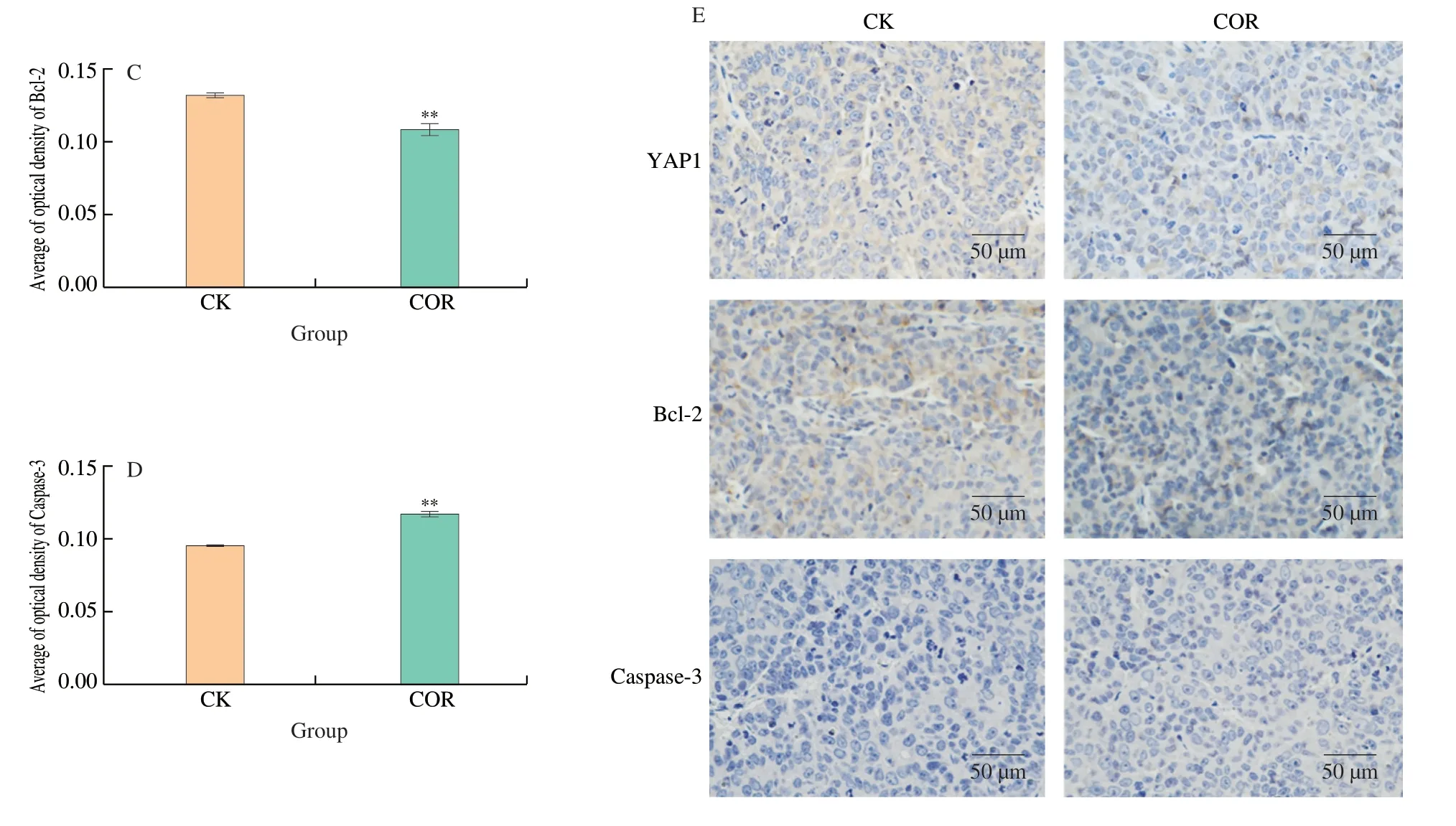
Fig.8 (Continued)
Invitrostudies have confirmed the cytotoxic and anticancer effects of cordycepin on HepG-2 cells.In this study,cordycepin reduced the cell survival of HepG-2 in a dose-dependent manner.The effects of cordycepin on the migration,proliferative ability,and apoptosis of HepG-2 cells were detected by the wound healing assay,colony formation assay,and Hoechst 33258 staining,respectively.The results demonstrated that cordycepin inhibited the motility and proliferation of HepG-2 cells and induced apoptosis in a concentration-dependent manner,which confirmed the anti-HCC effect of cordycepin.
We performed transcriptome sequencing analysis for further investigation of the molecular mechanism underlying the effect of cordycepin against HCC.A total of 403 differential genes were identified by transcriptome sequencing analysis,of which the expression of 228 DEGs was upregulated and that of 175 DEGs was downregulated.The results indicated that these DEGs may be involved in biological processes such as proliferation and migration of malignant cells,which are inhibited by cordycepin in HCC.The KEGG database provided information regarding the advanced functions of organisms,relationship between genes and cordycepin,and related metabolic pathways,which helped analyze the possible mechanism of action of cordycepin against cancer.In this study,KEGG enrichment analysis revealed that the HCC cells treated with cordycepin were mainly enriched in the Hippo signaling pathway.We further verified the correlation between the inhibitory effect of cordycepin on liver cancer and the Hippo signaling pathway.The genes related to the Hippo signaling pathway were screened for identifying the intersection genes with differential expression,and a PPI network of the intersection genes of DEG-Hippo signaling pathway was constructed using the online String database.We further screened four genes that may play an important role in the Hippo signaling pathway,namely,GBP3,ETV5,GPRC5AandFOSL1.The Kaplan Meier-plotter and GEPIA databases were used for analyzing the relevance of these important genes in the clinical characteristics and prognosis of patients with HCC.The results demonstrated that the expression ofYAP1,MST1,GBP3andETV5correlated with the prognosis of patients with HCC.
The Hippo signaling pathway is primarily involved in the regulation of cell proliferation and programmed cell death.VP,an inhibitor of YAP,was used in this study for investigating the role of the Hippo signaling pathway in the inhibitory effect of cordycepin against HCC.YAP/TAZ is the main effector molecule that acts downstream of the Hippo signaling pathway,and upregulation of the expression of YAP increases the proliferative ability of HCC[12].The expression ofMST1was upregulated following treatment with cordycepin,and was significantly different from that of the control group (P< 0.05).It has been shown that MST1/MST2 functions by suppressing theYAP1protein by binding to Ser127,which inhibits the progression of HCC[13].Last1 is a member of the Dbf2-associated nuclear protein kinase family,which together with Mst1 constitutes a part of the Hippo signaling pathway that inhibits excessive cell proliferation.Deletion of the Last1 gene leads to increased cell proliferation and migration[14].Xu et al.reported that guanosinebinding proteins (GBPs) play a key role in cellular autoimmunity,and GBP3,which is highly expressed in gliomas,promotes the proliferation of glioma cells[15-16].Identification of genes related to the Hippo signaling pathway revealed that the COR+VP group significantly downregulated the expression levels ofYAP1,GBP3,andETV5mRNAs and upregulated the expression levels ofMST1andLAST1mRNAs,compared to those of the group treated with cordycepin alone.The results indicated that cordycepin may inhibit the proliferative and migration ability of HepG-2 cells through the Hippo signaling pathway.Meng et al.reported that ETV5 can enhance cell proliferation,migration,and EMT processes through the PIK3CA signaling pathway,while treatment with cordycepin downregulates the expression of ETV5,possibly via the Hippo signaling pathway,and subsequently inhibits cell proliferation and migration[17].
Apoptosis is an important mechanism of cell death and cordycepin induces apoptosis in HepG-2 cells;however,the precise mechanism underlying the effect of cordycepin remains to be elucidated.Bax and Bcl-2 are important factors in the mitochondrial apoptotic pathway,and the main regulators of endogenous apoptosis.Bax has a pro-apoptotic effect while Bcl-2 has an anti-apoptotic effect,and caspase-3 is a key inducer of apoptosis[18-20].The intracellular ratios of Bax and Bcl-2 are normally relatively stable;however,the Bax/Bcl-2 ratio in HCC cells may undergo alterations following treatment with cordycepin.In order to determine whether the apoptosis induced by cordycepin in HepG-2 cells is related to the Hippo signaling pathway,we examined the expression of apoptosis-related genes with or without pretreatment with VP,and the effect of the presence or absence of inhibitors on the apoptosis of HepG-2 cell was determined by Hoechst 33258 staining.The results revealed that cordycepin upregulated the expression levels ofCASP3,Bax,andCytcgenes,downregulated the expression ofBcl-2andPARP1mRNAs,and increased the Bax/Bcl-2 ratio,thus promoting apoptosis.HepG-2 cells were pre-treated with VP followed by cordycepin incubation for another 48 h (COR+VP) significantly regulated the expression of apoptosis-related genes.The proteins in the Bcl-2 family play an important role in apoptosis[21].Owing to its significant inhibitory effect on the expression and anti-apoptotic function of Bcl-2,cordycepin can help improve the efficiency of chemotherapeutic drugs.As aforementioned,cordycepin regulates the expression of Bcl-2 and Bax by regulating the Hippo signaling pathway in HepG-2 cells,activates the mitochondrial pathway,releases Cytc and activates caspase-3,and finally promotes apoptosis in HepG-2 cells,thereby resulting in an anticancer effect (Fig.8).
In addition to the inhibition of HepG-2 cell proliferation by cordycepininvitro,we also identified theinvivoand preclinical antitumor efficacy of cordycepin.To this end,we determined theinvivoantitumor efficacy on HepG-2-bearing male nude mice.Our study has demonstrated that cordycepin has strong antitumor activity against HepG-2 xenograft tumors,and has been shown to significantly reduce the volume of xenograft tumors and delay tumor progression.The expression of YAP1,caspase-3,and Bcl-2 proteins in the tumor tissues was subsequently detected,and the results demonstrated that cordycepin could suppress the expression of YAP1 and Bcl-2 in tumor tissues.The positive expression rate of caspase-3 was higher in the tumor tissues of the cordycepin-treated group than in the control group,indicating that cordycepin could induce apoptosis in tumor tissues.Additionally,the dose of cordycepin administered in this study had no detectable toxicity in mice,including no significant effects on the weights of mice,indicating that cordycepin has a satisfactory safety profile.
In this study,the expression of differential genes in HCC cells before and after treatment with cordycepin was determined using a high-throughput sequencing technology and bioinformatics analysis.The possible biological functions of these differential genes in HCC cells and related pathways were further elucidated,and the possible role of the Hippo signaling pathway in the inhibitory effect of cordycepin against HCC was further investigated,which provided novel insights for the treatment of HCC.
5.Conclusion
Cordycepin can act as a natural drug and induce apoptosis in HCC cells bothinvitroandinvivo.Cordycepin may inhibit the expression ofYAP1by promoting the expression ofMST1andLAST1in HepG-2 cells,and regulate the expression ofGBP3andETV5by regulating the Hippo signaling pathway,so as to inhibit the proliferation and migration of HepG-2 liver cancer cells (Fig.9).At the same time,cordycepin regulates the expression of Bax and Bcl-2 through the Hippo signaling pathway,which activates the mitochondrial apoptosis pathway.The release of cytochrome c into the extracellular space activates caspase-3,which eventually leads to apoptosis.GBP3andETV5may be potential targets for the treatment of liver cancer.
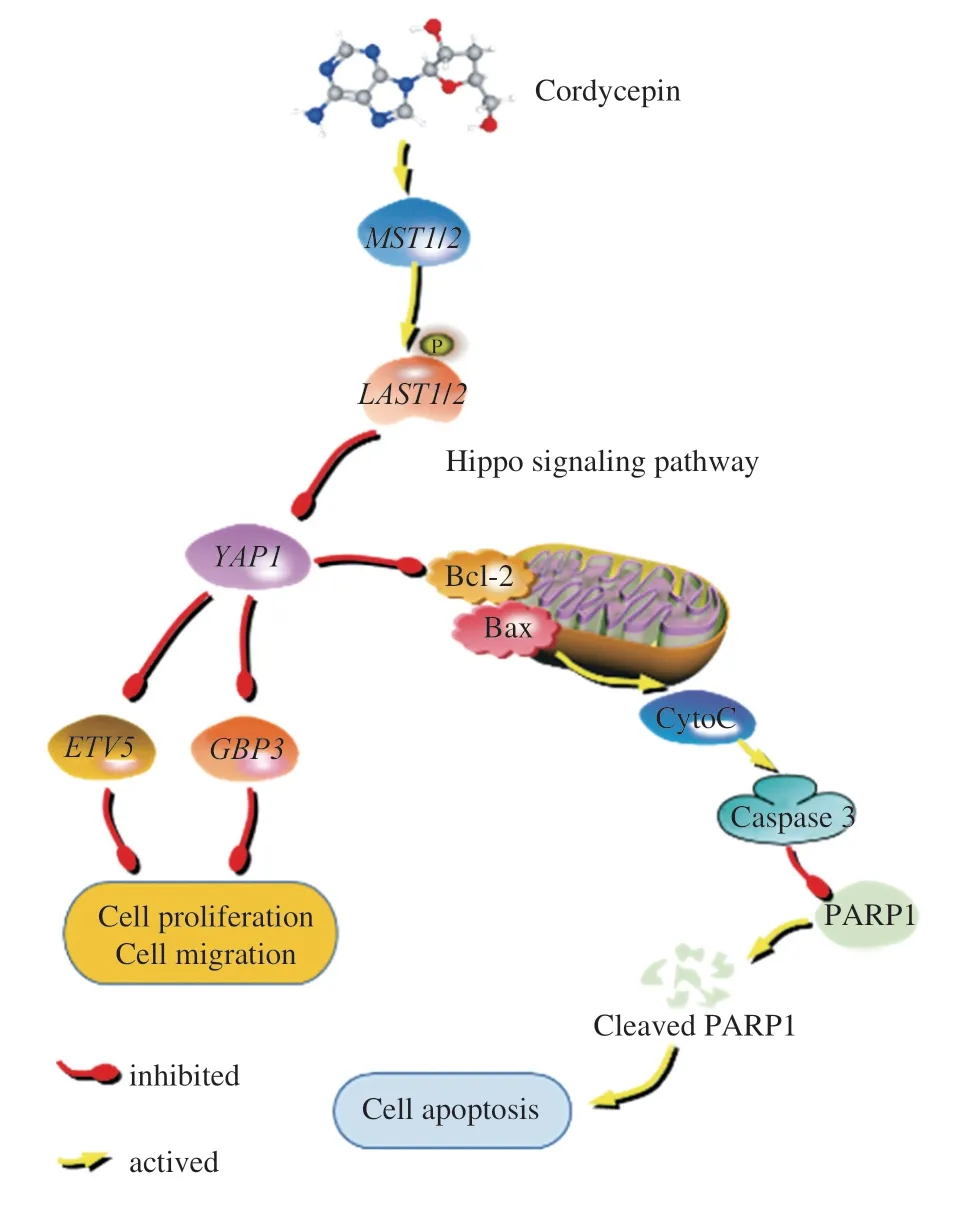
Fig.9 The potential mechanism of action of cordycepin is mediated via the modulation of the Hippo signaling pathway,resulting in the observed anticancer effects of cordycepin.
Conflicts of interest
There are no conflicts to declare.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (81503187).
Appendix A.Supplementary data
Supplementary data associated with this article can be found,in the online version,at http://doi.org/10.26599/FSHW.2022.9250090.

- 食品科学与人类健康(英文)的其它文章
- Protective effects of oleic acid and polyphenols in extra virgin olive oil on cardiovascular diseases
- Diet and physical activity inf luence the composition of gut microbiota,benef it on Alzheimer’s disease
- Inf luence of nitrogen status on fermentation performances ofnon-Saccharomyces yeasts: a review
- Ganoderma lucidum: a comprehensive review of phytochemistry,eff icacy,safety and clinical study
- Resveratrol combats chronic diseases through enhancing mitochondrial quality
- Demonstration of safety characteristics and effects on gut microbiota of Lactobacillus gasseri HMV18