
金黄色葡萄球菌SaeP蛋白表达条件的优化
2020-03-23 11:42刘佳璐郑维范若辰权春善
中国乳品工业 2020年1期
刘佳璐,郑维,范若辰,权春善
(1.大连民族大学a.生命科学学院;b.生物技术与资源利用教育部重点实验室,辽宁大连116600;2.大连理工大学生命科学与技术学院,辽宁 大连116024)
0引言
金黄色葡萄球菌(Staphylococcusaureus),是导致奶牛乳房炎的主要病原菌之一[1-4],其中的SaeRS双组分信号转导系统调控多种毒力因子的表达,由传感器组氨酸激酶SaeS和响应调节剂SaeR以及脂蛋白SaeP、膜蛋白SaeQ等两种辅助蛋白组成。SaeP与SaeQ以及SaeS共同形成三元复合物[5-6],该三元复合物可以激发SaeS磷酸酶活性,对SaeR蛋白进行磷酸化,使下游的Sae操纵子表达,最终激发多种毒力因子生成。SaeP蛋白表达量较低,限制了Sae系统的深入研究。本课题以优化SaeP蛋白表达条件为目的,通过改变重组载体构建方式和诱导温度等方式,确定最优SaeP表达条件,提高SaeP蛋白的表达量,推动SaeRS系统的整体研究,为研究金黄色葡萄球菌致病机理提供理论依据。
1实验
1.1材料与试剂
1.1.1菌种和质粒
金黄色葡萄球菌、p ET-28a、Tuner(DE3)、C43(DE3)、BL21(DE3)-pLysS以及BL21(DE3)-RIL均由本实验室保存。含有saep基因的质粒pET-28a-saep由本实验室构建。Trans5αChemically Competent Cell(目录号:CD201)和BL21(DE3)Chemically Competent Cell(BL21(DE3)目录号:CD601)。
1.1.2主要试剂
Nco I,Xho I,T4 DNA Ligase,Dpn I,DL2,000 DNA Marker,Blue Plus II Protein Marker(14-120 ku),WB封闭液(in PBS)、SanPrep柱式DNA胶回收试剂盒,基因组提取试剂盒,质粒提取试剂盒,Axyprep PCR清洁试剂盒,配制培养基用酵母提取物、胰蛋白胨,其余试剂均为国产分析纯。
1.1.3溶液与培养基的配制
1 mmol/L IPTG的配制:将2.38 g IPTG溶解于水中,定容至100 mL。
LB培养基配方(1 L):胰蛋白胨10 g、酵母提取物5 g、氯化钠10 g,定容至1 L。配制固体培养基则再加入20 g的琼脂粉。配制完成后将p H值调节至7.0,使用时根据需要添加抗生素。
SOB培养基配方(1 L):蛋白胨20 g、酵母提取物5 g、氯化钠0.5 g加入950 mL水溶解后,加入浓度为0.25 mol/L的KCl溶液10 mL,用NaOH调节p H值到7.0,定容至1 L,高压灭菌20 min。(该溶液使用前加入5 mL灭菌的浓度为0.002 mol/L的MgCl2溶液)。
SOC培养基:将SOB培养基高压灭菌后,降低温度到60℃以下时,加入20 mL用0.22μm滤膜过滤除菌的浓度为1 mol/L葡萄糖溶液,即为SOC培养基。
1.1.4 4种感受态细胞的制备
采用氯化钙法[7]将Tuner(DE3)、C43(DE3)、BL21(DE3)-pLysS以及BL21(DE3)-RIL制作成相应的感受态细胞。
In the first experiment,we demonstrate the performance of the proposed algorithm under different angular distributed functions.The parameters of two ID sources areandThe numberof snapshots is set as T=500 and the Signal-to-Noise Ratio(SNR)is equal to 5 dB.We perform the algorithm in three cases:
1.2方法
1.2.1重组质粒pET-28a-saep的构建
通过酶切酶连的方式重新构建重组质粒p ET-28a-sae p。选取酶切位点为Nco I和Xho I,引物设计序列见表1。重组质粒构建完成后,通过双酶切的方法对其进行验证。
通过无限克隆法(Restriction Free Cloning,RF法)构建重组质粒p ET-28a-saep。用RF-Cloning设计引物。引物设计序列见表1。通过PCR仪对目的基因进行扩增,并且将目的基因与质粒pET-28a相连,完成重组质粒pET-28a-saep的构建。通过对重组质粒pET-28a-saep的PCR产物进行电泳验证。
引物的合成以及检测结果正确的质粒的测序工作均由上海生工生物工程技术服务有限公司完成。SDS-PAGE分析以及Western blot检测。通过Western Blot实验对蛋白进行验证,再通过SDS-PAGE蛋白电泳将两种方法构建的重组蛋白进行对比分析。
1.2.3宿主菌的优化
将重组质粒p ET-28a-saep分别转入大肠杆菌BL21(DE3)、Tuner(DE3)、C43(DE3)、BL21(DE3)-p LysS以及BL21(DE3)-RIL中,均匀涂布于带有卡那霉素抗性的LB固体培养基上,37℃过夜培养,挑取单菌落,培养10 h,再进行扩大培养,OD 600达到0.6时加入终浓度为0.5 mmol/L的IPTG。30℃诱导8 h后,分别取5种不同宿主的菌液进行超声破碎后,离心30 min,取上清作为样品加入进行SDS-PAGE分析。
1.2.4诱导剂浓度的优化
根据本文2.2.2的实验结果,选择最适宜的宿主菌BL21(DE3)进行诱导剂浓度的优化。将已经转入重组质粒pET-28a-saep的BL(DE3)接种LB液体培养基中,培养10 h,再进行扩大培养,至OD 600达到0.6时加入IPTG,至终浓度分别为0,0.5,0.9,1.3和1.5 mmol/L,30℃诱导8 h,分别取5种不同诱导剂浓度的菌液进行超声破碎后,离心30 min取上清作为样品进行SDS-PAGE分析。
1.2.5诱导温度及诱导时间的优化
根据上述2.2.3中的实验结果,将诱导剂终浓度定为0.5 mmol/L,进行下一步诱导温度及诱导时间的优化。将温度按梯度分为16,20,24,28以及30℃(原诱导温度),诱导时间分为8,12,24 h,分别进行平行实验,具体诱导温度与诱导时间的设置如表2所示。
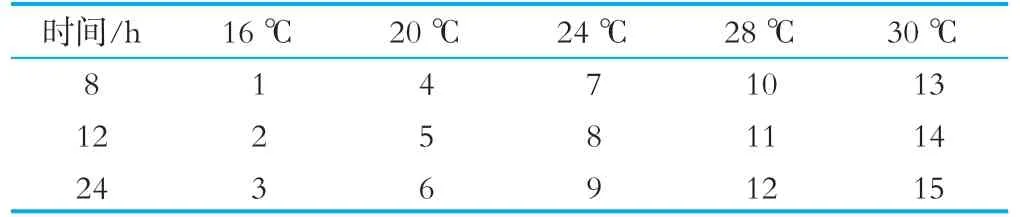
表2诱导时间及温度设置
按实验计划进行活化、转接、诱导、收菌和破碎,离心后取上清作为样品进行SDS-PAGE分析。
1.3数据统计
所有实验重复3次,实验结果平均值±标准偏差表示。采用Quantity One软件进行蛋白定量分析,采用2010软件进行数据分析,采用SPSS 17.0软件进行差异显著性分析(P≤0.05)。

表1引物序列
1.2.2构建方式的优化
将两种方式构建的重组质粒分别转入BL21(DE3)宿主中,涂布于含卡那霉素抗性的LB固体培养基上,37℃过夜培养,挑取单菌落,培养10 h,将菌液以1%的接种量接种于50 mL的LB液体培养基中,37℃,180 r/min进行培养,OD 600达到0.6时加入终浓度为0.5 mmol/L的IPTG。30℃诱导8 h后,将菌液进行超声破碎后离心30 min,取上清作为样品,进行
2结果与分析
2.1重组质粒的构建和验证
2.1.1酶切酶连法构建重组质粒pET-28a-saep验证结果
图1为酶切酶连法构建重组质粒p ET-28a-saep双酶切鉴定电泳结果图。由图1可以看出,重组质粒条带大小与预期相符,且双酶切后出现与目的基因条带大小一致的电泳条带,质粒测序结果正确,认定酶切酶连法成功构建重组质粒。
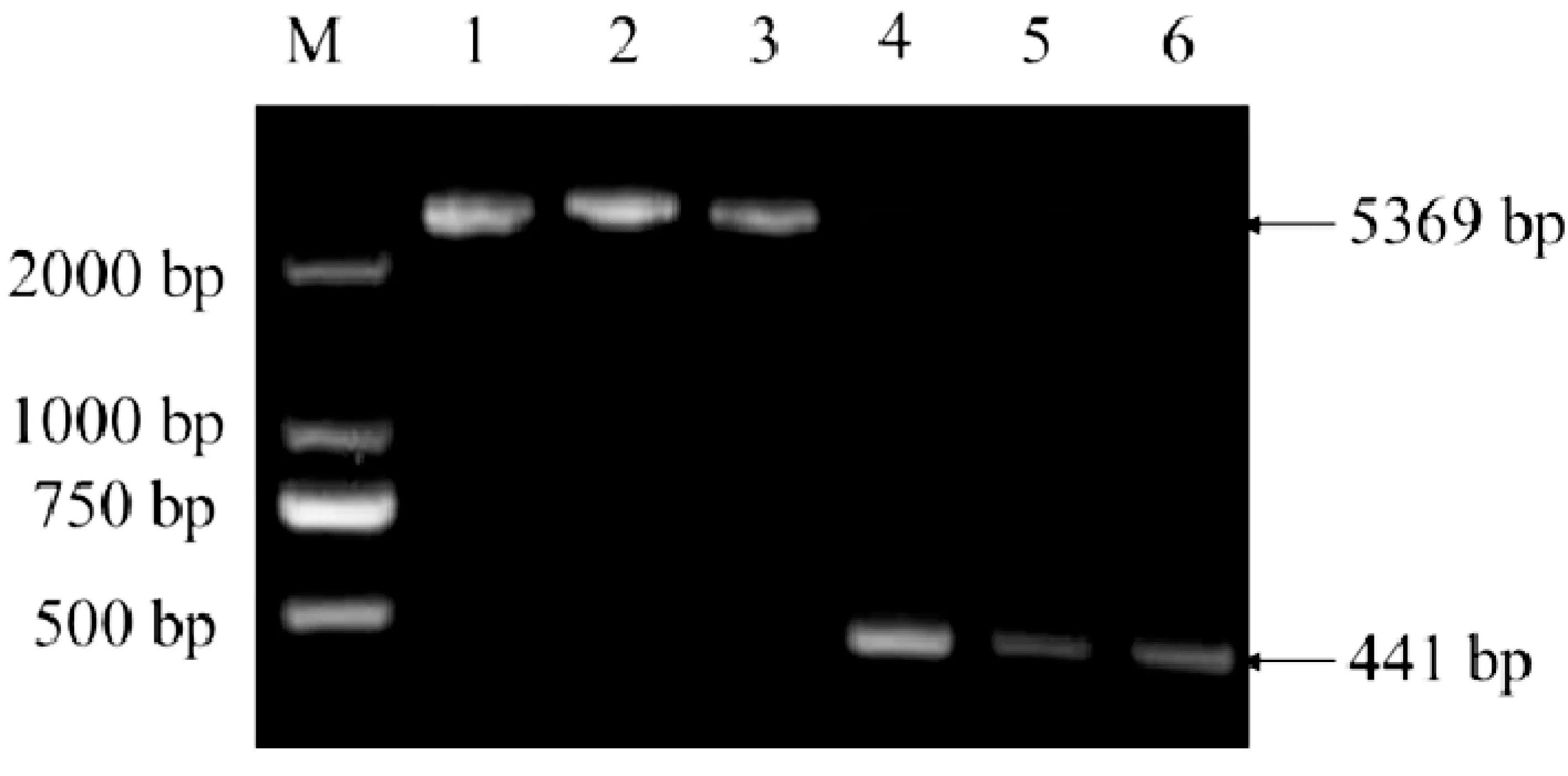
图1酶切酶连法构建重组质粒pET-28a-saep双酶切鉴定电泳结果
2.1.2 RF法构建重组质粒p ET-28a-saep验证结果
由图2可以看出,以重组质粒pET-28a-saep为模板,进行PCR扩增,能够扩增出单一且分子量大小符合预期的目的条带。质粒测序结果正确,认定通过RF法成功构建重组质粒。图2中,1~3为saep基因的PCR扩增产物。
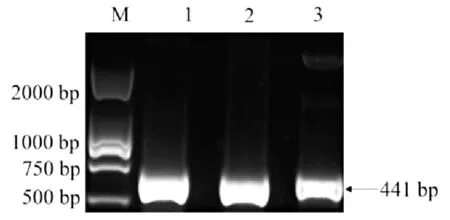
图2 PCR产物的验证结果
2.2重组蛋白表达验证结果
重组质粒pET-28a-saep转入大肠杆菌BL21(DE3)的诱导产物经SDS-PAGE分析,未添加IPTG诱导的产物与添加IPTG诱导后的产物在大约16 ku位置存在显著差异。Western blot结果如图3所示,表达产物能够与带有组氨酸标签的抗体结合,从而显现出一条清晰的条带,大小符合预期。
2.3构建方式的优化结果
由图4(a)可知,RF法与酶切酶连法构建的重组蛋白相对表达量有明显差异。通过Quantity One和SPSS软件对其进行分析可知,全菌体蛋白状态对比下,统计学分析显示RF法构建全菌体蛋白的相对表达量明显高于酶切酶连法构建全菌体蛋白的相对表达量,更利于后续纯化工作的进行。镍离子亲和层析纯化后的酶切酶连法构建重组表达蛋白进一步损失,而RF法构建重组蛋白SaeP的相对表达量大幅提高,所以选择RF法构建的重组质粒p ET-28a-saep继续后续实验的研究。
2.4宿主菌的优化结果
通过Quantity One和SPSS软件对重组蛋白SaeP的相对表达量进行分析。由图4(b)可知,通过RF法构建重组质粒p ET-28a-saep以大肠杆菌BL21(DE3)为宿主菌时相对表达量高于其他宿主菌,占菌体蛋白的19.9%,统计学分析显示以大肠杆菌BL21(DE3)为宿主菌与其他宿主菌相比,具有显著差异,所以选择以大肠杆菌BL21(DE3)为宿主菌进行后续实验。重组蛋白的表达量受基因自身性质、表达载体宿、主菌以及培养条件等多种因素的影响,通过对比不同宿主菌对于重组SaeP蛋白的表达量,以确定较为适合的宿主菌,提高了表达效率[8-11]。

图3重组表达产物SDS-PAGE蛋白电泳图及WesternBlot结果
2.5诱导剂浓度的优化结果
通过Quantity One和SPSS软件对重组蛋白SaeP的相对表达量进行分析,如图4(c)所示,重组蛋白的相对表达量在IPTG终浓度为0~0.5 mmol/L范围内,成上升趋势,当IPTG终浓度达到0.5 mmol/L时重组SaeP蛋白的相对表达量达到峰值,占菌体蛋白的20.16%。诱导剂IPTG的浓度与蛋白表达速率相关,在一定范围内提高IPTG的浓度会提高蛋白的表达量,但浓度进一步提高后,相对表达量反而有所减少,说明IPTG具有一定的细胞毒性,浓度过高不利于菌株的生长并且抑制细胞活性,所以适当降低IPTG的浓度可能更利于重组蛋白的表达[11-13]。统计学分析显示浓度为0.5 mmol/L的IPTG诱导重组蛋白相对表达量与其他浓度相比,具有显著差异,所以在本实验中,选择浓度为0.5 mmol/L作为诱导重组蛋白SaeP的诱导剂浓度。
2.6诱导温度及诱导时间的优化结果
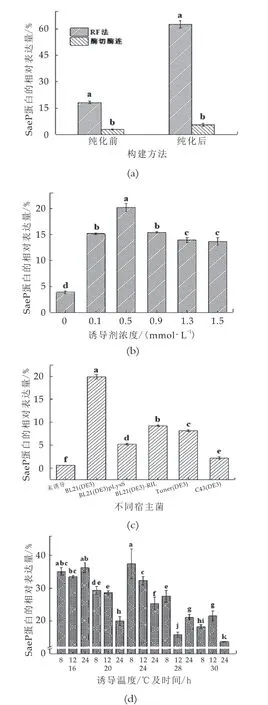
图4诱导条件优化结果
通过Quantity One和SPSS软件对重组蛋白SaeP的相对表达量进行分析。由图4(d)可知,在诱导条件为16℃诱导8 h,16℃诱导12 h,16℃诱导24 h及24℃诱导8 h相对表达量较高,通过对比实验结果发现低温诱导条件下重组蛋白表达效果更好。低温条件下菌体生长速率下降,很大程度上避免了菌体失活与质粒稳定性下降等问题,低温诱导能够提高外源蛋白稳定性和可溶性,从而增加了重组蛋白的表达量[14]。重组SaeP蛋白在16℃条件下诱导8 h后,并没有随着诱导时间的延长而明显增加。推测原因是随着诱导时间的增加,菌体内蛋白酶对重组蛋白的降解量增加,并且诱导时间过长产生的氨基酸会抑制目的蛋白的合成,所以长时间的诱导不利于重组蛋白的表达[15-16]。所以选择16℃诱导8 h为重组SaeP蛋白的诱导温度及诱导时间。
3结论
本研究通过改变重组载体构建方式、宿主菌、诱导剂终浓度、诱导时间诱导温度等,对金黄色葡萄球菌SaeP蛋白的表达条件进行优化,以提高SaeP蛋白的表达量。前期实验结果表明,无限克隆方法获得的表达载体在各条件的试验中,表达量均高于酶切酶连方法所得到的表达载体,因此条件优化部分全部以无限克隆方法得到的表达载体为主要研究对象。成功优化金黄色葡萄球菌SaeP蛋白的诱导表达条件,最佳表达条件为:通过无限克隆法建重组质粒,以BL21(DE3)为转化宿主,IPTG浓度为0.5 mmol/L,诱导温度为16℃,诱导时间为8 h,金黄色葡萄球菌SaeP蛋白的最高表达量占总蛋白的35.14%,较原诱导条件提高25.49%,为促进金黄色葡萄球菌的Sae系统对于毒力因子生成研究的提供支持。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
江苏农业科学(2022年2期)2022-02-15
江西农业学报(2021年4期)2021-04-20
江西农业学报(2021年1期)2021-02-01
科学(2020年3期)2020-11-26
北华大学学报(自然科学版)(2020年5期)2020-10-21
三农资讯半月报(2020年11期)2020-06-21
当代水产(2020年3期)2020-06-15
上海医药(2016年23期)2016-12-22
