
甜菜抗逆基因P5CS的克隆及盐胁迫下的表达分析
2019-07-11 09:45张皓崔杰李俊良李昕晏王琮玉
中国糖料 2019年3期
张皓,崔杰,李俊良,李昕晏,王琮玉
(1.内蒙古农业大学农学院,呼和浩特010019;2.哈尔滨工业大学化学与化工学院,哈尔滨150000)
0 引言
甜菜(BetavulgarisL.)为藜科甜菜属植物,是我国重要糖料作物,主要在东北、西北及华北地区大面积种植[1],该作物具有抗逆性强、适应性广的特点。了解甜菜的抗逆机理,对于解决西北及华北甜菜主产区干旱和半干旱面积大、盐碱发生率高制约甜菜生产及提高农民收入水平都具有重要的作用。目前,甜菜耐盐性主要进行利用蛋白质组学和基因组学的技术筛选耐盐基因研究,而对其耐盐机制的研究还较少[2]。对二倍体栽培甜菜品系M14采用不同浓度NaCl胁迫处理,鉴定出其根、叶中的差异蛋白各为43个和75个,这些蛋白质主要参与光合作用、能量代谢、蛋白质折叠和降解以及逆境胁迫与防御等生理生化过程[3]。我们课题组前期也开展了甜菜耐盐性相关研究,如:甜菜耐盐品系筛选研究[4]、DDRT-PCR技术分析盐胁迫下甜菜耐盐相关基因差异表达研究[5]、利用双向电泳技术初步筛选甜菜盐胁迫相关蛋白研究[6]以及甜菜耐盐相关microRNA筛选及其靶基因表达研究[7]等。不仅获得了耐盐品系,还找到了耐盐相关基因片段及参与调控的代谢过程。本研究在前期工作基础上,以筛选的甜菜耐盐品系“O”68为材料,克隆了调节脯氨酸代谢的关键酶基因P5CS,并进行生物信息学分析,进一步了解掌握盐胁迫下P5CS基因表达情况,对了解逆境下脯氨酸积累的分子机理,提高作物的抗逆性意义重大。
1 材料与方法
1.1 实验材料
1.1.1 植物材料
甜菜“O”68品系是经过筛选的耐盐材料[4],甜菜双6品系是盐敏感材料,均为哈尔滨工业大学化工与化学学院甜菜课题组选育。
1.1.2 试剂盒及载体
RNA提取采用TAKARA的Mini BESTPlant RNA Extraction Kit试剂盒按说明书进行。反转录采用Takara公司的PrimeScriptTM RT reagent Kit(Perfect Real Time),按说明书操作。T-载体和胶回收试剂盒均为TAKARA公司产品。转化选用商品化大肠杆菌DH5α感受态细胞。
1.2 实验方法
1.2.1 甜菜幼苗的培育和处理
温室培育甜菜幼苗,控制室内温度、光照、通风,定期更换营养液。待甜菜长出2对、3对真叶时选择长势一致的甜菜幼苗进行盐胁迫处理。盐胁迫过程:300 mmol/L NaCl溶液分别处理0 h、12 h、24 h、48 h和72 h。
1.2.2 抗逆相关P5CS基因的克隆
提取甜菜幼苗叶片总RNA,反转录成cDNA后,进行PCR扩增,“产物”回收后与T-载体连接,经转化大肠杆菌、培养、鉴定阳性克隆,用于测序。
1.2.3 抗逆相关P5CS基因生物信息学分析
在NCBI(http://www.ncbi.nlm.nih.gov/)上对克隆的基因序列进行比对,利用TMHMM Server v.2.0在线预测氨基酸序列的跨膜区域。
1.2.4 盐胁迫下甜菜幼苗中P5CS基因的表达分析
采用实时荧光定量PCR技术对两个品系2对、3对真叶期不同时间盐处理的甜菜幼苗叶片进行P5CS基因的表达情况分析。
2 结果与分析
2.1 抗逆相关P5CS基因的克隆
2.1.1 总RNA的提取及质量检测
甜菜幼苗叶片总RNA的提取按着试剂盒方法进行,用1%琼脂糖凝胶电泳检测,结果如图1。
从图上可以看到,提取的总RNA条带清晰,几乎无降解,紫外可见分光光度计检测总RNA提取纯度,提取的RNA OD260/OD280在1.8~2.1之间,纯度较高,可用于下一步的实验。

图1 总RNA提取结果Fig.1 Total RNA extraction results

图2 P5CS基因克隆电泳图谱Fig.2 P5CS gene cloning and electrophoresismap
2.1.2P5CS基因的克隆与测序
将上述提取的总RNA反转录成cDNA,使用高保真Taq酶克隆P5CS基因,电泳检测结果如图2。在1、3、5和7号泳道上出现了大小约2 300 bp的条带,与预期相符,初步断定样品克隆成功。
PCR产物经加A尾反应及胶回收后与T-载体进行16℃过夜连接,经转化大肠杆菌、抗性平板培养筛选,对单菌落进行PCR扩增进行重组子的鉴定,筛选结果获得如图2同样大小的目标条带,可用于后续测序。
通过测序与序列分析结果表明,甜菜P5CS基因其全长为2 325 bp,编码框为2 151 bp,共编码716个氨基酸(见图3)。
2.2 P5CS基因的生物信息学分析
2.2.1 序列比对
经BLASTN进行序列比对发现:克隆的P5CS核苷酸序列与多个物种中的P5CS基因序列有高度的同源性,与蒺藜(KC292266.1)、烟草(HM854026.1)、水稻(AY574031.1)、枸杞(KF771023.1)、石蒜(KT247899.1)、碱茅(HQ637435.1)、大豆(FM999730.1)、海滨锦葵(KR029088.1)、芸豆(EU340347.1)的同源性分别为74.80%、74.31%、70.51%、73.75%、73.97%、69.26%、73.80%、72.25%,说明所克隆的片段为P5CS基因。分析还发现甜菜与C4植物玉米(DQ864376.1)、甘蔗(EF620362.1)以及桑科植物桑树(KC202259.1)的同源性不是很高,分别为56.02%、32.00%、52.55%,构建亲缘关系树见图4。

图3 P5CS基因序列Fig.3 P5CS gene sequence

图4 甜菜P5CS基因与其他物种的核苷酸序列亲缘关系树Fig.4 Genetic relationship tree between sugar beet P5CS gene and other species
将克隆的P5CS基因氨基酸序列与上述物种BLASTP比对,结果发现:与蒺藜、烟草、水稻、枸杞、石蒜、碱茅、大豆、海滨锦葵、芸豆的同源性分别为78.07%、77.25%、75.24%、77.19%、78.35%、73.02%、76.36%、78.21%、78.19%,甜菜与玉米、甘蔗、桑树的同源性也不很高,分别为59.08%、35.06%、55.08%,见图5。构建亲缘关系树见图6。

图5 甜菜P5CS基因氨基酸序列与其他物种多序列比对结果Fig.5 Comparison of amino acid sequence of sugar beet P5CS gene with multiple sequences of other species
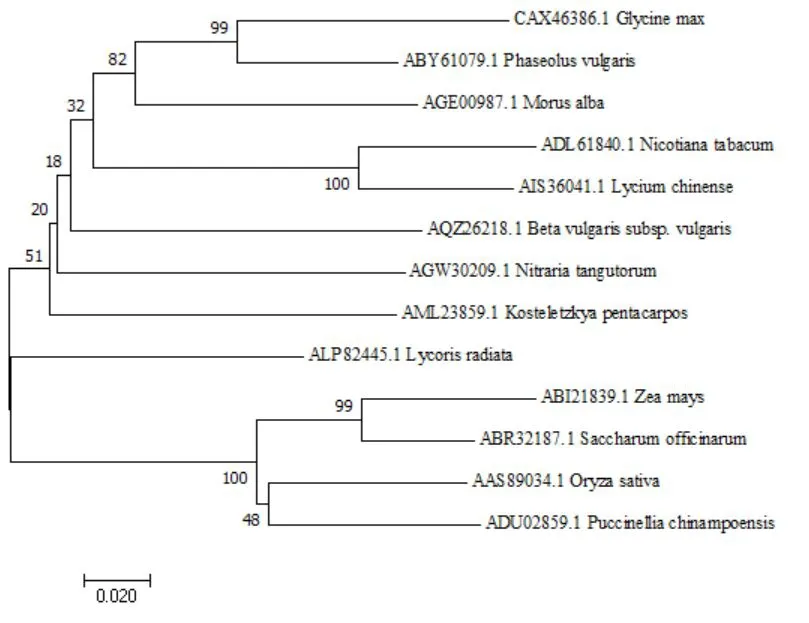
图6 甜菜P5CS基因与其他物种的核苷酸序列亲缘关系树Fig.6 Sugar beet P5CS gene is related to the nucleotides sequence of other species
核苷酸及氨基酸序列比对结果均证实,已克隆的片段确定为甜菜的P5CS基因。序列提交到GENBANK,已通过审核,基因ID:KY019201。
2.2.2 氨基酸理化性质分析
氨基酸理化性质分析表明,分子量为77.4 kDa,等电点(pI)为5.72,负电荷残基数谷氨酸(Glu)+天冬氨酸(Asp)为96,正电荷残基数赖氨酸(Lys)+精氨酸(Arg)为83,不稳定系数为37.89(<40),说明该蛋白比较稳定;疏水系数为-0.095,说明该蛋白是亲水蛋白。该蛋白序列中亮氨酸(Leu)含量最多,其次为丙氨酸(Ala)、缬氨酸(Val)、甘氨酸(Gly)、天冬氨酸(Asp)、硒半胱氨酸(Sec)。P5CS蛋白的氨基酸组成及含量见图7。
经TMHMMServer v.2.0软件预测氨基酸序列的跨膜区域可知:该序列无跨膜区域,蛋白均在膜外,属于膜外蛋白。
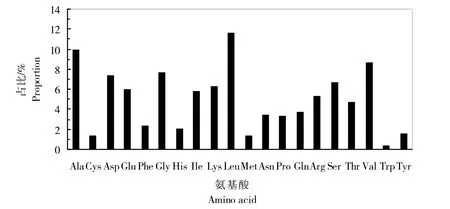
图7 P5CS蛋白的氨基酸组成及含量Fig.7 Amino acid composition and content of P5CSprotein
2.2.3 结构域分析
用软件Inter Pro对氨基酸序列进行结构域预测,共预测到8个结构域,包含有Δ1-吡咯啉-5-羧酸合酶(P5CS)结构域。此外还发现,该蛋白参与的生物过程包括脯氨酸生物合成过程(GO:0006561),代谢过程(GO:0008152)以及氧化还原过程(GO:0055114)。其作用是催化活性(GO:0003824)、谷氨酸-5-激酶活性(GO:0004349 activity)、谷氨酸-5-半醛脱氢酶活性(GO:0004350)以及氧化还原酶活性。
2.2.4 盐胁迫下甜菜幼苗P5CS基因表达情况分析
本研究对2对、3对真叶期甜菜幼苗进行了不同时间高盐胁迫处理P5CS基因表达量变化分析,结果如图8。从图中可以看出不同品系、不同生长时期P5CS基因的表达量随着胁迫处理时间的延长均呈现上调趋势。说明随着受盐胁迫的程度逐渐加深,P5CS基因的表达量增加,进而增加酶活性,促进脯氨酸合成代谢,以此增强幼苗的抗胁迫能力。图中还可以看出,耐盐品系“O”68和盐敏感品系双6的P5CS基因的表达存在差异,耐盐品系“O”68上调更加明显。
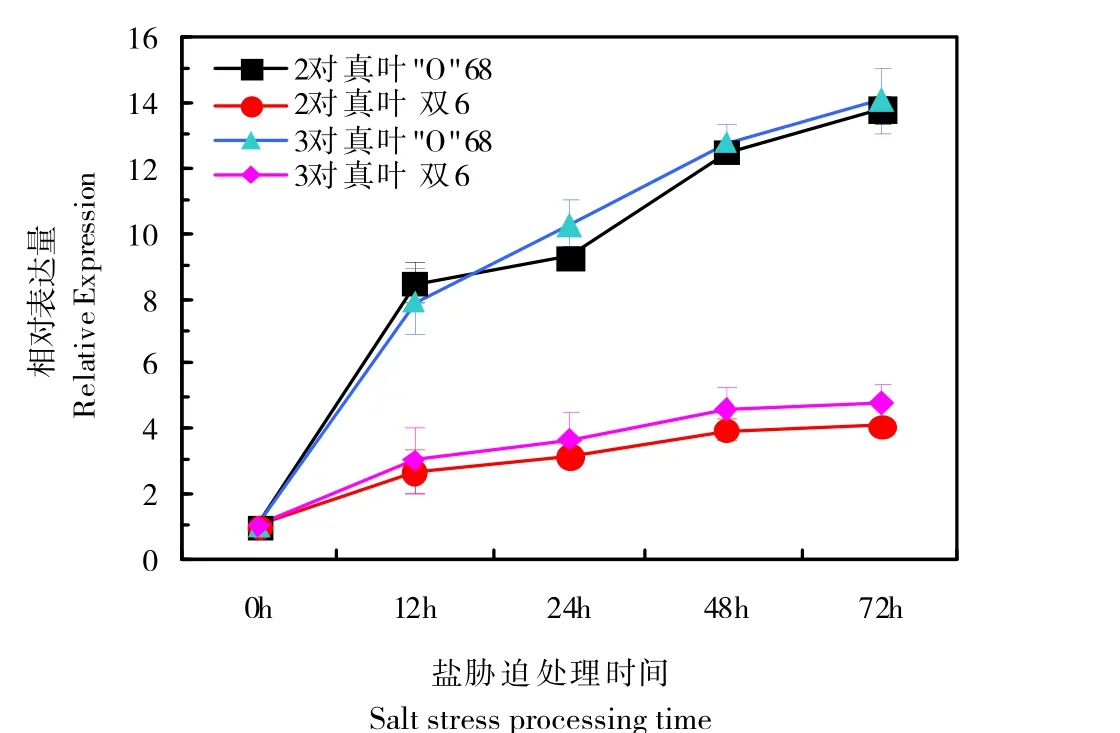
图8 盐胁迫下甜菜幼苗叶片中P5CS基因相对表达量变化Fig.8 Changes of relative expression of P5CS gene in leaves of sugar beet seedlings under salt stress
3 结论
本研究成功克隆了P5CS基因(ID:KY019201),生物信息学分析ORF为2 151 bp,编码由716个氨基酸组成的蛋白,该蛋白具有P5CS酶结构域,是脯氨酸合成代谢关键酶。qRT-PCR分析表明:盐胁迫下甜菜幼苗P5CS基因表达量随着胁迫处理时间的延长呈上调趋势,且幼苗受胁迫程度加深,而P5CS基因的表达上调明显。
4 讨论
与脯氨酸代谢相关的酶有多种,△1-吡咯啉-5-羧酸合成酶(P5CS)是由P5CS基因编码的双功能酶,是催化脯氨酸(Pro)生物合成的限速酶[7]。P5CS基因已从拟南芥、烟草、水稻、冰草等多种植物中分离出来,对其功能的多方面研究表明,P5CS基因在多种植物呈过表达或超量表达,转基因植株对干旱、高盐及重金属等非生物胁迫的抗逆性及耐受性明显增强,由此证明,P5CS基因可显著提高植物细胞的渗透调节能力。此外,在拟南芥中,已经分离了两个具有不同功能的P5CS基因,即P5CS1和P5CS2,其中P5CS1基因似乎在所有器官中普遍存在并表达,并且可通过应激迅速诱导,而P5CS2只在经历分裂的细胞中表达[8]。作物中水稻P5CS1基因同样也受盐碱、干旱、低温等非生物胁迫诱导,在各器官组织中普遍表达;P5CS2基因仅在成熟植物中表达,并可被氯化钠和甘露醇诱导。
本研究克隆的甜菜P5CS基因与不同物种的P5CS基因在序列上存在一些差别,该基因的克隆对后续转P5CS基因构建突变体以及根据P5CS基因表达评估甜菜受胁迫程度都具有重要意义。
猜你喜欢
中老年保健(2022年3期)2022-11-21
沈阳农业大学学报(2022年3期)2022-08-16
中国糖料(2022年2期)2022-04-06
黑龙江粮食(2021年9期)2021-12-12
今日农业(2021年21期)2021-11-26
安徽化工(2021年3期)2021-05-29
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
