
2 个芹菜品种泛变应原Api g 4 基因的克隆与分析
2013-07-10 03:39李梦瑶侯喜林熊爱生
植物资源与环境学报 2013年1期
李梦瑶,王 枫,侯喜林,蒋 倩,马 静,熊爱生
(南京农业大学园艺学院作物遗传与种质创新国家重点实验室农业部华东地区园艺作物生物学与种质创制重点实验室,江苏南京210095)
肌动蛋白抑制蛋白(profilin)是1 种相对分子质量12 000 ~15 000 的蛋白质,其功能是在植物体内将肌动蛋白单体隔开、抑制肌动蛋白丝的生长,并在植物细胞中参与微丝的快速重组,例如胞质分裂、胞质流动、细胞延长以及花粉管和根毛生长等[1-2]。由于其广泛分布于各种有致敏作用的植物器官中,因而profilin 也被称为泛变应原(panallergen)[3]。泛变应原是一类氨基酸序列高度保守的蛋白质,被认为是引起花粉和植物性食物间发生交叉反应的主要因素之一[1]。自白桦(Betula platyphylla Suk.)中次变应原Bet v 2 被证实为 profilin[4]起,目前已有学者从棕榈〔Trachycarpus fortunei (Hook.) H. Wendl.〕、桃(Amygdalus persica Linn.)、油 橄 榄 (Olea europaea Linn.)和梯牧草(Phleum pratense Linn.)等许多植物中均分离出泛变应原 profilin[5-9]。
旱芹(Apium graveolens Linn.)俗称芹菜,是伞形科(Apiaceae)一年或多年生草本植物,起源于地中海和中东地区,目前在世界范围内均有栽培。芹菜中含有丰富的胡萝卜素、维生素、挥发油及有机酸等多种营养成分,具有很高的食用和药用价值,其中的芹菜素和黄酮等生物活性成分还具有降低血压、调节血脂和降血糖的功效[10-11]。但芹菜是常见的过敏性食物之一,目前已从芹菜中分离出的过敏原蛋白有Api g 1、Api g 2、Api g 4 和 Api g 5[12-13]。因 Api g 4 与白桦中的Bet v 2 具有较高的同源性且能发生变态反应,引起相关科研人员的关注。
鉴于此,作者从形态相近、地域来源不同的2 个芹菜品种‘津南实芹’(‘Jinnanshiqin’)和‘美国西芹’(‘Meiguoxiqin’)中分别克隆出Api g 4 基因,利用生物信息学方法推导其氨基酸序列,并对其氨基酸组成、疏水性、空间结构及其与其他植物的进化关系进行预测与分析,以期为芹菜致敏性及过敏原之间关系的研究提供理论基础。
1 材料和方法
1.1 材料
供试芹菜品种为本实验室保存的‘津南实芹’和‘美国西芹’,种植于南京农业大学人工气候室和江浦实验农场。大肠杆菌菌株DH5α 由本实验室保存;质粒载体 pMD18-T Vector、DNA 分子量 marker、Ex Taq DNA 聚合酶、Prime Script RT reagent Kit 和 SYBR Premix Ex Taq 试剂盒等均为宝生物工程(大连)有限公司生产;RNA simple Total RNA Kit 由天根生化科技(北京)有限公司生产。
实验仪器有超净工作台(苏州安泰空气技术有限公司)、电热恒温培养箱(上海博讯实业有限公司医疗设备厂)、全温震荡培养箱(美国精骐有限公司)、PCR仪(德国Eppendorf 公司)、DYY-6C 琼脂糖凝胶电泳仪(北京市六一仪器厂)、凝胶成像系统(美国Bio-Rad 公司)和ABI7300 型实时荧光定量PCR 仪(美国应用生物系统公司)。
1.2 方法
1.2.1 不同组织中总RNA 提取及cDNA 合成 采用RNA simple Total RNA Kit 分别从‘津南实芹’和‘美国西芹’成熟植株的根、茎和叶等组织中提取总RNA。用Prime Script RT reagent Kit 将总 RNA 反转录成cDNA。
1.2.2 Api g 4 基因的克隆 根据 GenBank 中 Api g 4基因序列(登录号为AF129423)设计1 对引物NXR29和NXR30,序列分别为5'-TTGCATGCGGACGGTGCT GTA-3'和 5'-GGAATAACTAGTCCCGGACATT-3'。以2 个品种的cDNA 为模板进行扩增。PCR 反应程序为:94 ℃ 预变性5 min;94 ℃ 变性30 s,54 ℃ 退火30 s,72 ℃ 延伸60 s,共30 个循环;最后于72 ℃ 延伸10 min。PCR 产物用质量体积分数1.2%琼脂糖凝胶电泳并回收目的条带,连接到pMD18-T 载体上并转化至大肠杆菌 DH5α 中,提取质粒经PCR 鉴定后委托南京金斯瑞生物科技有限公司测序。
1.2.3 实时定量PCR 反应 用芹菜 actin 基因作为参考基因,与目标基因一起扩增,表达检测引物为ACTIN-F 和 ACTIN-R,序列分别为5'-GTTAGTAT ATACCGTCCTTC-3'和5' -GTACTTTAGCTCCACGA CCG-3'。根据2 个品种的Api g 4 基因序列设计表达检测引物NXR81 和NXR82,序列分别为5'-CGAAT TAGACTCCTTTTCTCC-3'和5' -CTACTTGGACAAT GAGGTCC-3'。采用 SYBR Premix Ex Taq 试剂盒并按照操作说明进行实时定量PCR。以芹菜actin 基因的转录表达水平为内参,计算目的基因相对转录表达水平 2-ΔΔCt。其中,ΔΔCt=(Ct靶基因-Ct内参)处理组-(Ct靶基因-Ct内参)对照组(式中,Ct 表示每个 PCR 反应管内荧光信号到达设定域值时所经历的循环数)[14]。
1.3 序列分析
采用Clustal X 软件进行氨基酸组成与多序列比对;利用 GenBank 数据库的 BLASTp 程序计算同源性;使用 MEGA4. 0 软件进行进化树分析[15];通过SWISS-MODEL(http:∥swiss-model. expasy. org)建立蛋白质空间结构模型[16]。
2 结果和分析
2.1 芹菜Api g 4 基因的克隆与序列分析
分别以‘津南实芹’和‘美国西芹’2 个品种的cDNA 为模板、采用 NXR29 和 NXR30 引物进行 PCR扩增,分别得到长度约为400 bp 的片段(见图1),其片段长度与预期片段一致。序列分析结果表明:‘津南实芹’和‘美国西芹’的Api g 4 基因均含有1 个长度405 bp 的开放阅读框(ORF,open reading frame),分别编码134 个氨基酸。其中‘津南实芹’Api g 4 基因的核苷酸序列及其编码的氨基酸序列见图2。2个品种Api g 4 基因的核苷酸序列有3 个碱基的差异,分别为第85 位的 G/A、第149 位的 C/T 和第318 位的C/T;2 个品种的Api g 4 基因编码的氨基酸序列有2个位点的差异,分别为第29 位的Val/Thr 和第50 位的Thr/Ile。
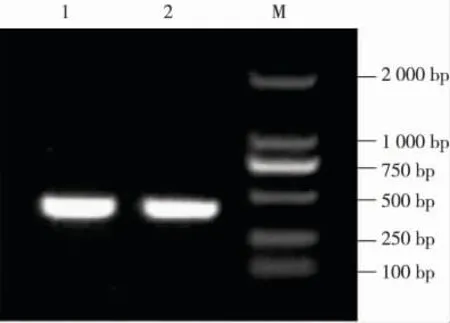
图1 2 个芹菜品种Api g 4 基因cDNA 的PCR 扩增图谱Fig. 1 PCR amplification pattern of cDNA of Api g 4 gene from two cultivars of Apium graveolens Linn.

图2 芹菜品种‘津南实芹’Api g 4 基因的核苷酸序列及其编码的氨基酸序列Fig. 2 Nucleotide sequence of Api g 4 gene from Apium graveolens‘Jinnanshiqin’and its encoding amino acid sequence
2.2 芹菜Api g 4 基因及其他植物泛变应原基因编码的氨基酸序列比较及同源性分析
BLASTp 同源性检索与比对[17]结果表明:‘津南实芹’和‘美国西芹’Api g 4 基因编码的氨基酸序列与欧芹〔Petroselinum crispum (Miller)Nyman ex A.W. Hill〕、胡萝卜(Daucus carota Linn.)、油橄榄(Olea europaea Linn.)、大豆〔Glycine max (Linn.)Merr.〕、欧洲榛(Corylus avellana Linn.)、蓖麻(Ricinus communis Linn.)、栓皮栎(Quercus suber Linn.)、毛果杨(Populus trichocarpa Torr. et A. Gray)、山腚(Mercurialis annua Linn.)、樟树〔Cinnamomum camphora (Linn.)Presl〕、葡萄 (Vitis vinifera Linn.)、垂枝桦 (Betula pendula Roth.)、葎草〔Humulus scandens (Lour.)Merr.〕、玉山筷子芥 (Abaris lyrata Linn.)、烟 草 (Nicotiana tabacum Linn.)、拟南芥〔Arabidopsis thaliana (Linn.)Hey.〕、蒺藜苜蓿(Medicago truncatula Gaertn.)和苹果(Malus pumila Miller)的泛变应原氨基酸序列相似度较高,其中与同科植物欧芹和胡萝卜的同源一致性分别达到94%和91%,与其他种类的同源一致性约为80%。将‘津南实芹’和‘美国西芹’Api g 4 基因编码的氨基酸序列与上述种类泛变应原基因编码的氨基酸序列进行多重比对,结果表明(图3):这些泛变应原的氨基酸序列保守区域较大,且连续相同的氨基酸残基数目较多,最长片段的氨基酸残基数达到8 个,分别为KYMVIQGE 和AVIRGKKG,推测它们可能是潜在的抗原决定簇,能够引发机体的过敏反应。

图3 2 个芹菜品种Api g 4 基因编码的氨基酸序列与其他植物泛变应原氨基酸序列的多重比对结果Fig. 3 Alignment result of amino acid sequences encoded by Api g 4 gene from two cultivars of Apium graveolens Linn. with that of panallergens from other species
为了进一步分析从‘津南实芹’和‘美国西芹’中分离的Api g 4 基因编码的蛋白质与其他植物泛变应原的进化关系,构建了同源进化树(图4),结果表明:‘津南实芹’和‘美国西芹’的Api g 4 蛋白与同科的欧芹和胡萝卜所含的泛变应原蛋白进化关系最近。
2.3 ‘津南实芹’Api g 4 基因编码的氨基酸序列疏水性/亲水性分析
对克隆获得的‘津南实芹’Api g 4 基因编码的氨基酸序列进行了DNAMAN 疏水性/亲水性分析,结果见图5。结果表明:该蛋白的第105 位缬氨酸(Val)疏水性最强,第27 位丙氨酸(Ala)和第26 位丙氨酸(Ala)的疏水性其次;第89 位赖氨酸(Lys)亲水性最强,第88 位甘氨酸(Gly)和第56 位天冬氨酸(Asp)的亲水性其次。总体上看(图5),‘津南实芹’Api g 4 基因编码的蛋白属于疏水性蛋白。此外,对‘美国西芹’Api g 4 基因编码的氨基酸序列的疏水性/亲水性分析结果也显示:‘美国西芹’的Api g 4 基因编码的蛋白也属于疏水性蛋白。
2.4 芹菜Api g 4 蛋白结构预测与分析
以橡胶树〔Hevea brasiliensis (Willd. ex A. Juss.)Muell. Arg.〕的变应原 Hevb8 的 Chain A(PDB ID:1G5U)为模型,通过SWISS-MODEL 进行三维结构同源建模,结果显示(图6):‘津南实芹’和‘美国西芹’泛变应原蛋白Api g 4 与Hevb8 在蛋白质核心区域的氨基酸序列相似性分别为78.2 %和78.9 %,比对参数分别为E-value=8e-60 和E-value=3e-60。‘津南实芹’的Api g 4 蛋白与‘美国西芹’的Api g 4 蛋白有2 个氨基酸位点的差异,二者具有相似的三维空间结构,均具有 3 个 α 螺旋和 7 个 β 折叠。

图4 基于泛变应原氨基酸序列的2 个芹菜品种及其他植物的系统进化树Fig. 4 Phylogenetic tree of two cultivars of Apium graveolens Linn.and other species basis on amino acid sequences of panallergens
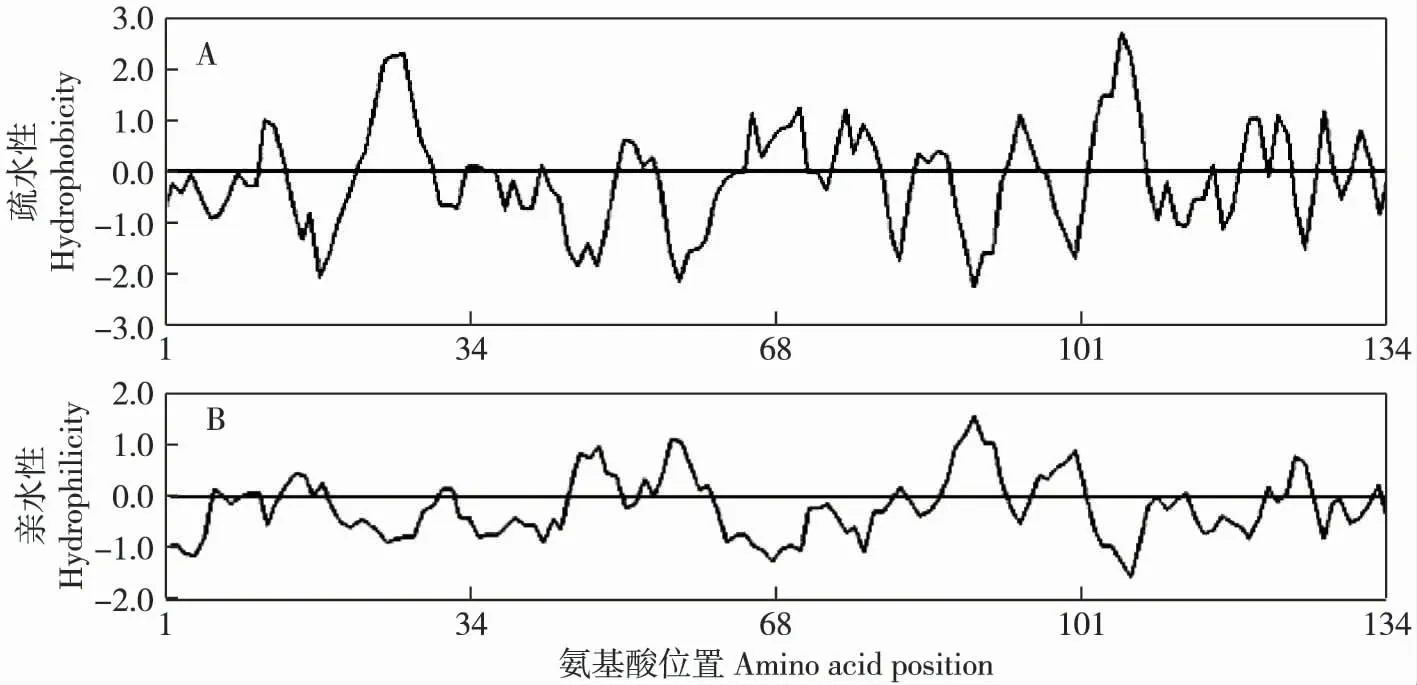
图5 ‘津南实芹’Api g 4 基因编码的氨基酸序列的疏水性(A)和亲水性(B)Fig. 5 Hydrophobicity (A)and hydrophilicity (B)of amino acid sequence encoded by Api g 4 gene from Apium graveolens‘Jinnanshiqin’
2.5 芹菜不同组织中Api g 4 基因的表达分析
通过实时定量PCR 检测Api g 4 基因在‘津南实芹’和‘美国西芹’不同组织中的表达水平,结果见图7。由图7 可见:Api g 4 基因在‘津南实芹’和‘美国西芹’根中的表达水平均最高,在2 个品种的茎和茎尖分生组织中的表达水平相对较低,在叶中的表达水平很弱。由2 个品种间的比较可见:Api g 4 基因在‘美国西芹’茎中的相对表达量显著高于‘津南实芹’茎;在‘美国西芹’茎尖分生组织中的相对表达量则低于‘津南实芹’,由此可见,Api g 4 基因的表达水平在2 个品种间具有一定的组织特异性。

图6 2 个芹菜品种Api g 4 蛋白的三维结构建模Fig. 6 The three-dimensional structural model of Api g 4 protein from two cultivars of Apium graveolens Linn.
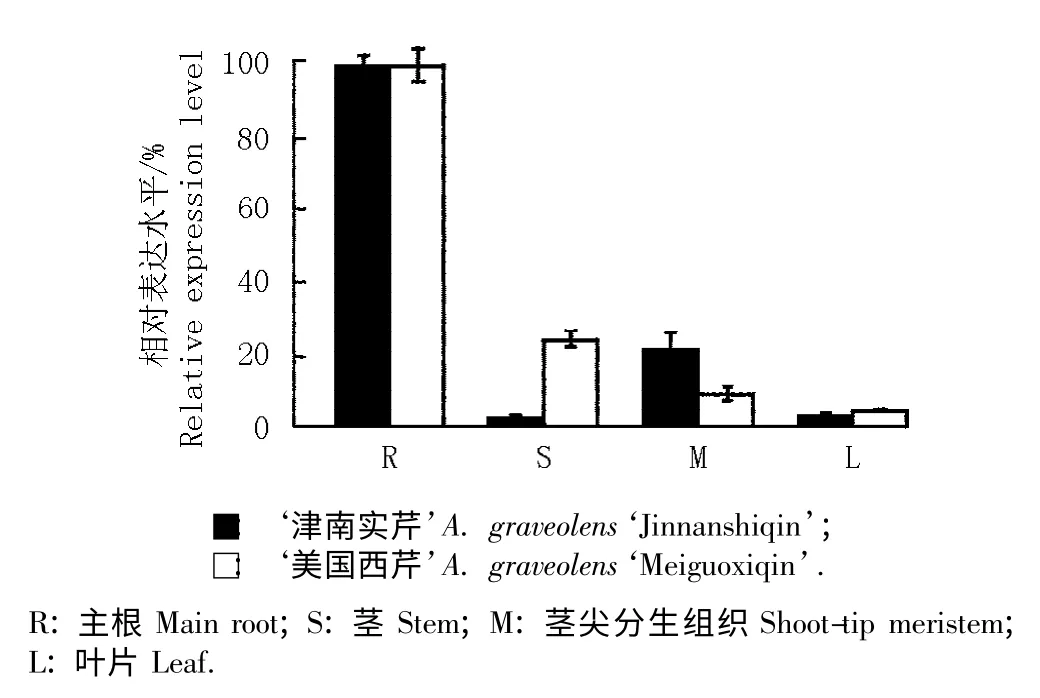
图7 2 个芹菜品种不同组织中Api g 4 基因相对表达量的比较Fig. 7 Comparison of relative expression amount of Api g 4 gene in different tissues of two cultivars of Apium graveolens Linn.
3 讨 论
变态反应(也称过敏反应)是指机体受到抗原物质刺激时发生的一类特殊免疫反应[18]。泛变应原能够引起过敏病人发生变态反应,导致身体组织损伤和免疫功能紊乱[19]。开展对泛变应原的研究、寻找可能引起过敏反应的抗原决定簇的氨基酸序列,能够为从分子水平上改善或修饰变应原提供理论依据。
本实验采用的‘津南实芹’是中国本土培育的品种,而另一品种‘美国西芹’来源于美国,二者虽然起源不同,但在外部形态上有一定的相似性。从‘津南实芹’和‘美国西芹’中获得的Api g 4 基因有3 个核苷酸位点的差异,其编码的氨基酸序列有2 个位点的差异,这些差异可能与某些特定的生理功能有关。
2 个芹菜品种Api g 4 基因主要在根中表达,在茎和茎尖分生组织中的表达量相对较低,在叶中表达很弱;另外,Api g 4 基因在‘津南实芹’和‘美国西芹’茎和茎尖分生组织中的表达量呈现相反的规律,可见该基因的表达具有较明显的种类及组织特异性。Api g 4 基因在植物的生长发育过程中有重要作用,其表达特点可能与泛变应原Api g 4 在植物体内参与不同的生理过程有关。芹菜体内包含的能诱发过敏反应的过敏原较多,过敏机制也较为复杂,引发过敏反应也可能是过敏原间相互作用的结果。
氨基酸序列同源性及进化树分析结果显示:‘津南实芹’和‘美国西芹’的Api g 4 蛋白与其他植物的泛变应原氨基酸序列高度保守,同源性很高。这些泛变应原含有较多连续相同的氨基酸片段,推测这些片段可能为潜在的抗原决定簇,能够引发机体的过敏反应。在进化树上2 个芹菜品种与其同科植物欧芹和胡萝卜归于同一个进化分支,在进化上具有相对较近的关系,这一结果也为探讨伞形科植物的亲缘关系提供了佐证。此外,从进化树上可知:来源于不同种类的泛变应原序列遗传距离较近,表明不同种类间泛变应原蛋白的进化关系较近。据此推测高等植物中的泛变应原蛋白可能是由共同的祖先蛋白进化而来的,并且是高等植物中广泛存在的一类重要蛋白。
[1]VIETHS S,SCHEURER S,BALLMER-WEBER B. Current understanding of cross reactivity of food allergens an pollen[J]. Annals of the New York Academy of Sciences,2002,964(1):47-68.
[2]RAMAEHANDRAN S,CHRISTENSEN H E,ISHIMARU Y,et al.Profilin plays a role in cell elongation,cell shape maintenance,and flowering in Arabidopsis[J]. Plant Physiology,2000,124:1637-1647.
[3]VALENTA R,DUCHENE M,EBNER C,et al. Profilins constitute a novel family of functional plant pan-allergens[J]. The Journal of Experimental Medicine,1992,175:377-385.
[4]VALENTA R,DUCHENE M,PETTENBURGER K,et al. Identification of profilin as a novel pollen allergen;IgE auto-reactivity in sensitized individuals[J]. Science,1991,253:557-560.
[5]ASTURIAS J A,IBARROLA I,FERNÁNDEZ J,et al. Pho d 2,a major allergen from date palm pollen, is a profilin:cloning,sequencing,and immunoglobulin E cross-reactivity with other profilins[J]. Clinical and Experimental Allergy, 2005,35(3):374-381.
[6]RODRÍGUEZ-PEREZ R,FERNÁNDEZ-RIVAS M,GONZÁLEZMANCEBO E,et al. Peach profilin:cloning,heterologous expression and cross-reactivity with Bet v 2[J]. Allergy,2003,58(7):635-640.
[7]SCHEURER S,WANGORSCH A,HAUSTEIN D,et al. Cloning of the minor allergen Api g 4 profilin from celery (Apium graveolens)and its cross-reactivity with birch pollen profilin Bet v 2[J]. Clinical and Experimental Allergy,2000,30(7):962-971.
[8]ASTURIAS J A,ARILLA M C,GOMEZ-BAYON N,et al. Cloning and expression of the panallergen profilin and the major allergen(Ole e 1)from olive tree pollen[J]. Journal of Allergy Clinical Immunology,1997,100:365-372.
[9]ASTURIAS J A,ARILLA M C,BARTOLOME B,et al. Sequence polymorphism and structural analysis of timothy grass pollen profilin allergen (Phl p 11)[J]. Biochimica et Biophysica Acta,1997,1352:253-257.
[10]严建刚,张名位,杨公明,等. 芹菜提取物的降血脂与抗氧化作用的研究[J]. 中国食品学报,2005,5(3):1-4.
[11]邢 渊,丁博儒. 芹菜对糖尿病及并发症的辅助治疗作用[J].实用糖尿病杂志,2006,3(1):60-61.
[12]WÜTHRICH B. Zur Nahrungsmittelallergie. Häufigkeit der symptome und der allergieauslösenden Nahrungsmitteln bei 402 Patienten-Kuhmilchallergie-Nahrungsmittel und Neurodermitis atopica[J]. Allergologie,1993,16:280-287.
[13]GADERMAIER G,EGGER M,GIRBL T,et al. Molecular characterization of Api g 2,a novel allergenic member of the lipidtransfer protein 1 family from celery stalks[J]. Molecular Nutrition and Food Research,2011,55(4):568-577.
[14]PFAFFL M W. A new mathematical model for relative quantification in real-time RT-PCR [J]. Nucleic Acids Research,2001,29:e45.
[15]TAMURA K,PETERSON D,PETERSON N,et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Molecular Biology and Evolution,2011,28:2731-2739.
[16]SCHWEDE T,KOPP J,GUEX N,et al. SWISS-MODEL:an automated protein homology-modeling server[J]. Nucleic Acids Research,2003,31:3381-3385.
[17]ALTSCHUL S F,MADDEN T L,SCHĀFFER A A,et al. Gapped BLAST and PSI-BLAST:a new generation of protein database search programs[J]. Nucleic Acids Research,1997,25:3389-3402.
[18]陈 鹏,郭彦飞,闫 倩. 植物源过敏蛋白[J]. 生命的化学,2011,31(3):465-472.
[19]KAZEMI-SHIRAZI L,NIEDERBERGER V,LINHART B,et al.Recombinant marker allergens:diagnostic gatekeepers for the treatment of allergy[J]. International Archives of Allergy and Immunology,2002,127:259-268.
猜你喜欢
浙江医学(2022年7期)2022-05-16
中山大学学报(医学科学版)(2022年1期)2022-02-28
中国中西医结合耳鼻咽喉科杂志(2021年3期)2022-01-01
科学大众(2021年9期)2021-07-16
医学综述(2021年3期)2021-03-27
农家参谋(2020年11期)2020-01-03
北京广播电视报(2019年7期)2019-03-26
现代园艺(2018年1期)2018-03-15
家庭医药·快乐养生(2017年6期)2017-06-16
恋爱婚姻家庭·养生版(2015年5期)2015-05-14
