
新藤黄酸抑制人肝癌细胞BEL-7402自噬作用研究
2020-06-08 06:34王玉涵苏婧婧李庆林
安徽中医药大学学报 2020年3期
展 凡,王玉涵, 王 萌,郑 凤,苏婧婧,程 卉,李庆林
(安徽中医药大学科研实验中心 新安医学教育部重点实验室,安徽 合肥 230038)
肝细胞癌(hepatocellular carcinoma,HCC)是中国常见的恶性肿瘤之一,其死亡率居肿瘤中第3位[1]。手术切除、肝移植是HCC主要的治疗方法[2],但预后不良导致的HCC患者5年生存率低于5%[3]。越来越多的研究表明,自噬与HCC的危险因素相关,如氧化应激、代谢功能障碍、肝酒精紊乱和脂肪肝疾病[4-5]。细胞自噬是由基因调控并在进化上保守的细胞内降解通路。在自噬过程中,自噬体与溶酶体融合形成自噬溶酶体,其可降解细胞质中受损、变形的大分子物质和细胞器,并将降解产物循环利用,从而为细胞的正常生存、代谢提供原料[6]。自噬作为抗肿瘤药物治疗的潜在作用靶点近年来已得到确认。但中药活性成分新藤黄酸(gambogenic acid,GNA)对人肝癌细胞BEL-7402自噬的影响尚不清楚。本研究通过体外试验,初步探讨GNA抑制人肝癌细胞BEL-7402增殖的作用与自噬的关系。
1 仪器与试剂
1.1 主要仪器 CO2培养箱:日本SANYO公司;洁净工作台(SW-CJ1F):江苏省苏净集团;倒置显微镜(Olympus CKX41)和倒置荧光显微镜(Olympus BX51):日本Olympus公司;全自动多功能酶标仪(SpectraMax M2e):美国MD公司;灭菌锅:上海申安医疗器械有限公司。
1.2 主要试剂和药品 GNA:陕西省宝鸡市辰光生物科技有限公司(纯度98%,批号HN045248198);RPMI 1640培养基:美国Gibco公司;胎牛血清:杭州四季青生物工程材料有限公司;氯喹(Chloroquine ,CQ)(货号C6628):Sigma公司;雷帕霉素(Rapamycin ,Rap)(货号53123-88-9):Sigma公司;阿霉素(货号 25316-40-9):Selleck公司;anti-beclin 1(批号3495T)、anti-LC3B(批号3868T)、anti-p62 (批号8025T)和anti-actin (批号3700T):美国CST公司。
2 方法
2.1 细胞活力和药物敏感性检测 将对数生长期的细胞按照5×103/mL接种于96孔板。设立空白对照组(0 μmol/L GNA)和不同浓度给药组,每组设6个复孔。待细胞贴壁良好,弃培养基,每孔分别加入0、2、4、8、16 μmol/L的GNA溶液100 μL,放置37 ℃,5% CO2的培养箱24、48、72 h后弃去培养基,在终止培养前4 h每孔加入MTT培养液20 μL继续孵育。孵育终止弃去培养液,每孔加入二甲基亚砜150 μL,摇床上避光振荡10 min后用酶标仪在570 nm波长处测每孔的光密度(optical density,OD)值,按照上述过程,重复3次,计算细胞存活率。
2.2 单丹磺酰尸胺(monodansylcadaverine,MDC)染色荧光显微镜观察自噬体的形成 将对数生长期的细胞以每孔3×105个接种于6孔板,待细胞生长良好,按照实验要求分为空白对照组、Rap组(50 μmol/L)和GNA(1.5、3、6 μmol/L)给药组。细胞培养24 h,收集对照组和给药组细胞,PBS洗2遍,用1 mL PBS重悬,加入1 μL MDC储存液(浓度为50 mmol/L),使MDC的终浓度为0.05 mmol/L。37 ℃避光孵育30 min,PBS洗涤3遍。吸取微量细胞悬液滴于玻片,盖上盖玻片,用荧光显微镜观察自噬小体。
2.3 透射电子显微镜观察自噬体水平 按照试验要求设立空白对照组、Rap组(50 μmol/L)和GNA(1.5、3、6 μmol/L)给药组。细胞培养24 h,收集细胞,离心使细胞沉淀成团,将戊二醛沿着管壁缓慢加入用于固定细胞,放入4 ℃冰箱。次日用PBS洗涤细胞3遍,每次5 min;再用锇酸于4 ℃固定1.5 h,PBS洗3次,每次5 min;按以下步骤脱水:50%乙醇10 min,70%乙醇10 min,90%乙醇10 min,100%乙醇10 min,100%丙酮10 min 2次;环氧树脂包埋过夜;再放入45 ℃烘箱内固化12 h;切片后用醋酸铀和枸橼酸铅染色,在透射电子显微镜下观察细胞的超微结构。
2.4 Western blot法检测自噬相关蛋白微管相关蛋白1轻链3(microtubule associated protein 1 light chain 3,LC3)、p62、beclin 1的表达情况 将对数生长期的细胞按照每孔5×106个接种于6孔板,待细胞生长良好,按照试验要求分为空白对照组、Rap组(50 μmol/L)和GNA(1.5、3、6 μmol/L)给药组以及CQ组(10 μmol/L),细胞培养24 h。提取各组总蛋白,BCA试剂盒检测蛋白质浓度。蛋白上样量定为20 μg。进行SDS-PAGE电泳,结束后将蛋白转至NC膜,在5%脱脂奶粉封闭液中封闭2 h,弃封闭液并洗膜。加入对应的一抗,4 ℃孵育过夜,洗膜后加入二抗,摇床慢摇2 h,洗膜后曝光处理。并计算目标蛋白与内参蛋白(β-actin)条带灰度比值,以此来确定各组蛋白的相对表达量。以上实验重复3次。

3 结果
3.1 GNA对人肝癌细胞BEL-7402存活率的影响 不同浓度的GNA作用于BEL-7402细胞24、48、72 h后,BEL-7402细胞存活率随时间和剂量依赖性降低,IC50分别为6.408、4.077、3.001 μmol/L。见图1。
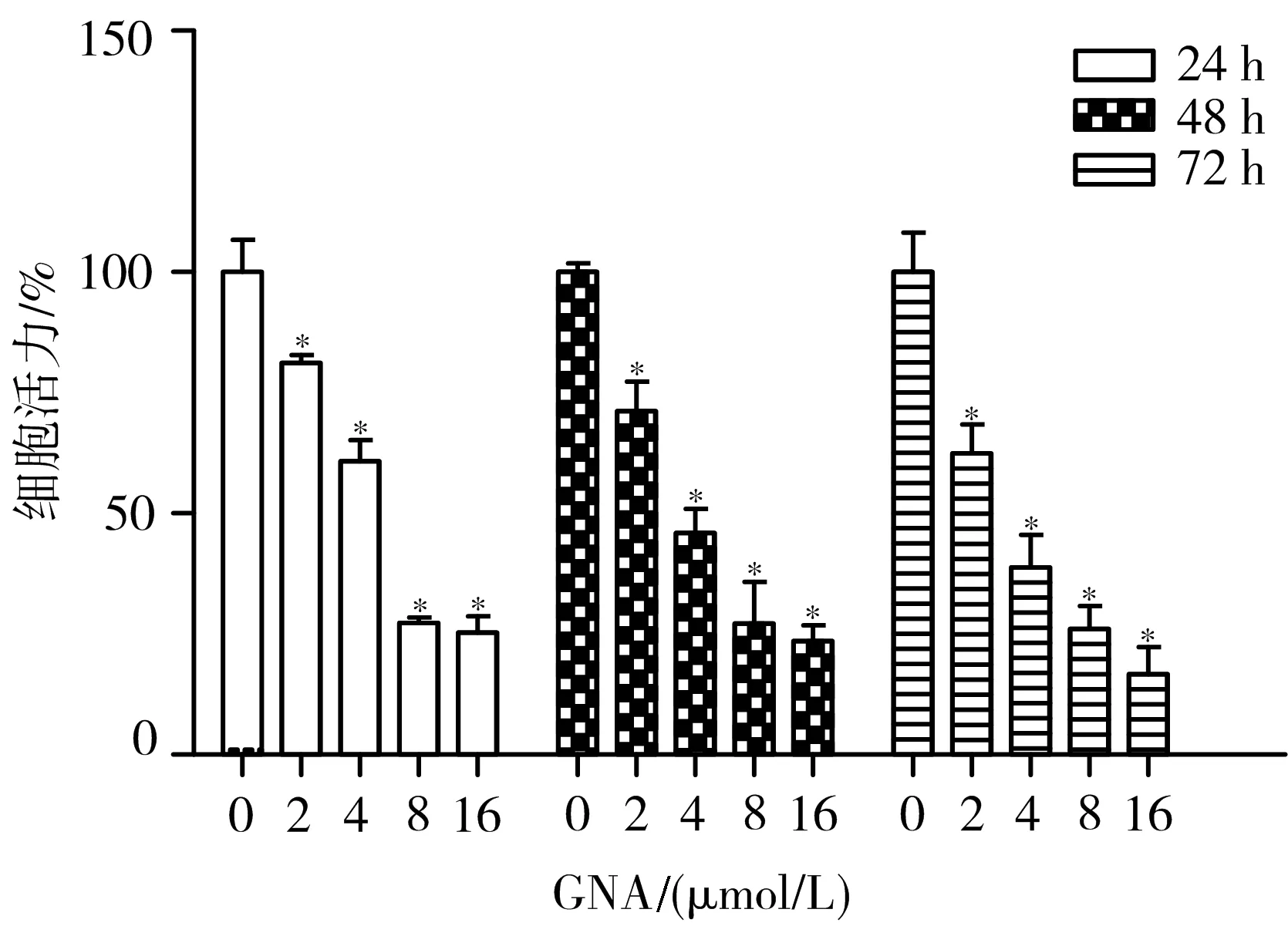
注:与空白对照组比较,*P<0.05
3.2 GNA对人肝癌细胞BEL-7402形态的影响 空白对照组细胞贴壁生长,细胞数目较多且排列紧密,胞质均匀透明;Rap组细胞贴壁生长较少,细胞脱落、悬浮增加,形状不一,细胞碎片化;不同浓度GNA组,随药物浓度增加,细胞贴壁数量减少,排列不紧密,细胞悬浮增加,高浓度GNA组出现细胞碎片化。见图2。
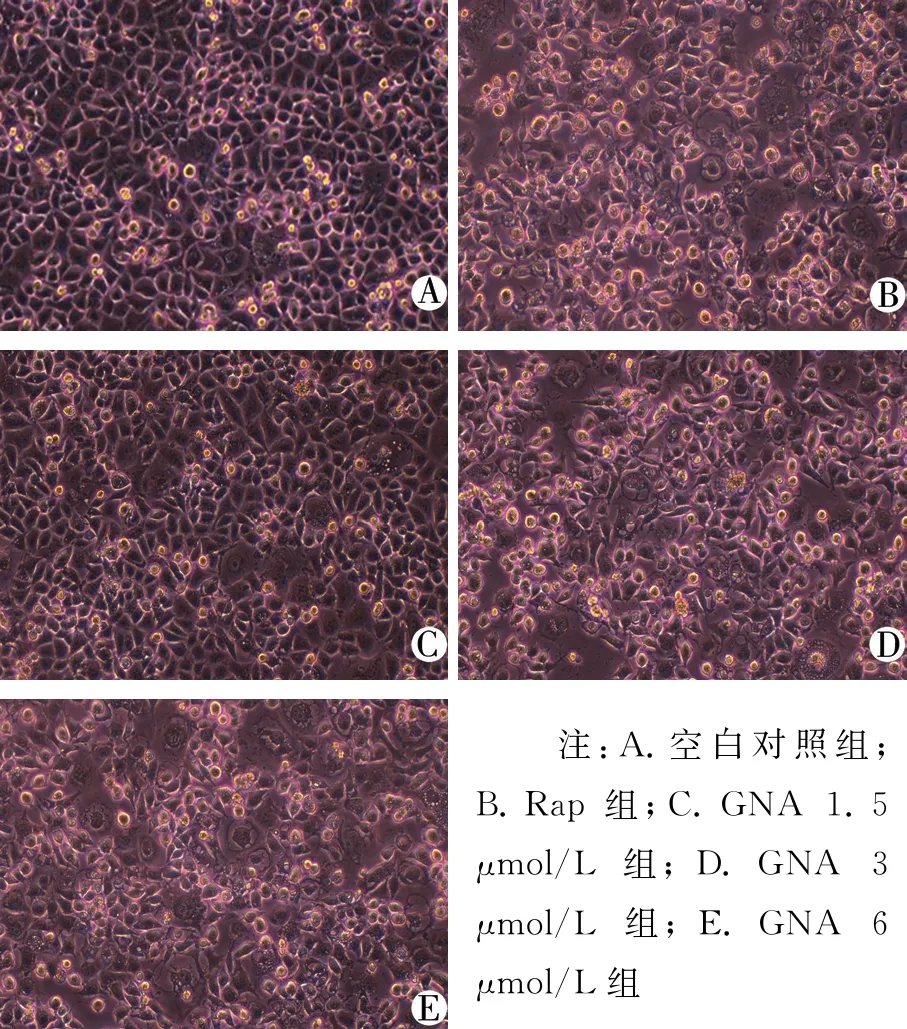
注:A.空白对照组;B.Rap组;C.GNA1.5μmol/L组;D.GNA3μmol/L组;E.GNA6μmol/L组
图2 倒置显微镜下观察GNA对人肝癌细胞BEL-7402形态的影响(10×40倍)
3.3 GNA对人肝癌细胞BEL-7402自噬小体形成的影响 荧光显微镜观察发现,空白对照组荧光强度较弱,自噬小体的数量较少;自噬诱导剂Rap组自噬小体数目较多;随GNA浓度增加,荧光强度增加即自噬小体增多。见图3。

注:A.空白对照组;B.Rap组;C.GNA1.5μmol/L组;D.GNA3μmol/L组;E.GNA6μmol/L组
图3 GNA对人肝癌细胞BEL-7402细胞自噬小体形成的影响(MDC染色,10×40倍)
3.4 GNA对人肝癌细胞BEL-7402自噬小体水平的影响 透射电子显微镜观察细胞自噬小体,空白对照组细胞形态正常,结构清晰,偶见少量自噬小泡;Rap组细胞浆中出现大量自噬空泡聚集;随GNA浓度的增加,自噬泡数目增多,堆积明显。图中箭头所指均为自噬小体。见图4。

注:A.空白对照组;B.Rap组;C.GNA1.5μmol/L组;D.GNA3μmol/L组;E.GNA6μmol/L组
图4 透射电子显微镜观察GNA对人肝癌细胞BEL-7402自噬小体水平的影响(1×20 000倍)
3.5 GNA对人肝癌细胞BEL-7402自噬相关蛋白表达的影响 不同浓度(1.5、3、6 μmol/L)GNA作用细胞24 h后,随药物浓度的增加,Beclin1、p62和LC3-Ⅱ蛋白表达水平均显著增加(P<0.05),LC3-Ⅰ蛋白水平无明显变化;Rap组各自噬相关蛋白的表达说明Rap可诱导人肝癌BEL-7402细胞发生自噬(见图5Ⅰ)。单独加入10 μmol/L CQ可降低Beclin1蛋白表达,增加LC3-Ⅱ蛋白表达以及p62蛋白的积累,结果提示GNA对人肝癌BEL-7402细胞自噬的影响可能与其抑制自噬后期进程有关(见图5Ⅱ)。
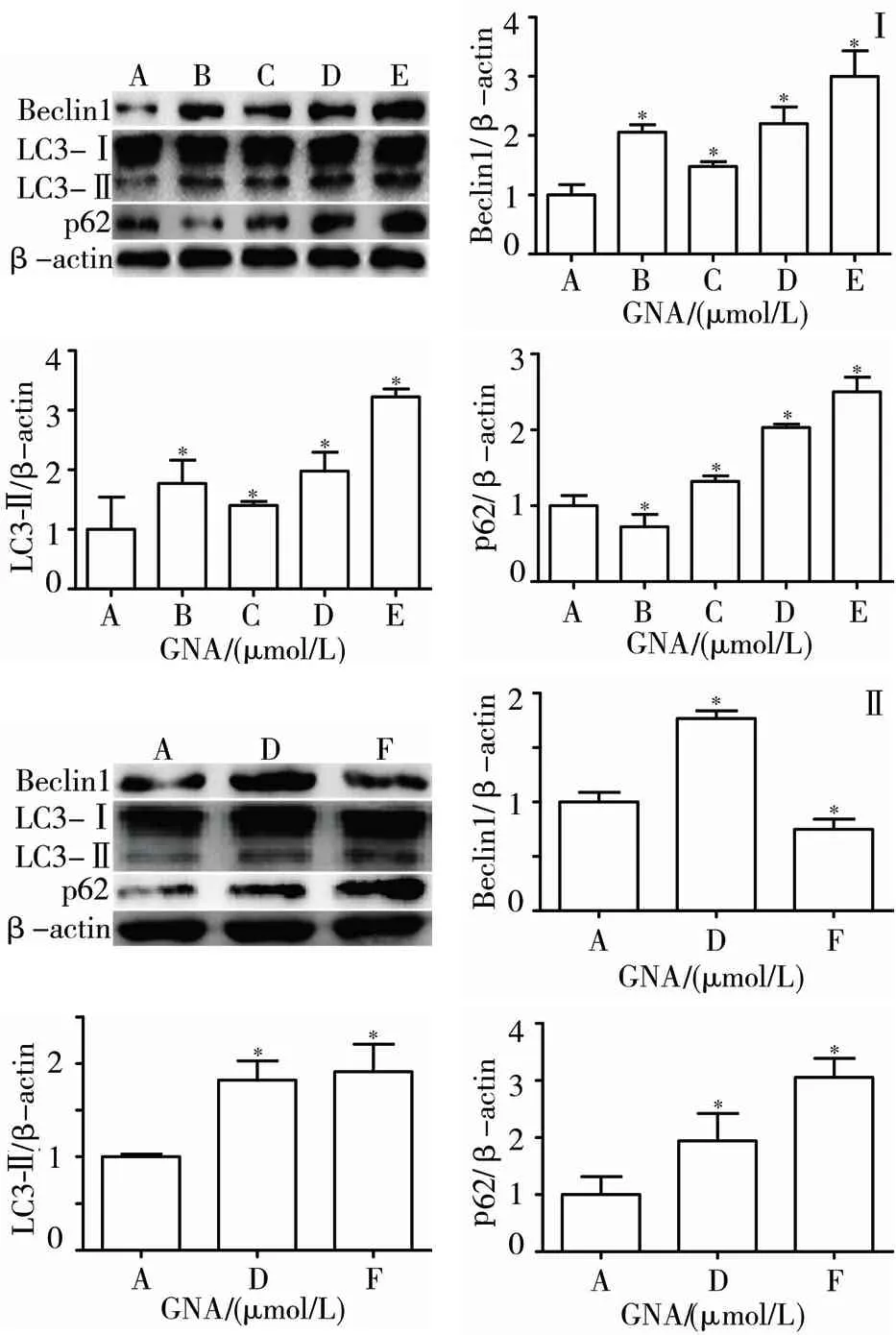
注:A.空白对照组;B.Rap组;C.GNA 1.5 μmol/L组;D.GNA 3 μmol/L组;E.GNA 6 μmol/L组;F.10 μmol/L CQ组;与空白对照组比较,*P<0.05

4 讨论
近年来,自噬被认为与癌症的发生和发展有关,当肿瘤细胞处于饥饿、化学治疗、低氧等应激情况时,肿瘤细胞内自噬水平升高,为肿瘤细胞生存提供必须的营养和能量,肿瘤细胞存活能力增强。越来越多的研究表明,中药有效成分可通过调控肿瘤细胞自噬水平来抵抗癌症,增加癌症治疗的可能性[7-8]。
中药藤黄系藤黄科植物分泌的干燥树脂[9],黄棕色,呈不规则块状。1984年吕归宝从藤黄中分离得到GNA,其具有良好的抗肿瘤作用。近年来,本课题组对GNA的系列研究发现,GNA能够抑制多种肿瘤细胞增殖,其抗肿瘤作用与调控线粒体氧化应激、调节细胞周期、诱导细胞凋亡以及抑制肿瘤细胞保护性自噬有关[10-14]。本研究表明,GNA在一定剂量范围呈时间和剂量依赖性抑制BEL-7402细胞增殖。GNA处理后,MDC染色荧光显微镜观察和透射电子显微镜观察结果均显示,细胞内自噬小体堆积。同时,Western blot检测结果表明,自噬上游蛋白Beclin1的表达增加,提示自噬被激活;LC3-Ⅰ向LC3-Ⅱ的转变增加,也提示自噬小体数量的增多。但是这些证据并不能特异性反映自噬水平的增加,如果自噬进程后期溶酶体功能受到抑制,也可能导致自噬体堆积,LC3-Ⅱ含量积累。值得一提的是,笔者发现LC3-Ⅰ向LC3-Ⅱ的转变的同时,LC3-Ⅰ的含量并没有显著变化,因此推测,GNA诱导的自噬可能并不是完整过程的自噬。此外,自噬溶酶体降解底物p62的表达增加,p62的总细胞蛋白表达水平与自噬水平呈反相关。因此笔者认为,GNA可能有阻断BEL-7402细胞自噬后期进程中溶酶体的功能。加入自噬溶酶体抑制剂CQ处理后结果显示,CQ对LC3和p62蛋白的调节作用与GNA一致,进一步证实了GNA可能部分通过抑制自噬促进肿瘤细胞死亡的猜测。但是GNA的这种抑制自噬促进细胞死亡的信号途径有待于进一步的研究。
猜你喜欢
中西医结合心脑血管病杂志(2022年19期)2022-11-19
新农业(2022年21期)2022-11-18
中国临床解剖学杂志(2022年1期)2022-11-15
检验医学与临床(2022年12期)2022-06-27
中国兽医学报(2022年4期)2022-06-17
河南科技(2022年6期)2022-04-22
食管疾病(2020年4期)2020-12-08
看世界·学术下半月(2020年7期)2020-09-10
现代职业教育·高职高专(2018年7期)2018-05-14
中国医药导报(2018年3期)2018-03-07
