
油用向日葵籽实品质性状的遗传研究
2013-09-17 11:58包海柱高聚林胡树平
中国粮油学报 2013年7期
包海柱 高聚林 马 庆 胡树平
(内蒙古农业大学农学院1,呼和浩特 010019)
(内蒙古农牧业科学院2,呼和浩特 010031)
向日葵(Helianthus annuus L.)是世界上仅次于棕榈、大豆、菜籽的重要油料作物和生物能源材料[1],也是我国北方地区的主要油料作物。向日葵油富含人体必需的不饱和脂肪酸,其中亚油酸(顺9,顺12-十八碳二烯酸C18∶2)质量分数约65%左右,油酸质量分数(十八碳-顺 -9-烯酸 C18∶1)约23%[2-3]。现代医学研究表明,亚油酸、油酸具有降血脂,促进细胞生长,免疫调节等生理功能,并且在提高免疫功能及抗肿瘤等方面都表现出潜在的价值[4]。因此,向日葵油的品质性状越来越引起人们的关注。
品质育种是改进向日葵油质的重要途径。目前,关于油用向日葵品质方面的研究还不多见,即便有涉及品质方面的研究,也大多集中在含油率性状上[5-8],且多以胚的核基因为主要研究对象,忽视了细胞质和母体植株的遗传效应;分析方法上,国内外的学者常采用表型分析及世代平均数的分析方法,而这两种方法难以同时分析直接(种子)效应、母体效应和细胞质效应[9];除此之外,以往研究中[5-8],还存在着试验材料或为杂交种,试验设计存在未构建遗传群体等问题。基于上述原因,笔者认为有必要通过改进试验设计、供试材料和分析方法来开展油用向日葵子实品质性状的遗传研究。
作物产量、品质等性状大多属于数量性状。在不同环境下,基因的表达存在着表达类型和程度上的差异性[10]。因此,性状表现会同时受到基因的遗传主效应(加性效应、显性效应等)及其与环境互作效应等多套遗传体系的影响。除此之外,由于植物的种子与其母体植株相差一个世代,种子的营养物质主要靠母体植株提供,因此种子性状的遗传各加复杂化[9]。针对此问题,Zhu J 等[11-12]采用混合线性遗传模型,提出了具有生物学意义的,包括种子核基因、母体植株核基因和细胞质基因在内的3套遗传效应的遗传模型,用于分析二倍体种子性状。
本研究以油用向日葵为材料,通过构建遗传群体及交配体系,采用朱军[9],Zhu J等[11-12]提出的包括与环境互作在内的二倍体种子性状的混合线性遗传模型以及统计方法,首次对油用向日葵籽实含油率、亚油酸含量及油酸含量等品质性状进行了遗传分析,旨在揭示油用向日葵籽实品质性状的遗传规律,为油用向日葵品质育种与改良提供参考。
1 材料与方法
1.1 试验材料
本研究以9个不同基因型自交系为亲本,按5×4不完全双列杂交(NC designⅡ)方式构建了遗传群体及交配体系,组配出20个F1杂交组合及20个F2代材料,加上9个亲本构建了49个供试遗传群体。
该9个自交系由内蒙古农业大学所选育;材料间无血缘关系,并经过了一般配合力(GCA)测定,籽实品质性状存有差异,并且在生物学性状、农艺性状、产量构成等方面有丰富的遗传基础,能够保证所构建群体的遗传多样性。
1.2 试验设计
试验于2012年在呼和浩特市内蒙古农业大学教学农场(E111°39',N40°49')和包头市土默特右旗萨拉齐镇(E110°31',N40°33')两地同时进行。两处试验地块均为沙壤土,肥力中等,地势平坦,具有排灌条件。田间种植采用Lattice(7×7)平衡设计,试验小区为6.0 m行长,0.7 m行距,5行区,小区面积21.0 m2,保苗6.0 ×104株/ha,3 次重复,生育期的田间管理与当地大田生产技术相同。
1.3 籽实品质性状的测定
成熟期小区单收脱粒(亲本植株上获得是亲本籽实,F1植株上获得是F2代籽实,F2植株上获得是F3代籽实),晾晒、清选后,用钟鼎式分样器取样。取样结束后,统一测定粗脂肪含量(含油率)和亚油酸含量、油酸含量。参照蔡乾蓉等[13]的方法测定含油率,亚油酸含量、油酸含量的测定参照张志军等[14]的方法。
1.4 数据统计分析
本研究中应用包括基因与环境互作效应在内的二倍体种子混合线性遗传模型对油用向日葵籽实品质性状进行分析。性状观测的表型值Y分解如下[9]
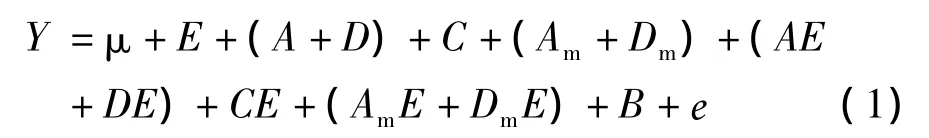
方程(1)中,μ为群体均值,E是环境效应,A为直接加性效应,D为直接显性效应,C是细胞质效应,Am是母体加性效应,Dm是母体显性效应,AE为直接加性×环境互作效应,DE为直接显性×环境互作效应,CE为细胞质 ×环境互作效应,AmE是母体加性×环境互作效应,DmE为母体显性×环境互作效应,B为环境内的区组效应,е为残余效应。
相似于方程(1),对于指定性状T2,T1的单个条件表型值能分解为[15]

方程(2)中,指定性状间的条件方差分量包含性状T1排除性状T2影响后额外的遗传变异量,通过计算其净效应的遗传贡献率=1-/VT2)来揭示指定性状T2对T1的遗传贡献程度[16],此模型在本研究中用于分析籽实含油率、亚油酸含量、油酸含量之间的相互遗传贡献,进而说明控制品质性状的基因关系。所分析的遗传贡献率包括,加性贡献率(CrA)、显性贡献率(CrD)、细胞质贡献率(CrC)、母体加性贡献率(CrAm)及母体显性贡献率(CrDm)。
统计分析中采用Jackknife法抽样技术、最小范数二阶无偏估算(MINQUE)法估算观测值的各项方差分量,包括加性方差(VA)、显性方差(VD)、细胞质方差(VC)、母体加性方差(VAm)、母体显性方差(VDm)、加性 ×环境方差(VAE)、显性 ×环境方差(VDE)、细胞质×环境方差(VCE)、母体加性×环境方差()、母体显性×环境方差()、机误方差(Ve)及表型方差(Vp)。应用调整无偏预测(AUP)法预测观测值的遗传效应值。应用SAS9.0和QGA运算程序[9]对数据进行处理。
2 结果与分析
2.1 油用向日葵亲本及杂交后代籽实品质性状的平均表现及差异性检验
油用向日葵在不同环境下籽实含油率、亚油酸含量、油酸含量的平均表现列于表1,方差分析结果列于表2。方差分析表明,供试材料除亲本籽实含油率和F3籽实亚油酸含量在不同环境(地点)的差异未达显著水平外,其余品质性状的环境效应(地点)、品种效应及品种×环境效应均达0.01极显著水平或0.05显著水平的差异,说明所观测的3个品质性状在不同环境下的遗传变异幅度较大,所构建的遗传群体材料适合对含油率、亚油酸、油酸等品质性状做进一步的相关遗传分析。
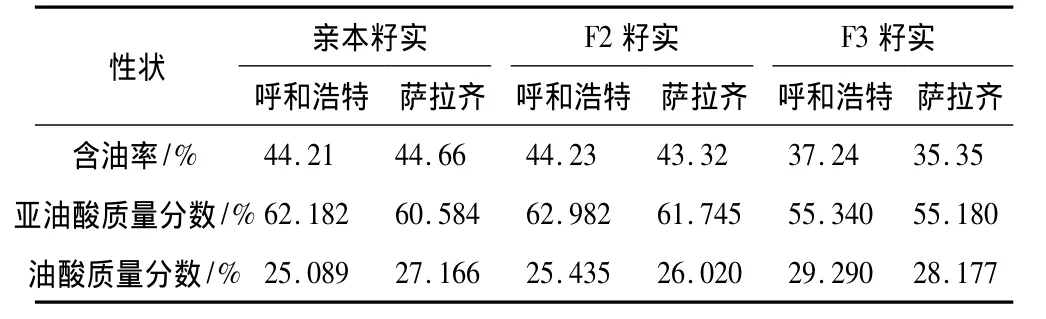
表1 供试材料籽实含油率、亚油酸含量及油酸含量平均表现
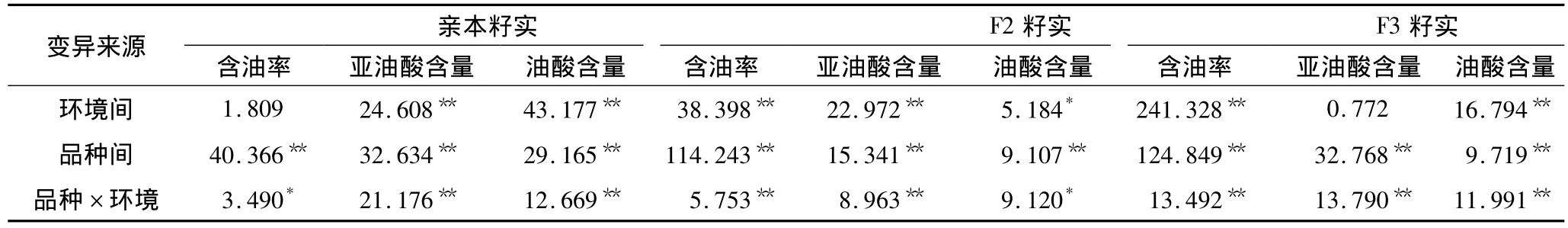
表2 供试材料籽实品质性状方差分析(F值)
2.2 油用向日葵籽实品质性状的遗传效应分析
不同环境条件下,油用向日葵籽实3个品质性状的各遗传方差分量见表3。表3显示,种子核基因对3个品质性状的遗传效应不同,含油率未检测到直接加性效应,油酸含量未检测到直接显性效应,亚油酸含量则未检测到直接遗传效应(VA+VD)。母体效应(VAm+VDm)、细胞质基因效应(VC)在3个品质性状上的效应均达到极显著水平。由此说明,油用向日葵籽实含油率、亚油酸含量、油酸含量等品质性状的表达会同时受到胚、细胞质及母体植株等3套遗传体系基因的影响。
由表3可看出,含油率、亚油酸含量的细胞质效应方差最大,母体效应方差(VAm+VDm)次之,且细胞质效应方差(VC)远大于相应的直接遗传效应(VA+VD)方差,说明细胞质基因效应和母体基因效应是控制含油率和亚油酸含量表现的主要遗传效应,含油率、亚油酸含量的细胞质基因效应占各自遗传主效应(VG=VA+VD+VC+VAm+VDm)的 77.78% 和69.16%。由于细胞质效应可通过母本传递,因此,在油用向日葵籽实品质改良中对含油率和亚油酸含量通过母本进行选择是有效的。另外,含油率、亚油酸含量及油酸含量的母体显性效应方差(VDm)均达到了极显著水平,表明母体杂种优势也是影响品质性状的一个因素。各性状的机误方差分量均达到极显著水平,说明环境误差或者抽样误差对测定值有一定的影响。
研究中还发现,各性状的表现受到了不同程度的基因与环境互作效应影响,含油率、亚油酸含量和油酸含量的基因与环境互作效应(VAE+VDE+VCE++),分别占到了表型方差(Vp)的0.605%、2.186%和42.094%,说明基因与环境互作效应对含油率、亚油酸含量的影响小,而油酸含量的表现易受环境条件影响。
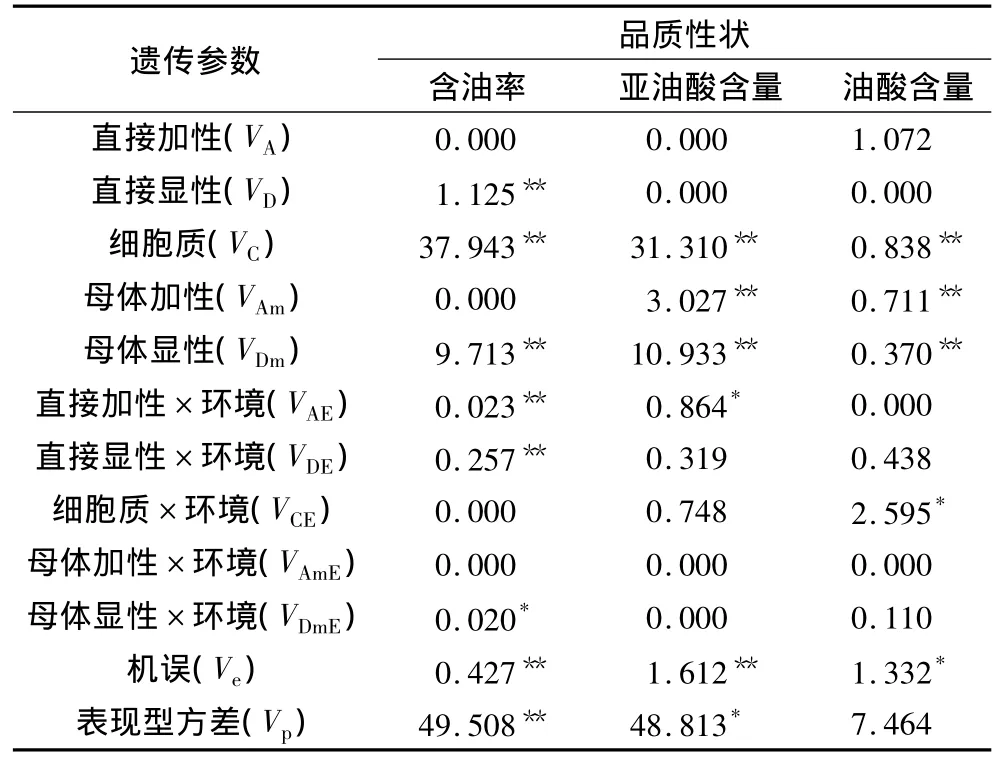
表3 油用向日葵籽实品质性状方差分量估算值
2.3 油用向日葵籽实含油率、亚油酸含量及油酸含量的遗传相关分析
在生物界,广泛存在着基因与环境互作的遗传效应,性状间的相关性会受到环境的影响。因此,从表型相关中将遗传相关分离出来,有助于解析性状间本质的遗传关系和制定适当的育种策略。基于这种考虑,本研究将基因型相关分解为直接加性相关(rA)、直接显性相关(rD)、细胞质相关(rc)、母体加性相关(rAm)、母体显性相关(rDm)、直接加性×环境相关(rAE)、直接显性×环境相关(rDE)、细胞质×环境相关(rCE)、母体加性×环境相关()及母体显性×环境相关()等10项相关分量。
油用向日葵籽实3个品质性状间的遗传相关分析见表4。由表4可知,表型相关系数(rp)与遗传相关系数(rG)均达到极显著水平,基因型相关系数大于表现型相关系数,二者在数值上较为接近。含油率与油酸含量之间、亚油酸含量与油酸含量之间存在极显著的表型负相关和遗传负相关;含油率与亚油酸含量之间存在极显著的表型正相关和遗传正相关;残余效应或随机误差对结果有一定影响(re=0.171**);直接加性和直接显性相关在3个品质性状之间的均未被检测到(rA=0,rD=0)。
含油率与油酸含量的遗传负相关,主要归因于有极显著水平的细胞质和母体显性负相关。由此表明,母体杂优效应及细胞质效应对含油率和油酸含量的表现存在反向表达。
含油率与亚油酸含量的遗传正相关主要归因于二者间有较大的细胞质、母体显性效应的正相关,表明母体杂优效应与细胞质效应对含油率与亚油酸含量的表现存在正向表达。又因细胞质效应可以通过母本传递,因此在配制杂交种时,能够做到同时利用含油率和亚油酸含量的杂种优势。可见,在油用向日葵品质育种中,通过母本(不育系)的选择可同步提高含油率和亚油酸含量的表现。需要注意的是,这两个性状间存在微弱的直接显性互作负相关(rDE=-0.208*),在某些特殊环境中,在进行高含油率的自交系选育时有可能会降低亚油酸含量。
表4还显示,亚油酸含量与油酸含量之间存在极显著水平的细胞质、母体加性、母体显性及直接显性×环境互作效应的负相关,表明不论是杂种优势利用还是自交系的杂交选育,在后代中同步提高亚油酸和油酸含量的可能性不大。
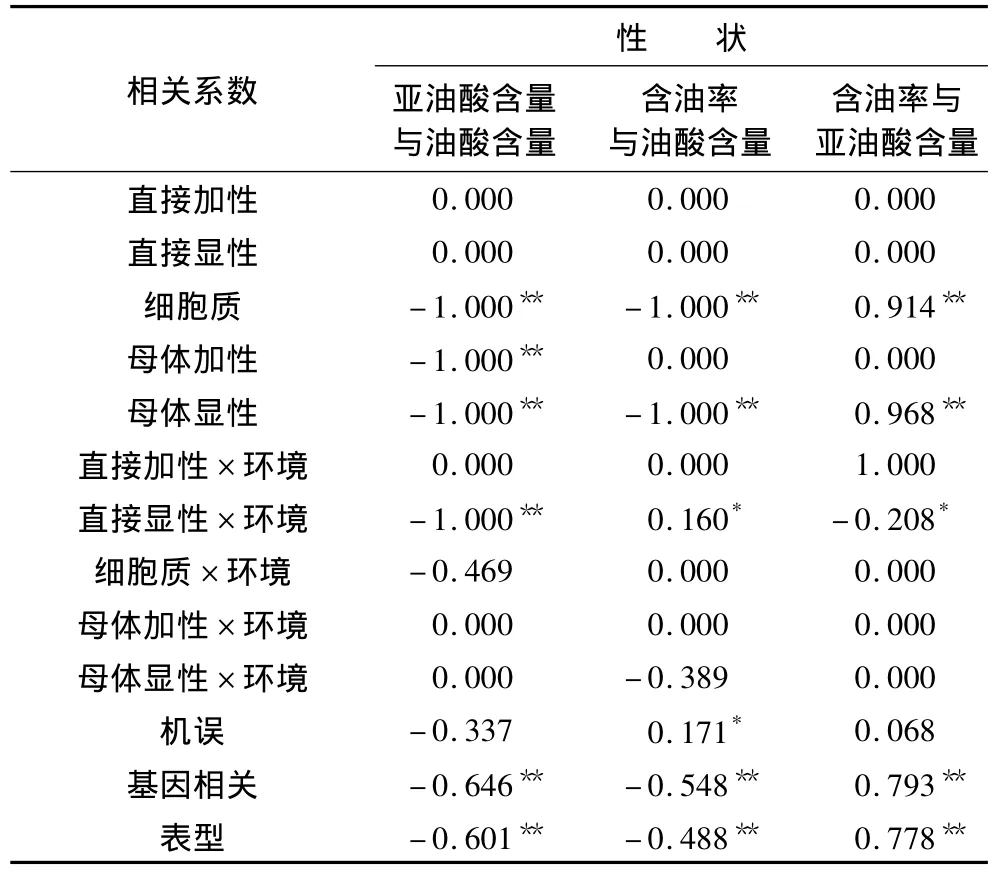
表4 油用向日葵籽实品质性状间的遗传相关系数估算
2.4 油用向日葵籽实含油率、亚油酸含量及油酸含量的遗传率分析
前文的分析表明,油用向日葵籽实品质性状受胚、细胞质、母体基因及环境互作的影响。故,可将狭义遗传率可分解为普通狭义遗传率和互作狭义遗传率[9],其中普通狭义遗传率表达式为,互作狭义遗传率表达式为为普通狭义直接遗传率为普通细胞质遗传率为普通母体遗传率为直接环境互作遗传率为细胞质环境互作遗传率,为母体环境互作遗传率。由于这两种狭义遗传率在杂交后代的选择中都是有效的,因此根据二者数量值的大小,可明确各遗传分量在总的遗传率中的相对重要性。
油用向日葵籽实含油率、亚油酸含量及油酸含量的遗传率估计值见表5。分析表明,含油率、亚油酸含量的普通狭义遗传率分别为76.64%和70.34%;互作狭义遗传率(=)分别为0.10%和3.30%,这两个性状的普通狭义遗传率远大于各自的互作狭义遗传率,并且主要归因于各自的细胞质遗传率)。因此对选系而言,这两个性状的选择适宜在杂交早期世代进行,并根据母体植株品质性状的总体表现进行选择,能够取得较好的改良效果。
油酸含量的各狭义遗传率分量中,达到显著及以上水平的只有细胞质互作遗传率表明油酸含量的选择适宜在特定环境或多点条件下通过母体选择,有可能会取得较好的育种效果。

表5 油用向日葵籽实含油率、亚油酸含量及油酸含量的遗传率估算值
2.5 油用向日葵籽实3个品质性状互为条件的基因效应分析
基因连锁或一因多效等原因造成了性状间的遗传相关,了解性状间相互遗传贡献及基因间表达关系,对育种实践中性状的直接或间接选择以及多个性状的同步改良具有重要意义。前文分析了含油率、亚油酸含量及油酸含量之间的遗传相关性。现在此基础上,分析此3个品质性状彼此间的遗传贡献。在分析之前参照温永仙[15]的方法,做如下说明:(1)若性状T2对T1的遗传贡献率与T1对T2的遗传贡献率都达到显著水平,说明控制性状T1和T2的遗传基因是连锁的或是一因多效的;(2)若性状T2对T1的遗传贡献率达到显著水平,而T1对T2的遗传贡献率未达到显著水平,说明控制性状T2的基因同时影响性状T1的基因,而控制性状T1的基因独立于控制性状T2的基因;(3)若性状 T2对 T1的遗传贡献率与性状T1对 T2 的遗传贡献率都未达到显著水平,说明控制性状T1和性状T2遗传基因是相互独立的。
3个品质性状彼此间的遗传贡献率见表6,未达到显著性水平的贡献率在表中未列出。由表6可知,油酸含量对含油率的细胞质遗传效应有较大贡献率(CrC=0.997 1**),而含油率对油酸含量的细胞质遗传贡献率未被检测到显著水平,表明控制油酸含量的细胞质基因同时影响着含油率的表达,而控制含油率的细胞质基因独立于控制油酸含量的细胞质基因;对于母体效应而言,控制油酸含量的的母体加性基因独立于控制含油率的母体加性基因,油酸含量与含油率彼此间的母体显性遗传贡献率都达到极显著水平,表明控制这两个性状的母体显性基因存在基因连锁或一因多效现象。
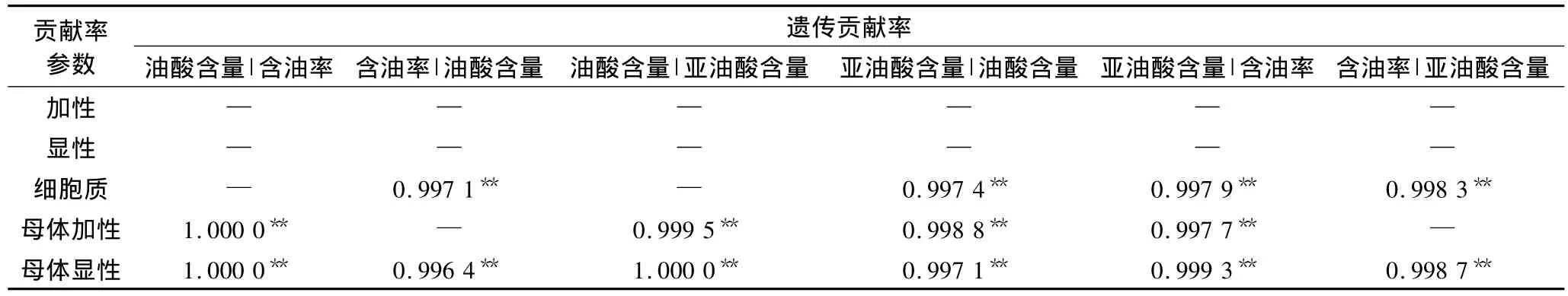
表6 油用向日葵籽实含油率、亚油酸含量及油酸含量的相互遗传贡献率估计值
油酸含量与亚油酸含量彼此间的母体效应(母体加性+母体显性)的相互遗传贡献率均达了极显著水平,表明控制油酸含量与亚油酸含量的母体加性基因和母体显性基因均存在基因连锁或一因多效;对二者的细胞质基因效应而言,控制油酸含量的细胞质基因能够影响亚油酸含量的的表达,而控制亚油酸含量的则独立。
控制含油率和亚油酸含量的细胞质基因、母体显性基因对彼此的遗传贡献率均达了极显著水平;控制含油率的母体加性基因同时影响着亚油酸含量表达,而控制亚油酸含量的母体加性基因则是独立,表明控制这两个性状的细胞质基因和母体显性基因存在基因连锁或一因多效现象。
3 讨论与结论
在植物种子的建成及发育过程中,母体植株为其提供了所需要的全部营养物质。因此,种子的性状除受控于种子本身二倍体胚核基因外,还可能受到二倍体母体植株核基因的控制。另外,细胞质基因也可能通过控制叶绿体(线粒体)的合成而影响植物的光合作用或呼吸来间接控制种子的性状表现[9]。
本研究中,通过对油用向日葵籽实品质性状的分析发现,含油率、亚油酸含量、油酸含量等3个籽实品质性状的表现受到胚、细胞质及母体植株等遗传体系的影响。含油率和亚油酸含量的表现主要受控于细胞质效应基因和母体效应基因,油酸含量以细胞质互作效应为主。其中,母体显性效应和细胞质效应在3个品质性状上均被检测到极显著水平,因此,在油用向日葵籽实品质改良中,利用杂种优势和细胞质效应能通过母本传递的特点,通过母本进行3个品质性状的选择和改良是有效。
对作物性状进行遗传相关分析有益于解析性状间复杂的协同关系,便于杂交育种中对性状的直接或间接选择与评判。由于加性效应具有能在后代中累加和稳定遗传的特点,对于直接加性相关显著的成对性状,可以在早期世代进行间接选择与固定;而对以显性相关为主的成对性状,宜于在高代进行间接选择或通过杂优利用来表达;而对有较高环境互作相关的成对性状则应在不同环境下进行选择[16]。
研究中发现,含油率与油酸含量之间存在极显著的细胞质、母体显性的负相关;亚油酸含量与油酸含量之间存在极显著的的细胞质、母体(母体加性、母体显性)及直接显性互作负相关。因此,不论是杂种优势利用还是自交系的杂交选育,对含油率与油酸含量成对性状以及亚油酸含量与油酸含量成对性状进行同步改良的难度较大。对于含油率与亚油酸成对性状而言,由于二者之间存在极显著的正向母体显性和细胞质效应相关,因此,通过母本(不育系)的选择可同步提高杂交种的含油率和亚油酸含量,另外在自交系杂交选育时,也可考虑利用这种相关性,因为母体显性效应会对后代产生延迟遗传现象,即F1植株上收获的F2种子的品质性状将不会有大变异。遗传率分析表明,含油率、亚油酸含量的细胞质遗传率()最高,受环境影响小,根据母体植株上品质性状的总体表现进行早代选择能取得较好的改良效果。值得注意的一点是,亚油酸含量与油酸含量之间存在遗传负相关性(rG= -0.646**),含油率与油酸含量之间存在微弱的显性互作(rDE=0.160*),因此,在特定环境中提高含油率的同时,有可能会相对提高油酸含量。
成对性状间互为条件的贡献率分析表明,控制含油率的细胞质基因独立于控制油酸含量的;控制油酸含量的母体加性基因独立于含油率的,因此在广谱环境下不适宜对这两个性状进行同步改良。油酸含量与亚油酸含量的母体效应基因存在连锁或为同一基因,又因这两个性状间是负相关。所以,如果选择了高亚油酸含量为目标,势必会产生低油酸含量结果,反之亦然。控制含油率、亚油酸含量的细胞质基因和母体显性基因皆存在着基因连锁或一因多效现象,又因二者之间的正向相关性,说明无论是自交系的杂交选育还是不育系的回交转育,再到杂优利用的杂交种配制,通过对母本的正确选配,能够容易获得高含油率、高亚油酸含量的自交系或“三系”杂交种。综上诉述不难看出,在进行油用向日葵的品质育种时,如何突破母本(不育系)的品质性状是至关重要的一个环节。
[1]J,Feng,B,A,Vick,Mi- Kyung Lee,.et al.Construction of BAC and BIBAClibraries from sunflower and identification of linkage group - specific clone by overgo hybridization[J].Theor.Appl.Genet.,2006(113):23 -32
[2]Belhassen E,Auge G,Ji J,et a1.Dynamic management of genetic resources:first generation ayalysis of sunflower artifical populations[J].Genet Sel Evo1.,1994,26(1)L:241 -253
[3]刘公社,阿兰.博让,等.向日葵研究与开发[M].北京:中国科学技术出版社,1994:1-2
[4]郭传瑸,马大权.不饱和脂肪酸的抗肿瘤作用[J].中国临床营养杂志,2000(8):2,133 -135
[5]Q Ahmad.The effects of some agronomic traits and seed characteristics on yield of sunflower[J].Euphytic,1992,2:11 -14
[6]孙敏,高新梅,刘壮,等.油用型向日葵子实产量与农艺性状相关及通径分析[J].中国油料作物学报,2008(10):273-274
[7]雷中华,向理军,石必显.向日葵9个主要性状之间相互关系分析[J].新疆农学,2006,43(1):31-33
[8]谢宗铭,李万云,陈寅初,等.油葵自交系子实含油率性状配合力的鉴定[J].新疆农业科学,2003,40(3):145 -147
[9]朱军.遗传模型分析方法[M].北京:中国农业出版社,1997:130-150
[10]吴国海.玉米几个数量性状在不同发育阶段的基因效应分析[J].遗传学报,1987,14(5):363 -369
[11]ZHU J.Mixed model approaches for estimating genetic variances and covariances[J].Journal of Biomathematics,1992,7(1):1-11
[12]ZHU J,Weir B S.Analysis of cytoplasmic and maternal effects:I.a genetic model for diploid plant seeds and animals[J].Theor.Appl.Genet.,1994,89(2):153 -159
[13]蔡乾蓉,吴卫,郑有良,等.紫苏属籽粒含油率及其脂肪酸分析[J].中国粮油学报,2009,24(1):84-87
[14]张志军,张鑫,李会珍,等.发育紫苏种子营养累积与脂肪酸组分变化[J].中国粮油学报,2011,26(5):30 -32
[15]温永仙.基于遗传贡献率的陆地棉营养品质性状动态发育研究[J].生物数学学报,2007,22(3):539-546
[16]肖炳光,朱军,卢秀萍,等.烤烟主要农艺性状的遗传与相关分析[J].遗传,2006,28(3):317 -323.
猜你喜欢
课外生活·趣知识(2022年2期)2022-02-08
作物研究(2021年4期)2021-09-05
西南石油大学学报(自然科学版)(2021年3期)2021-07-16
江苏农业科学(2019年7期)2019-09-17
高考·中(2019年6期)2019-09-10
中国洗涤用品工业(2019年4期)2019-05-11
消费导刊(2018年15期)2018-11-24
中国新技术新产品(2017年15期)2017-07-07
科学中国人(2016年30期)2016-07-14
科技创新导报(2016年7期)2016-05-14
