
外源激素和温度对切花菊侧芽萌发与内源激素含量的影响
2018-03-07 06:50王小乐房伟民陈发棣管志勇姚建军薛建平
江苏农业学报 2018年1期
赵 凤, 王小乐, 房伟民, 张 飞, 陈发棣, 管志勇, 姚建军, 薛建平
(1.南京农业大学园艺学院,江苏 南京 210095; 2.上海虹华园艺有限公司,上海 200070)
菊花(Chrysanthemummorifolium)是中国十大传统名花和世界四大切花之一,也是中国主要出口鲜切花,其中单头标准菊是其主要类型[1]。标准切花菊生产需要大量人工进行侧芽侧蕾的抹除工作,而少侧芽标准切花菊品种的选育与应用可减少该环节工作,节省大量人力物力。但少侧芽品种在繁育阶段也面临母本萌芽少、采穗率低等问题,在高温阶段尤其突出,难以通过扦插的方式大量繁育种苗。因此,研究少侧芽标准切花菊品种的侧芽萌发和生长的调控技术及其机理具有重要的实践和理论意义。
植株侧枝的生长发育与其自身部位、激素和温度有关,如甘蔗中不同部位植株侧芽萌发有显著的差异[2-3]。而植物激素对侧枝的发育有重要影响,其中,IAA和6-BA是在生产上应用比较广泛的植物生长调节剂,如外源生长素IAA可以显著抑制水稻分蘖[4]和杨树枝条的形成[5],而6-BA能促进梨枝条的发育[6],也影响苹果幼苗的分枝[7-8]。此外,植株侧枝发育还与温度有关,如相关实践和李俊香等研究结果均表明,随着温度升高,菊花的侧芽生长加快,但在高温下会受到明显抑制[9]。目前,植物生长调节剂促进分枝的应用主要集中在果树和大田作物等方面,在菊花的生产上未见报道。切花菊精の一世属于少侧芽标准型切花菊品种,近几年得到了市场的认可并在生产中大量应用,但育苗时也面临侧芽少、采穗率低的问题,在一定程度上限制了它的大量扩繁应用。
本研究主要利用精の一世组培苗,探究不同部位的侧芽发生特点及生长素、细胞分裂素和温度对精の一世侧芽发生与生长的影响,为了解其侧芽萌发生长的调控机制,进而为提高少侧芽菊花品种的繁殖效率提供技术和理论指导。
1 材料与方法
1.1 试验材料
供试材料为标准切花菊品种精の一世无菌苗,来源于南京农业大学中国菊花种质资源保存中心,该品种侧芽萌发量较少。试验所用的试剂6-苄氨基腺嘌呤(6-BA)、萘乙酸(NAA)为化学纯,冰醋酸(CH3COOH)为分析纯,甲醇(CH3OH)为色谱纯。ZT(玉米素)、GA(赤霉素)、IAA(3-吲哚乙酸)、ABA(脱落酸)为色谱纯。
1.2 试验方法
1.2.1 不同部位的离体茎段侧芽萌发特点 取生长旺盛的组培苗,以1LB(第一节位)为靠近顶芽向下数的第一片叶子所在的节位,以此类推将其分成上部(1LB和2LB)、中部(3LB和4LB)和下部(5LB和6LB),分别接种到MS培养基中, 3次重复,每个重复5瓶,每瓶接种3~4个茎段,常温下培养。分别在第7 d、第14 d、第21 d、第35 d统计组培苗不同部位侧芽萌发(展开2片成熟叶)的数目,并计算萌芽率。同时测定35 d时侧芽长度、长枝(长度大于等于1 cm)数目,并计算长枝率(长枝数目除以萌芽数目)。
内源激素测定:35 d时,取不同位置侧芽, 液氮速冻,-80 ℃保存备用。每次取样9株,3次重复。超高效液相色谱(UPLC)法同时测定样品中的IAA、ZT、GA和ABA含量,测定方法参照侯凯等的方法[10]。
1.2.2 6-BA和NAA对离体茎段侧芽萌发的影响 取生长旺盛的组培苗中部节位的茎段,在MS培养基中分别添加0.1 mg/L、0.5 mg/L、1.0 mg/L的NAA和0.5 mg/L、1.0 mg/L、2.0 的6-BA,MS培养基为对照,常温下培养。28 d后测定茎段侧芽萌发数、畸形(侧芽矮化)数、侧芽长度、长枝数并计算萌芽率(萌芽数目除以节位数目)、畸形率(畸形芽数目除以节位数目)和长枝率,并取样测定IAA、ZT、GA和ABA含量,测定方法同方法1.2.1。
1.2.3 不同温度对离体茎段侧芽萌发的影响 取生长旺盛的组培苗中部节位的茎段接种到MS培养基,设4个温度处理,分别为昼/夜温25 ℃/20 ℃、22 ℃/17 ℃、28 ℃/ 23 ℃、30 ℃/ 25 ℃,其中25 ℃/20 ℃为对照,光周期为光照16 h/黑暗8 h,光照度为3 000 lx。每处理3次重复,每重复3瓶,每瓶接3个茎段。在处理后第14 d、第21 d、第35 d时分别跟踪调查茎段萌芽数,并计算萌芽率,在35 d时测定侧芽长度、长枝数目,计算长枝率,并测定IAA、ZT、GA和ABA含量,测定方法同方法1.2.1。
1.3 数据与分析
应用SPASS20.0和Microsoft Excel 2007软件进行数据统计与分析。
2 结果与分析
2.1 精の一世不同部位侧芽萌发特点及其内源激素含量差异
表1表明,精の一世侧芽的萌发速度和生长状况与其部位有关,中部茎段侧芽萌发速度比上部和下部快,其中在接种7 d时,中部侧芽的萌芽率达36%,上部为18%,下部为10%;14 d时,中部侧芽萌发率也最高,为70%;35 d时,上部、中部、下部的侧芽萌发率均达到100%。而35 d时中部和下部的侧芽长度分别为2.42 cm和2.32 cm,均显著高于上部的1.32 cm;长枝率从上部至下部逐步提高,下部侧芽长枝率最高,达98%,上部侧芽的长枝率最低,仅为56%,说明中部和下部侧芽的质量较上部好。
表1精の一世不同部位侧芽萌发
Table1Axillarybuddingsatdifferentpartofinvitrostemof‘SeiNoIssei’
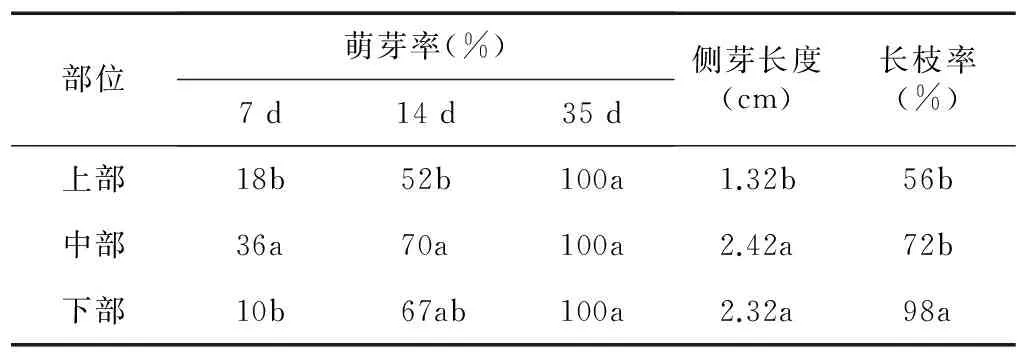
部位萌芽率(%)7d14d35d侧芽长度(cm)长枝率(%)上部18b52b100a1.32b56b中部36a70a100a2.42a72b下部10b67ab100a2.32a98a
同一列数据后不同小写字母表示差异显著(P<0.05)。
图1表明,不同部位的内源激素含量有显著差异,上部的IAA和ABA含量最高,分别达到860.58 ng/g和639.49 ng/g,均显著高于中部、下部。而不同部位的ZT和GA的含量也均有显著差异,下部最高、中部最低。图2表明,不同部位ZT/ABA、GA/ABA、IAA/ABA、ZT/IAA和(GA+IAA+ZT)/ABA均有显著差异,中部ZT/IAA最高,为0.36,下部次之,为0.21,上部最低,仅为0.11,因此中部茎段最易萌发,而中、下部ZT/ABA、GA/ABA、IAA/ABA、ZT/IAA和(GA+IAA+ZT)/ABA均高于上部,也印证了中部、下部侧芽萌发率、侧芽长度和长枝率均高于上部的现象。
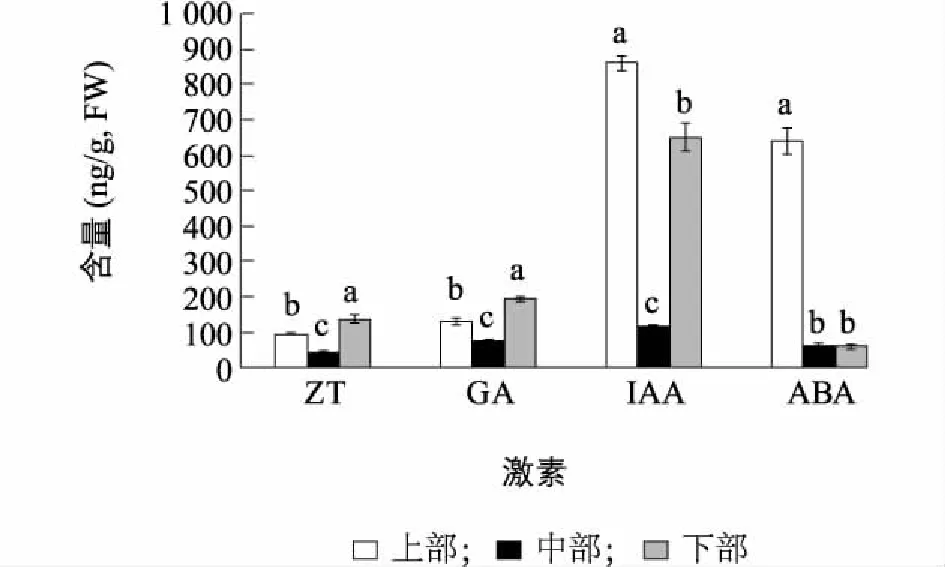
不同小写字母表示差异显著(P<0.05)。图1 精の一世不同部位侧芽的内源ZT、GA、IAA、ABA含量Fig.1 The levels of endogenous ZT, GA, IAA and ABA in axillary buds at different part of ‘Sei No Issi’
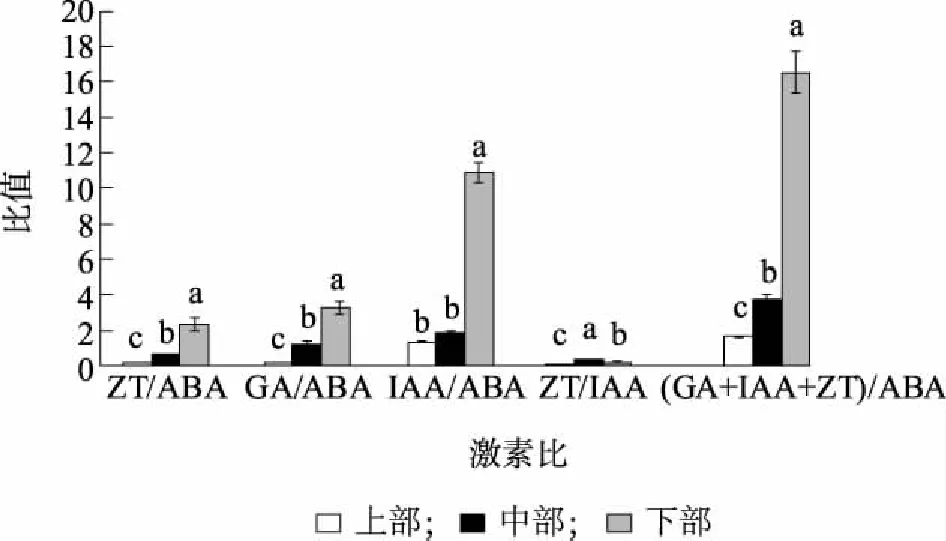
不同小写字母表示差异显著(P<0.05)。图2 精の一世不同部位侧芽的内源激素比例Fig.2 Ratio of different endogenous hormones in axillary buds at different part of in vitro stem of cut chrysanthemum ‘Sei No Issi’
2.2 不同浓度的NAA和6-BA对侧芽萌发的影响及对内源激素的影响
表2表明,随着NAA浓度的增加,精の一世侧芽的萌发率降低,由83%下降到30%;各处理的侧芽长度也显著低于对照,由1.28 cm减少到0.45 cm;侧芽的长枝率也降低,由对照的50%下降到20%。
表2不同浓度NAA处理对精の一世侧芽萌发的影响
Table2EffectofdifferentconcentrationsofNAAonaxillarybuddingsof‘SeiNoIssei’stem

NAA浓度(mg/L)萌芽率(%)畸形率(%)侧芽长度(cm)长枝率(%)083a11ab1.28a50a0.179a21a0.69bc26b0.555b0b0.91b35b1.030c0b0.45c20c
同一列数据后不同小写字母表示差异显著(P<0.05)。
表3表明, 0.5 mg/L和1.0 mg/L 6-BA均促进了侧芽的萌发, 2.0 mg/L 6-BA则抑制了侧芽萌发。此外,1.0 mg/L和2.0 mg/L 6-BA处理的侧芽畸形率均有明显提高,最高达到70%。可见,6-BA明显抑制了侧芽的生长,各处理的侧芽长度显著低于对照,长枝率也随之明显下降。
表3不同浓度6-BA处理对精の一世侧芽萌发的影响
Table3Effectofdifferentconcentrationsof6-BAonaxillarybuddingsof‘SeiNoIssei’
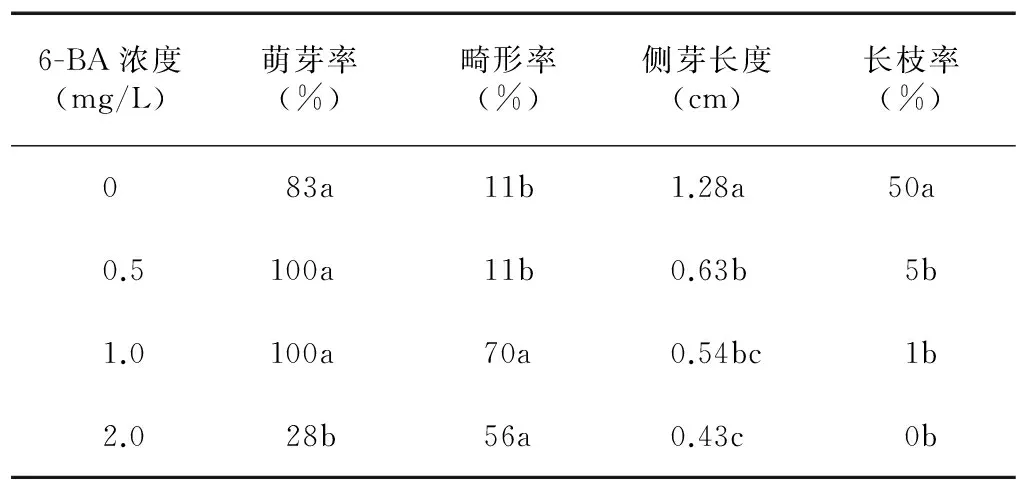
6-BA浓度(mg/L)萌芽率(%)畸形率(%)侧芽长度(cm)长枝率(%)083a11b1.28a50a0.5100a11b0.63b5b1.0100a70a0.54bc1b2.028b56a0.43c0b
同一列数据后不同小写字母表示差异显著(P<0.05)。
图3表明,外源NAA会引起植物体内ZT含量显著下降,由89.9 ng/g下降到12.5 ng/g,减少了86.1%;IAA含量显著上升,高达656.4 ng/g,增加了45.8%;而GA和ABA含量没有显著变化。而外源6-BA主要导致ZT含量显著升高,达121.2 ng/g;GA和IAA含量显著下降,GA仅为50.6 ng/g,下降了72.1%,IAA仅为273.6 ng/g,下降了39.2%。
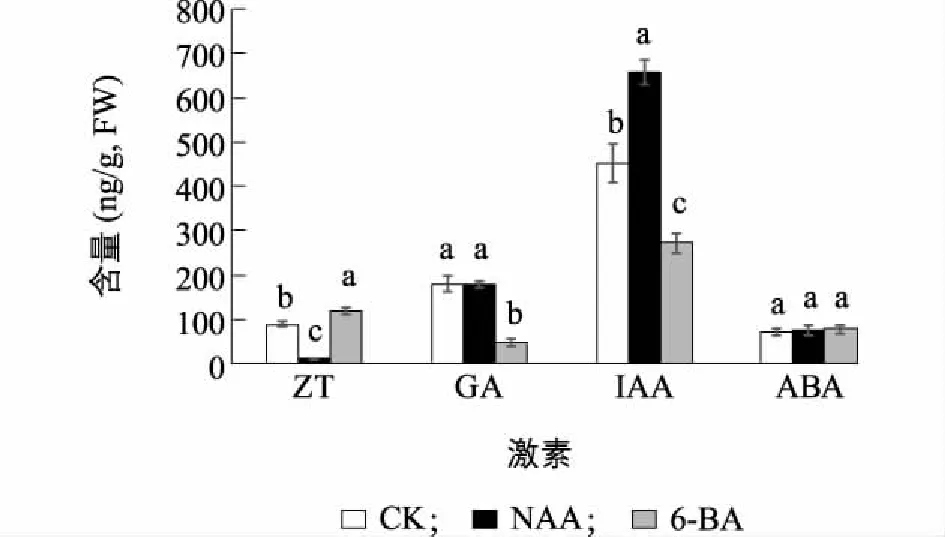
不同小写字母表示差异显著(P<0.05)。图3 NAA和6-BA处理对精の一世侧芽内源激素含量的影响Fig.3 Effects of NAA and 6-BA on levels of four endogenous hormones in ‘Sei No Issei’
图4表明,外源NAA对内源激素的比例也产生了影响,生长类激素与抑制类激素(GA+IAA+ZT)/ABA比例提高,达到11.1,而生长类激素相对含量的上升可以显著促进侧芽生长;外源NAA处理引起ZT/ABA和ZT/IAA显著降低,与对照相比ZT/ABA下降了87.3%,ZT/IAA下降了90.5%,而IAA/ABA则有显著提高,GA/ABA没有显著变化。而外源6-BA处理后显著提高了ZT/IAA,较对照增加了120%,ZT/ABA没有显著变化,而GA/ABA、IAA/ABA均有显著下降。
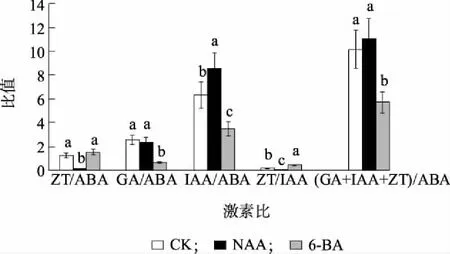
不同小写字母表示差异显著(P<0.05)。图4 NAA和6-BA处理对精の一世侧芽四种内源激素比例的影响Fig.4 Effects of NAA and 6-BA on ratio of four endogenous hormones in ‘Sei No Issei’
2.3 不同温度对茎段侧芽萌发生长及内源激素含量的影响
表4表明,温度的升高会显著抑制精の一世离体茎段侧芽的萌发和生长。22 ℃日温时,茎段在35 d的萌发率达100%,但是侧芽长度比25 ℃显著降低。在超过25 ℃日温后,随着温度的升高,侧芽的萌发率、侧芽长度、长枝率均逐渐降低。而在30 ℃日温培养条件下,茎段在14 d、21 d、35 d时萌发率、侧芽长度和长枝率都下降到最低,仅分别为55%、58%、67%,说明高温显著抑制精の一世侧芽的萌发和生长。
表4不同温度处理对精の一世侧芽萌发的影响
Table4Effectsoftemperaturesontheaxillarybuddingsof‘SeiNoIssei’

温度(℃)萌芽率(%)14d21d35d侧芽长度(cm)长枝率(%)2268b92a100a1.65b88a2583a92a100a2.08a92a2871b75b79b1.88a89a3055c58c67c1.37c77b
同一列数据后不同小写字母表示差异显著(P<0.05)。
图5表明,与25 ℃对照相比,30 ℃高温使侧芽ZT极显著降低,仅为219.2 ng/g,FW,降低了80.9%;而GA和ABA极显著升高,GA增加了1.21倍,达367 ng/g,FW,ABA增加了1.75倍,达603.5 ng/g,FW,IAA则没有显著变化。

*和**分別表示在0.05和0.01水平差异显著。图5 不同温度处理对精の一世侧芽四种内源激素含量的影响Fig.5 Effects of different temperatures on the level of four endogenous hormones in ‘Sei No Issei’
图6表明,高温使ZT/ABA、IAA/ABA以及ZT/IAA极显著降低,从而使生长促进类激素和生长抑制类激素比值(GA+IAA+ZT)/ABA极显著降低,由5.03降低到1.29,而GA/ABA没有显著变化。生长促进类激素和生长抑制类激素的比例极显著降低是引起30 ℃高温抑制侧芽萌发和生长的内在原因。

*和**分別表示在0.05和0.01水平差异显著。图6 不同温度处理对精の一世侧芽四种内源激素比例的影响Fig.6 Effects of different temperatures on the ratio of four endogenous hormones in ‘Sei No Issei’
3 讨 论
植株侧芽的萌发与生长受到侧芽中内源激素含量和激素间比例的变化调控,而侧芽的不同部位、外源激素和外界温度等均影响内源激素的变化。
激素是调控侧芽萌发的主要因素,抑制生长素的极性运输,能促进植株侧芽的生长[11-12],而CTK(细胞分裂素)能促进侧芽生长,从而抑制植株侧芽内IAA的合成及向外运输[13-14],赤霉素则能协同细胞分裂素共同促进植株侧芽的生长[15]。高贵宾等[16]对美丽箬竹盆栽苗地下茎侧芽萌发特征研究发现,IAA、GA对侧芽发笋影响比较大。本试验中,上部侧芽IAA和ABA含量均显著高于中下部,且中部ZT/IAA最高,下部次之,上部最低,较高的ZT/IAA比值会促进侧芽的萌发生长,这从精の一世离体茎段上部侧芽的萌发与伸长受到抑制,中部的萌发率和长枝率均高于上部的结果也得到了印证,这与肖关丽等[2]在甘蔗离体茎段的研究一致。
施用外源IAA和6-BA会对植株侧枝的生长发育产生影响。刘凯等[5]用0.01 mg/L IAA喷施杨树,可以明显抑制杨树枝条的形成;张庆伟等[7]发现喷施500 mg/L 6-BA能有效促进苹果幼苗分枝。本研究发现,一定浓度的外源生长素NAA抑制了精の一世侧芽的萌发生长, 0.5 mg/L 6-BA则促进侧芽的萌发,而1.0 mg/L 6-BA虽能促进侧芽萌发生长,但侧芽畸形率增加。外源激素会引起植株体内内源激素含量发生变化,从而影响植株的分枝发育,并且影响侧芽与侧枝的形成[17-18]。Kalousek等[19]发现细胞分裂素对生长素的合成与运输有重要影响,从而通过生长素的合成与运输来调控侧芽的萌发生长;刘杨等[20]认为,外源GA3和NAA提高了分蘖节中ABA的含量从而抑制分蘖节的生长;蔡铁等[21]认为外源激素主要通过影响内源IAA、ZT含量以及IAA/ZT和ABA/ZT来影响分蘖芽的生长进而调控小麦分蘖的发生。本试验发现精の一世侧芽的形成并不是受外源单一某种激素的影响,而是受多种激素相互作用的影响,外源NAA处理引起ZT/ABA和ZT/IAA显著降低,ZT/ABA降低则会抑制侧芽的发生,这与刘杨[20]、蔡铁[21]的研究结果一致;外源6-BA处理菊花试管苗中ZT/IAA上升,GA/ABA、IAA/ABA显著下降,而ZT/IAA的上升和GA/ABA的下降则会导致侧芽的发生与生长,这与Miguel等[17]、Kalousek等[19]和黄坚钦等[22]的研究结果一致。
精の一世在常温下侧芽正常萌发生长,30 ℃高温抑制了侧芽的萌发生长,表明精の一世侧芽的萌发生长与外部环境温度相关。这与李俊香等[9]发现随着温度升高,深志的侧芽在高温33 ℃下会受到明显抑制的结果相同,Schoellhorn 等[23]、Huh等[24]在菊花上的研究结果也类似。
温度会引起植株体内内源激素含量发生变化,从而影响植株的分枝发育。李俊香等[9]研究结果表明,温度可能通过影响深志茎尖中ZR含量及IAA/ZR值调控其侧芽的形成[9]。本试验中,对内源激素含量的测定发现,30 ℃高温下侧芽中ZT的含量极显著降低,GA和ABA含量极显著升高,引起ZT/IAA显著降低。由此推测,温度主要是通过影响内源细胞分裂素与生长素的含量与比例影响侧芽的萌发生长。同时本研究还发现生长促进类激素和生长抑制类激素比例(GA+IAA+ZT)/ABA的降低抑制了侧芽的萌发生长,表明侧芽的萌发生长可能是几种激素共同调控的结果。
综上所述,选择植株中部或下部茎段、添加0.5 mg/L 6-BA,25 ℃左右的培养温度有利于精の一世离体侧芽萌发和生长。当然,由于菊花侧芽的萌发与生长受到多种因素的影响,各种因素复合处理对侧芽萌发生长的影响还需要进一步的研究。
[1] 陈俊愉. 园林植物品种分类学[M]. 北京: 高等教育出版社,2001.
[2] 肖关丽,杨清辉,李富生,等. 甘蔗腋芽液体培养萌发与内源激素关系[J]. 西南农业大学学报,2001,23(6): 538-541.
[3] DOMAGALSKA M A,LEYSER O. Signal integration in the control of shoot branching [J]. Nature Reviews Molecular Cell Biology,2011,12: 211-221.
[4] 蔡 铁,徐海成,尹燕枰,等. 外源IAA、GA3和ABA影响不同穗型小麦分蘖发生的机制 [J]. 作物学报,2013,39(10): 1835-1842.
[5] 刘 凯,余 敏,陈海燕,等. 外源IAA和GA3对杨树应拉木形成的影响[J]. 西北农林科技大学学报(自然科学版),2016,44(3):125-132.
[6] 郑传亮,张 琦. 6-BA 和 GA对新梨七号芽体抽枝能力的影响[J]. 新疆林业,2013(6):21-22.
[7] 张庆伟,宋春晖,刑利博. 6-BA和 GA4+7 喷施处理及其他措施促进长富 2 号苹果幼苗分枝的效果[J].果树学报,2011,28 (6): 1071-1076.
[8] 王东洪. 不同种源山桐子冬芽休眠的温度特性研究[D].郑州:河南农业大学,2012.
[9] 李俊香,温 超,刘凤栾,等. 温度对切花菊‘深志’侧芽形成的影响[J]. 中国农业大学学报,2014,19(1): 74-79.
[10] 侯 凯,陈郡雯,申 浩,等. 川白芷内源激素的提取纯化和高效液相色谱法同步测定 [J]. 核农学报,2013,27(5): 653-657.
[11] STIRNBERG P,CHATFIELD S P,LEYSER H M O.AXRIbud growth inArabidopsisacts after lateral bud formation to inhibit lateral bud growth inArabidopsis[J]. Plant Physiology,1999,121:839-847.
[12] REDDY S K,FINLAYSON S A. Phytochrome B promotes branching inArabidopsisby suppressing auxin signaling [J]. Plant Physiology,2014,164: 1542-1550.
[13] 李春俭. 植物激素在顶端优势中的作用 [J]. 植物生理学通讯,1995,31(6): 401-406.
[14] 李春俭. 乙烯在豌豆植株顶端优势中的作用 [J]. 植物生理学报,1997,23(3): 283-287.
[15] NI J,GAO C C,CHEN M S, et al. Gibberellin promotes shoot branching in the perennial woody [J]. Plant Cell and Physiology,2015,56(8): 1655-1666.
[16] 高贵宾,潘雁红,吴志庄,等. 美丽箬竹盆栽苗地下茎侧芽萌发特征研究 [J]. 植物科学学报,2016,34(3): 460-468.
[17] MIGUEL L C,LONGNECKER N E,MA Q,et al. Branch development inLupinusangustifoliusL. I. Not all branches have the same potential growth rate [J]. Journal of Experimental Botany,1998,49(320):547-553.
[18] EMERY R J N,LONGNECKER N E,ATKINS C A. Branch development inLupinusangustifoliusL. II. Relationship with endogenous ABA,IAA and cytokinins in axillary and main stem buds [J]. Journal of Experimental Botany,1998,49(320):555-562.
[19] KALOUSEK P,BUCHTOVá D,BALLA J,et al. Cytokinins and polar transport of auxin in axillary pea buds [J]. Acta Universitatis Agricuturae Et Silviculturae Mendelianae Brunensis,2010,54(4):79-87.
[20] 刘 杨,丁艳锋,王强盛,等. 植物生长调节剂对水稻分蘖芽生长和内源激素变化的调控效应[J]. 作物学报,2011,37(4): 670-676.
[21] 蔡 铁,徐海成,尹燕枰,等. 外源IAA、GA3和ABA影响不同穗型小麦分蘖发生的机制 [J]. 作物学报,2013,39(10): 1835-1842.
[22] 黄坚钦,刘 力,章滨森,等. 雷竹地下鞭侧芽内源激素的动态变化研究 [J]. 林业科学,2002,38(3): 38-41.
[23] SCHOELLHORN R K,BARRETT J E,BARTUSKA C A,et al. Elevated temperature affects axillary meristem development inDerzdrarzthema×grandiflorum‘Improved Mefo’ [J]. Hort Science,2001,36 (6): 1049-1052.
[24] HUH Y J,LIM J H,PARK S K,et al. Endogenous indole-3-acetic acid and transzeatin ribosides in relation to axillary bud formation in standard chrysanthemum [J]. Horticulture Environment and Biotechnology,2011,52(2): 128-132.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
现代园艺(2021年23期)2021-12-01
湖北农业科学(2021年13期)2021-08-04
World Journal of Clinical Cases(2020年9期)2020-05-13
西藏农业科技(2019年3期)2019-11-04
种子(2019年4期)2019-05-28
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
西部林业科学(2016年6期)2017-01-04
浙江农业科学(2016年11期)2016-05-04
