
提取−消退范式中复合刺激对恐惧消退的影响*
2017-02-01 02:54庄楚群2王文清胡静初张蔚欣王鹏贵郑希付
心理学报 2017年3期
庄楚群,2王文清胡静初张蔚欣王鹏贵郑希付
(1华南师范大学心理学院, 广州 510631) (2广东广雅中学, 广州 510160)
1 前言
识别环境中的潜在危险因素对人类的生存有着重要的作用, 而当恐惧学习产生不适应性时, 会导致过度的恐惧和焦虑, 给个体生活带来困扰, 如创伤后应激障碍PTSD (post-traumatic stress disorder)。因此, 探讨如何有效消退由创伤经历引起的恐惧记忆,降低恐惧反应对人类有着重要的理论意义和应用价值。
当前探讨恐惧习得与消退的模型主要基于巴甫洛夫的经典条件反射作用, 即将条件刺激(中性刺激, CS, 如:声音)与非条件刺激(恐惧刺激, US,如:电击)进行多次匹配联结训练后(CS-US), 单独呈现CS即可引发恐惧反应; 而当CS多次独立呈现而不匹配US (CS-no US)时, 对CS的恐惧反应会逐渐消退, 这即为传统的恐惧消退训练方法(Davis,Walker, & Myers, 2003)。传统的恐惧消退训练方法虽能有效地抑制恐惧记忆的表达, 但这种方法存在恐惧返回的现象, 如自发恢复、续新、重建等效应(Myers & Davis, 2002, 2007)。近年来, 研究者们将焦点聚集在条件性恐惧记忆的再巩固过程, 提出了提取−消退(retrieval-extinction, ret-ext)的行为干预范式(Monfils, Cowansage, Klann, & LeDoux, 2009)。
传统的记忆理论认为, 记忆只需要一次的巩固就可以形成稳定; 而记忆再巩固理论(reconsolidation)认为, 记忆在每一次的提取激活后都会暂时重返不稳定状态, 必须经过一个新的巩固阶段才能保持,在此阶段里需要有新的蛋白质合成; 若在这一阶段进行药物或行为干预, 原有记忆很容易受到干扰,从而可能改写(rewrite)或清除(erase)原有的记忆(Nader, Schafe, & LeDoux, 2000; Walker, Brakefield,Hobson, & Stickgold, 2003)。从进化的观点来解释,记忆再巩固是一种适应更新机制, 在激活原有记忆之后允许新的信息融入到原有记忆当中, 以便更好的适应当前的生存环境(Schiller et al., 2010)。Monfils等(2009)首次提出了基于记忆再巩固的提取−消退行为干预范式, 该范式是通过单独呈现一个条件刺激 CS激活原有恐惧记忆, 进入再巩固时间窗后再利用传统的消退训练将新的安全信息融入到长时记忆中, 从而弱化原有的恐惧记忆, 阻断或抑制恐惧反应(Monfils et al., 2009; Auber,Tedesco, Jones, Monfils, & Chiamulera, 2013)。
在以往关于巴甫洛夫的条件性恐惧消退的研究中, 研究者们大多数是采用一个单独的刺激作为CS来诱发被试的恐惧反应, 再利用这个单独的CS进行传统的消退训练或提取−消退(Bradley, Greene,Russ, Dutra, & Westen, 2005); 也有研究者采用多个同种感官刺激CS, 每个CS单独与同一个非条件刺激US形成条件性恐惧, 并证明同时呈现多个CS进行传统的消退训练效果更好(Rescorla, 2000;Witnauer & Miller, 2012)。在现实情境中, 预测危险的信号可能是有多个不同的线索, 也可能来自多个感官通道, 而在临床干预时, 只能重现其中的一个或一种线索。因此, 本研究将尝试把提取−消退范式应用到更复杂的恐惧记忆中, 采用多感官复合刺激(声音+图片)作为条件刺激和提取线索, 探究采用单个线索、复合线索的提取试次, 能否有效地减少复合刺激的条件性恐惧记忆, 以及两者之间有何差异, 提高提取−消退范式在心理治疗中的临床应用价值。
2 方法
2.1 被试
被试为 40名本科院校大学生, 通过自愿报名参加实验。被试均为右利手, 无躯体疾病及精神障碍, 视力或矫正视力正常, 无色盲色弱, 无听力障碍, 且之前没有参加过类似情境实验。
实验前向被试说明:实验过程中会在手臂处接受轻微电击, 电击仪器的使用是经过科学评定的,绝不会对您造成任何伤害, 如果在实验过程中有任何不适症状, 可随时提出终止实验。三天实验结束后将支付您相应的报酬。
被试需签署实验知情同意书, 并填写《状态−特质焦虑量表》。被试随机分配进入“声音提取−消退”组(T 组), “图片提取−消退”组(P 组)及“复合线索提取−消退”组(T+P组)。实验过程中有3名被试未能准时参加实验或未按要求操作, 在数据分析中予以剔除, 故有效被试共有37名, 其中T组12人(4名男生), P组12人(5名男生), T+P组13人(5名男生)。被试年龄范围为 18~22岁(M=19.78,SD=1.18)。状态−特质焦虑量表结果显示:3组被试在状态焦虑分量表(F(2,34)=2.63,p> 0.05)和特质焦虑分量表(F(2,34)=2.98,p> 0.05)上的得分均无显著差异。
2.2 实验材料
本实验采用两组图片和声音组合作为条件刺激(CS), 图片分别为蓝色正方形和黄色正方形, 两个图片具有相同的亮度, 背景为白色(参考 Schiller等2010年研究), 声音分别为不同乐器(钢琴和吉他)弹奏的一段音乐, 两组声音具有相同的响度和音调(材料评定阶段, 95.7%被试能清晰分辨这两种声音)。两组条件刺激呈现时间为4000 ms。其中一组条件刺激在 38%的试次后会跟随 US, 称作 CS+;另一组则始终不会跟随 US, 称作 CS−。为平衡图片和声音对不同被试间的影响, 在实验中, 图片和声音的四种不同组合有相同的概率充当CS+。参考Schiller等人(2010)的研究, 本实验采用腕部电击作为非条件刺激 US来诱发被试的恐惧情绪, 电击持续时间为 200 ms, 电击强度会按照被试的疼痛敏感度在正式实验开始前进行评定。
2.3 测量指标
使用 Spirit NeXus-10型生物记录仪系统记录被试的皮肤电。将Ag/AgCl电极分别缠在被试左手食指和无名指的末端指腹上, 电极连接在生理记录仪的GSR100C模块上记录皮电, 采样率为120 Hz。所采集的生理数据进行离线处理。数据处理的方法参考Schiller团队:取CS呈现后500 ms ~ 4500 ms时间窗内的最高值和最低值, 两者之差为 CS诱发的皮电值, 在此之后, 所有被试的皮电值要进行范围校正, 除以每个被试的US诱发皮电值的平均值,最后所有数据平方根化以减少分布的偏度。第一天的恐惧习得阶段只分析那些不跟随US的CS+试次(Schiller et al., 2010)。
2.4 实验程序
电击强度评定:在正式实验之前先对每个被试所能接受的电击强度进行评定, 这个强度能引起被试不舒服感但不至于感到疼痛。评定先从较微弱的10 V开始, 之后逐渐增大强度, 直到被试报告强度已达到不舒服感但不至于感到疼痛(电击强度最高不超过50 V)。每次电击持续时间为200 ms, 电流为每秒 50脉冲。在接下来的各个实验环节中被试的电击强度不再改变。
在条件性恐惧的正式实验中, 所有材料都通过Eprime 2.0编程呈现。首先在屏幕中央呈现红色注视点“+”, 呈现时间为1000 ms, 注视点后呈现条件刺激CS (图片直接呈现在电脑屏幕, 声音由被试佩戴耳机获取), 呈现时间为4000 ms, 随后跟随非条件刺激US或者白屏, 呈现时间均为200 ms (如图1所示)。试次间的间隔(ITI)为10~12 s (Schiller et al.,2010)。条件刺激CS出现的次序以准随机的方式编排, 确保每个 CS不会重复出现两次以上。实验过程中用皮肤电仪器记录被试的 SCR值, 被试不需要做任何按键反应。本实验分为3天, 分别为第一天的习得阶段、第二天的提取−消退阶段、第三天的自发恢复−重建测试阶段。正式实验开始前, 主试先向被试详细解释指导语, 确保被试明白指导语的意思后, 进入第一天的习得阶段。
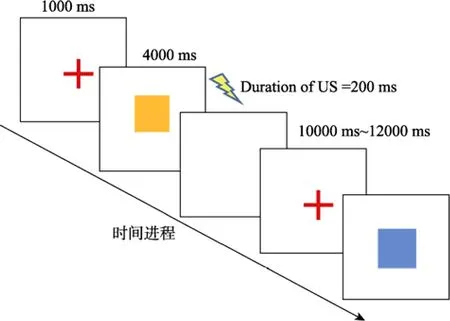
图1 实验流程示意图
第一天条件恐惧习得阶段:CS+呈现16次, CS−呈现10次。38% (6次)的CS+后跟随US, 而 CS−后不跟随任何刺激, 3组被试的习得阶段完全相同。CS+和 CS−均为复合刺激(声音+图片); US为微弱电击, 时间持续200 ms。
第二天提取−消退阶段:“声音提取−消退”组(T组)先单独呈现一次声音(前一天 CS+中的声音线索)以提取激活习得的恐惧记忆, 被试休息 10 min后,进行消退训练, CS+和CS−的声音线索各呈现10次,其后不跟随任何刺激; “图片提取−消退”组(P组)先单独呈现一次图片(前一天 CS+中的图片线索), 被试休息10 min后, 进行消退训练, CS+和CS−的图片线索各呈现 10次, 其后不跟随任何刺激; “复合线索提取−消退”组(T+P组)先单独呈现一次复合刺激线索(前一天CS+), 被试休息10 min后, 进行消退训练, CS+和CS− (第一天复合刺激)各呈现10次,其后不跟随任何刺激。
第三天自发恢复−重建测试阶段:先进行自发恢复测试, CS+和CS− (第一天复合刺激)各呈现10次, 其后不跟随任何刺激; 测试结束 2 min后, 向被试连续实施4次电击, 每次电击持续时间200 ms,电击间隔500 ms; 被试休息10 min后, 进行重建测试, CS+和CS− (第一天复合刺激)各呈现10次, 其后不跟随任何刺激。具体设计见表1。

表1 实验设计
3 实验结果
对收集的各个被试的皮电数据, 以刺激类型(CS+ / CS−)和试次为被试内因素, 组别(T组、P组、T+P组)为被试间因素做重复测量方差分析。
第一天恐惧习得阶段:重复测量方差分析结果显示, 刺激类型的主效应显著,F(1,34)=18.49,p<0.01, 偏η2=0.54; 试次的主效应显著,F(9,306)=2.62,p< 0.001, 偏η2=0.08; 刺激类型和试次的交互效应不显著,F(9,306)=1.16,p=0.55。组别效应不显著,F(2,34)=0.024,p=0.98; 刺激类型和组别的交互作用也不显著,F(2,34)=2.43,p=0.13, 说明3个组被试对CS+和CS−的恐惧反应不存在显著差异, 如图2所示。
第二天提取−消退阶段:在提取阶段, 无论提取−消退线索是声音(T组), 图片(P组)还是复合刺激(T+P组)都能成功的诱发被试的恐惧反应, 并且3组之间皮电值没有显著差异,F(2,34)=0.71,p=0.50, 说明提取是成功的。

图2 习得、消退阶段各组皮电SCR值的变化
消退阶段的重复测量方差分析结果显示:刺激类型的主效应显著,F(1,34)=10.74,p=0.002, 偏η2=0.24; 试次的主效应显著,F(9,306)=18.34,p<0.001, 偏η2=0.35; 刺激类型和试次的交互效应显著,F(9,306)=2.13,p<0.05, 偏η2=0.06, 即CS+和CS−之间 SCR的差异在消退阶段后期小于消退阶段开始时, 说明被试进行了成功的消退, 如图2所示。组别效应不显著,F(2,34)=0.10,p=0.91; 刺激类型和组别的交互作用也不显著,F(2,34)=1.90,p=0.17, 说明3组被试在消退过程中不存在显著性差异。
第三天自发恢复测试阶段:恐惧反应的自发恢复测试, 比较的是第三天再消退阶段的第一个试次CS引发的SCR值和第二天消退的最后一个试次的SCR值。重复测量方差分析结果显示:刺激类型的主效应显著,F(1,34)=13.69,p< 0.01, 偏η2=0.29;阶段的主效应显著,F(1,34)=36.78,p< 0.001, 偏η2=0.52; 刺激类型和阶段的交互效应显著,F(1,34)=9.51,p< 0.05, 偏η2=0.22; 事后比较发现, 被试只在CS+水平上出现自发恢复, 在CS−水平上没有出现自发恢复。组别效应边缘显著,F(2,34)=3.66,p< 0.05,偏 η2=0.18; 刺激类型和组别的交互作用不显著,F(2,34)=1.20,p=0.31。分离刺激类型进行进一步的分析发现, 在 CS+水平上:组别与阶段交互作用显著,F(2,34)=26.54,p< 0.001, 偏η2=0.61, 说明3组在 CS+水平上的自发恢复存在显著差异; 再分别对每个组在不同阶段上的皮电值进行配对样本t检验,结果显示, T组在两阶段上的皮电值差异显著,t(11)=–1.78,p< 0.001; P 组差异不显著,t(11)=–1.78,p=0.10; T+P 组差异不显著,t(12)=–0.50,p=0.63。说明T组出现了自发恢复, 而P组, T+P组没有出现自发恢复, 如图3所示。为了验证3组在CS+水平上的恐惧反应差异, 将每个被试在第三天再消退阶段的第一个试次和第二天消退阶段的最后一个试次相减, 再将 3组被试的 SCR差值做单因素方差分析,结果显示, 3组被试的 SCR差值存在显著差异,F(2,34)=26.54,p< 0.01, T组的SCR差值显著大于P组和T+P组, 如图4所示。

图3 再消退阶段各组皮电SCR值的变化

图4 各组自发恢复测试−SCR差值
第三天重建测试阶段:恐惧反应的重建测试,比较的是重建(单独呈现4次电击)后的第一个试次CS引发的SCR值和再消退的最后一个试次的SCR值。重复测量方差分析结果显示:刺激类型的主效应显著,F(1,34)=15.91,p< 0.001, 偏 η2=0.32; 阶段的主效应显著,F(1,34)=44.35,p< 0.001, 偏η2=0.57; 刺激类型和试次的交互效应不显著,F(1,34)=3.19,p=0.83。组别效应显著,F(2,34)=4.29,p<0.05, 偏η2=0.20; 刺激类型和组别的交互作用显著,F(2,34)=6.70,p< 0.01, 偏η2=0.33。分离刺激类型进行进一步的分析发现, 在CS+水平上:组别与阶段交互作用显著,F(2,34)=9.60,p <0.001, 偏η2=0.36, 说明3组在重建后, 由CS+引发的恐惧反应存在显著差异; 再分别对每个组在不同阶段上的皮电值进行配对样本T检验, 结果显示, T组在两阶段上的皮电值差异显著,t(11)=–4.28,p< 0.01;P组差异也显著,t(11)=–3.18,p< 0.01; T+P组在两阶段上的皮电值差异不显著,t(12)=0.29,p=0.78,说明T组和P组出现了重建效应, 而T+P组没有出现重建效应, 如图5所示。进一步比较3组在CS+水平上的恐惧反应差异, 将每个被试在重建后的第一个试次和再消退阶段的最后一个试次相减, 再将3组被试的SCR差值做单因素方差分析, 结果显示,3组被试的SCR差值存在显著差异,F(2,34)=9.60,p< 0.01。事后比较发现, T+P组的SCR差值显著小于T组和P组, 且P组的SCR差值也显著小于T组, 即T+P组 本研究采用多感官复合刺激(声音+图片)作为条件刺激, 考察采用单个线索(声音或图片)、复合刺激线索(声音+图片)进行提取−消退对条件性恐惧记忆的消退的影响。研究发现, 在第一天的习得和第二天的消退阶段, T组(声音提取−消退组)、P组(图片提取−消退组)、T+P组(复合刺激提取−消退组)的SCR值均无显著差异, 3组成功进行了习得和消退。在第三天的自发恢复测试中, T组出现了自发恢复, 而P组, T+P组没有出现, 在重建测试阶段,T组和P组均出现重建效应, 而T+P组没有出现重建效应。 图5 重建后各组皮电SCR值的变化 实验中, T组被试同时出现恐惧的自发恢复和重建效应, P组出现重建效应, T+P组既没有出现自发恢复, 也没有出现重建, 即未出现恐惧返回的现象, 表明完整的复合刺激线索的提取−消退效果优于单个线索。可能原因是, 在复合刺激 CS形成的条件性恐惧中, 不仅复合刺激CS与US形成了联结,复合刺激中的声音和图片也形成了联结(Durlach &Rescorla, 1980; Rescorla & Cunningham, 1978), 两种联结相对独立, 用复合刺激线索进行提取−消退可以同时破坏这两种联结, 使恐惧的消退效果更好。 单线索提取−消退组中, 声音提取−消退组, 同时出现了自发恢复和重建, 消退返回明显, 图片提取−消退组, 仅出现重建, 图片提取−消退的效果好于声音提取−消退。原因可能是相对声音刺激, 图片刺激比较强势。心理学家Treicher通过大量的实验证实, 人类获取的信息 83%来自视觉, 11%来自听觉。对于我们人类被试来说, 对视觉的敏感性明显强于听觉, 即倾向于把图片当做强线索(恐惧线索), 把声音当做弱线索(背景或情境线索), 因此在本研究中对图片的提取−消退效果比声音更好, 这也提示由复合刺激线索引发的条件性恐惧, 采用复合刺激中的单个较强线索进行提取−消退, 能更有效地消退被试的恐惧反应, 抑制恐惧的自发恢复和重建等恐惧复发行为。但是Jones等人对动物的研究发现, 大鼠对复合刺激中的声音线索的提取−消退效果强于灯光线索, 研究者认为大鼠的对听觉的敏感性强于视觉, 并倾向于把强度较弱的灯光当做背景(情境)线索, 而把强度较强的声音当做恐惧线索(Jones, Ringuet, & Monfils, 2013)。这也许就是人和动物的区别之一, 对于老鼠而言, 声音是强势刺激。 图6 各组重建测试-SCR差值 本研究发现, T组和P组在利用声音或图片进行提取−消退后, 两组被试均出现了重建的恐惧返回现象, 说明利用单一线索的提取−消退, 并不能完全减弱由复合刺激引起的恐惧反应, 消除条件性恐惧记忆。Dębiec, Díaz-Mataix, Bush, Doyère 和LeDoux (2010)的研究发现, 在大鼠习得复合刺激CS (灯光+声音)的条件性恐惧后的第二天, 一组大鼠使用灯光激活提取恐惧记忆后, 注射蛋白质合成抑制剂破坏再巩固, 另一组不进行提取, 直接使用灯光进行消退训练, 在第三天的长时记忆测试中,两组大鼠对灯光、声音的恐惧反应无显著差异, 也就是说, 单线索提取出现了消除复合刺激中另一个刺激的恐惧效果(Dębiec et al., 2010), 但研究者并未测试大鼠对原有复合刺激 CS的恐惧反应, 也没有进行复合刺激的提取−消退比较。 在本研究中, P组没有出现自发恢复, 但出现了重建, 且自发恢复测试P组的SCR值与T+P组无显著差异, 但重建测试时显著高于T+P组, P组的SCR值在自发恢复和重建两个指标上出现分离,即利用图片进行提取−消退能抑制自发恢复但不影响重建。这与Trent, Barnes, Hall和Thomas (2015)的研究结果是一致的, 研究者认为, 自发恢复和重建这两个指标都是用来衡量恐惧记忆返回或复发的情况, 且两个指标的方向基本是一致的, 但这两个指标的操作定义和神经机制具有本质的差别。在第三天单独呈现US后, 重建测试再次出现CS, 可激活 CS-US的联结记忆, 再次出现恐惧反应(Trent et al., 2015; Clem & Schiller, 2016)。 本研究在前人研究的基础上, 以人做被试, 将提取−消退范式应用于更复杂的条件性恐惧记忆,证明了利用单一线索的提取−消退, 并不能完全减弱由复合刺激引起的恐惧反应, 消除条件性恐惧记忆复合刺激线索引发的条件性恐惧, 也同时证明了采用复合刺激中的较强线索进行提取−消退, 恐惧消退的效果更好。但本研究尚不能明确在联结学习过程中刺激是如何进行编码的, 声音和图片这两个刺激线索是作为一个混合刺激进行储存, 还是作为与同一个US相连的不同的但相关的两个刺激储存;在提取激活时, 单一线索是激活了一种成分或者是两个成分都被激活, 这些方面仍需进一步的研究。 Auber, A., Tedesco, V., Jones, C.E., Monfils, M.H., &Chiamulera, C.(2013).Post-retrieval extinction as reconsolidation interference: Methodological issues or boundary conditions?.Psychopharmacology, 226(4), 631−647. Bradley, R., Greene, J., Russ, E., Dutra, L., & Westen, D.(2005).A multidimensional meta-analysis of psychotherapy for PTSD.American Journal of Psychiatry, 162(2), 214−227. Clem, R.L., & Schiller, D.(2016).New learning and unlearning: Strangers or accomplices in threat memory attenuation?.Trends in Neurosciences, 39(5), 340−351. Davis, M., Walker, D.L., & Myers, K.M.(2003).Role of the amygdala in fear extinction measured with potentiated startle.Annals of the New York Academy of Sciences, 985,218−232. Dębiec, J., Díaz-Mataix, L., Bush, D.E.A., Doyère, V., &LeDoux, J.E.(2010).The amygdala encodes specific sensory features of an aversive reinforcer.Nature Neuroscience,13(5), 536−537. Durlach, P.J., & Rescorla, R.A.(1980).Potentiation rather than overshadowing in flavor-aversion learning: An analysis in terms of within-compound associations.Journal of Experimental Psychology: Animal Behavior Processes,6(2), 175−187. Jones, C.E., Ringuet, S., & Monfils, M.H.(2013).Learned together, extinguished apart: Reducing fear to complex stimuli.Learning & Memory, 20(12), 674−685. Monfils, M.H., Cowansage, K.K., Klann, E., & LeDoux, J.E.(2009).Extinction-reconsolidation boundaries: Key to persistent attenuation of fear memories.Science, 324(5929),951−955. Myers, K.M., & Davis, M.(2002).Behavioral and neural analysis of extinction.Neuron, 36(4), 567−584. Myers, K., & Davis, M.(2007).Mechanisms of fear extinction.Molecular Psychiatry, 12(2), 120−150. Nader, K., Schafe, G.E., & LeDoux, J.E.(2000).The labile nature of consolidation theory.Nature Reviews Neuroscience, 1(3),216−219. Rescorla, R.A.(2000).Extinction can be enhanced by a concurrent excitor.Journal of Experimental Psychology:Animal Behavior Processes, 26(3), 251−260. Rescorla, R.A., & Cunningham, C.L.(1978).Recovery of the US representation over time during extinction.Learning &Motivation, 9(4), 373−391. Schiller, D., Monfils, M.H., Raio, C.M., Johnson, D.C.,Ledoux, J.E., & Phelps, E.A.(2010).Preventing the return of fear in humans using reconsolidation update mechanisms.Nature, 463(7277), 49−53. Trent, S., Barnes, P., Hall, J., & Thomas, K.L.(2015).Rescue of long-term memory after reconsolidation blockade.Nature Communications, 6, 7897. Walker, M.P., Brakefield, T., Hobson, J.A., & Stickgold, R.(2003).Dissociable stages of human memory consolidation and reconsolidation.Nature, 425(6958), 616−620. Witnauer, J.E., & Miller, R.R.(2012).Associative status of the training context determines the effectiveness of compound extinction.Journal of Experimental Psychology:Animal Behavior Processes, 38(1), 52−65.4 讨论


猜你喜欢
意林(2021年9期)2021-05-28
今传媒(2019年10期)2019-11-11
中华建设(2019年8期)2019-09-25
银幕内外(2019年4期)2019-09-10
当代陕西(2019年12期)2019-07-12
现代经济信息(2019年7期)2019-06-23
读友·少年文学(清雅版)(2018年2期)2018-09-10
发明与创新·中学生(2018年8期)2018-09-08
祝您健康(2018年9期)2018-09-04
科学生活(2015年3期)2015-01-03
