
羊耳菊活性部位对PXR高表达HepG2细胞中CYP3A4蛋白表达的影响*
2020-11-03 07:25杨畅宋菲何俊奇王永林李勇军兰燕宇陈一飞刘亭
贵州医科大学学报 2020年10期
杨畅, 宋菲, 何俊奇, 王永林, 李勇军, 兰燕宇, 陈一飞, 刘亭**
(1.贵州医科大学 省部共建药用植物功效与利用国家重点实验室 & 贵州省药物制剂重点实验室, 贵州 贵阳 550004; 2.贵州医科大学 民族药与中药开发应用教育部工程研究中心, 贵州 贵阳 550004; 3.上海药品评审核查中心, 上海 201203)
细胞色素P450酶属于血红素-硫醇盐蛋白的超基因家族,可介导临床约90%的药物代谢[1-2],其活性及表达量的变化直接影响其底物在体内的生物转换[3-4],是许多药物发生疗效改变、毒性反应及药物间相互作用的原因之一[5-6]。细胞色素P450 3A4酶(cytochrome P450 3A4 enzyme,CYP3A4)作为细胞色素P450酶家族中重要成员,在临床前研究中检测药物对CYP3A4的诱导或抑制作用,已成为必不可少的一个环节[7]。羊耳菊[Inulacappa(Buch.-Ham. ex D. Don)DC.]又名山白芷、白牛胆和见消肿等,为菊科旋覆花属植物羊耳菊的全草或根,具有消肿止痛、祛风解热的功效,常用于感冒发热、风湿疼痛、痈疮疔毒、咽喉肿痛及乳痈等症[8]。本课题组前期对羊耳菊药材进行了较为系统的研究,筛选出了羊耳菊药材发挥疗效作用的活性部位[9-12]。目前以羊耳菊活性部位(main active constituents fromInulacappa, IC-MAC)为主要成份的药物有鼻康片、双羊喉痹通颗粒、菊黄上清含片、莲菊感冒胶囊等制剂,临床上应用广泛[13-15],课题组前期研究还发现IC-MAC可下调CYP3A4的比活性[16],但机制尚不清楚。本文通过构建可高表达CYP3A4转录调节因子——孕烷X受体(pregnane X receptor, PXR)的人肝癌HepG2细胞系,研究IC-MAC对PXR的基因转录调节活性的影响,探讨IC-MAC下调CYP3A4活性的机制。
1 材料与方法
1.1 材料
1.1.1细胞株和细菌 人肝癌HepG2细胞株(江西阿普斯戴尔生物科技有限公司),DH5α感受态大肠杆菌(北京博迈德基因技术有限公司),质粒pcDNA3.1-PXR由浙江大学药学院陈枢青教授惠赠。
1.1.2主要试剂 IC-MAC(自制[16]),G-418遗传霉素硫酸盐、氨苄西林及溶菌肉汤(luria-bertani,LB)液体培养基(北京索莱宝科技有限公司),质粒提取试剂盒(杭州爱思进生物技术有限公司),FuGENE®6 转染试剂(北京普洛麦格生物技术有限公司),4S Red Plus 核酸染色试剂盒(上海BBI生命科学有限公司),改良杜氏伊格尔培养基(dulbecco’s modified eagle medium,DMEM)培养基、胎牛血清、辣根过氧化氢酶标记的山羊抗兔及山羊抗小鼠(美国Thermo公司),兔抗PXR,重组PXR蛋白和小鼠抗重组人β-肌动蛋白(recombinant human beta-actin,β-actin;英国Abcam公司)。
1.1.3主要仪器 EL304型电子分析天平(上海梅特勒-托利多仪器有限公司),二氧化碳恒温培养箱(美国Thermo公司),TS100型倒置显微镜(日本Nikon公司),数显立式压力蒸汽灭菌器(上海博讯实业有限公司医疗仪器厂),PowerPacBasic型电泳仪、Trans-Blot Turbo型蛋白快速转印仪及GBOXChemiXL1.4型凝胶成像系统(美国Bio-rad公司),ZDP-150型恒温振荡培养箱(上海精宏实验设备有限公司),SNAP i.d.2.0 MultiBlot Frame快速孵育仪(美国Millipore公司)。
1.2 方法
1.2.1质粒pcDNA3.1-PXR扩增及提取 将质粒pcDNA3.1-PXR转化至DH5α感受态大肠杆菌中,并接种于含50 mg/L氨苄西林的LB液体培养基中培养13 h,根据Axygen质粒提取试剂盒的说明书对质粒进行提取,质粒图谱如图1所示。

图1 pcDNA3.1-PXR质粒图谱Fig.1 Map of pcDNA3.1-PXR plasmid
1.2.2细胞培养 人肝癌细胞HepG2用含10%胎牛血清的DMEM培养基,37 ℃的5%CO2培养箱中培养,当细胞汇合至80%左右时进行传代。
1.2.3高表达PXR-HepG2细胞模型的构建 HepG2细胞以每孔1.5×105个的细胞密度接种于24孔板培养24 h,待细胞长至70%,将FuGENE 6转染试剂与pcDNA3.1-PXR质粒按2 ∶1的体积质量比转染进HepG2细胞,转染24 h后加含800 mg/L G418的DMEM完全培养基进行筛选培养,以重组PXR(n=1)为对照,利用Western blot检测PXR-HepG2和HepG2细胞(n=2)中 PXR的表达水平,筛选得到高表达PXR-HepG2细胞。
1.2.4IC-MAC处理PXR-HepG2和HepG2细胞 IC-MAC用二甲基亚砜(dimethyl sulfoxide,DMSO)配成母液1 000×(25、50及100 g/L),-20 ℃避光保存,加药时用预热至37℃的完全DMEM培养基稀释至1×(25、50及100 mg/L)。将细胞分为阴性对照组(DMSO组,0.1%)、药物处理组(25、50及100 mg/L IC-MAC)和阳性药物组[PXR激动剂利福平(rifampin,RIF;10 μmol/L)],分别处理24、48及72 h后,提取各组细胞总蛋白备用。
1.2.5PXR和CYP3A4蛋白表达 采用Western blot检测,制备10%的SDS-PAGE凝胶电泳的分离胶,在样品孔中依次加入1.2.3项下或1.2.4项下各组蛋白样品和蛋白Marker,80 V电泳20 min后再继续以120 V电泳60 min。电泳结束后利用半干转仪器将蛋白转移至已活化的PVDF膜上,并用SNAP快速转膜仪进行封闭和抗体孵育。BSA封闭30 min,一抗孵育(CYP3A4一抗孵育20 min,PXR一抗孵育30 min),TBST洗涤3 min,洗涤完毕后加二抗孵育10 min;TBST洗涤3 min,于PVDF膜上加发光液1 mL,室温反应1 min后用凝胶成像仪器拍照。
1.3 统计学分析
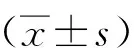
2 结果
2.1 高表达PXR的PXR-HepG2细胞模型构建
作为阳性对照,重组PXR蛋白出现单一条带,说明所用抗体能正确识别PXR蛋白;PXR-HepG2细胞中的PXR蛋白表达明显高于HepG2细胞,且差异有统计学意义(P<0.01),提示高表达PXR的PXR-HepG2细胞模型构建成功。见图2。

注:A为PXR蛋白电泳条带,B为PXR蛋白相对表达量;(1)与HepG2组比较,P<0.01。图2 PXR-HepG2和HepG2细胞中PXR的蛋白表达Fig.2 PXR protein expression in PXR-HepG2 and HepG2 cells
2.2 IC-MAC对PXR-HepG2细胞中CYP3A4蛋白表达的影响
作为阳性药物,RIF在PXR-HepG2细胞和HepG2细胞中均能上调CYP3A4的表达,提示PXR-HepG2和HepG2细胞对PXR激动剂敏感。25、50及100 mg/L IC-MAC分别干预PXR-HepG2和HepG2细胞24、48及72 h后,2种细胞中CYP3A4蛋白表达水平均较DMSO组上调,且24 h时PXR-HepG2组的上调倍数高于HepG2细胞,差异均有统计学意义(P<0.01)。见图3。

注:A、B分别为IC-MAC处理后PXR-HepG2和HepG2细胞中CYP3A4蛋白电泳条带,C、D分别为IC-MAC处理后PXR-HepG2和HepG2细胞中CYP3A4蛋白相对表达量;与DMSO组比较,(1)P<0.05,(2)P<0.01。图3 IC-MAC处理后PXR-HepG2和HepG2细胞中CYP3A4的蛋白表达Fig.3 CYP3A4 protein expression in PXR-HepG2 and HepG2 cells after treatment with IC-MAC
3 讨论
PXR是核受体超家族的成员,具有调节基因表达的功能[17-18]。当PXR与配体结合后,构象发生改变,与视黄醛X受体形成异二聚体,同时招募辅活化因子或辅阻遏物形成复合物,然后结合到靶基因启动子上的顺式作用元件,调节靶基因的转录[19]。自2000年Moore等[20]发现圣约翰草能激活PXR、诱导CYP3A4的表达后,PXR逐渐成为研究P450酶表达调控的热点靶蛋白。PXR调控的基因非常广泛,包括Ⅰ相代谢酶、Ⅱ相代谢酶和转运蛋白等[17-18]。研究表明,PXR是CYP3A4的主要转录调控因子[21-22]。
HepG2是一种永生化人肝癌细胞系,目前作为常用的体外细胞模型被广泛用于CYP3A4的相关研究实验中[23-24]。HepG2细胞虽具有较完整的Ⅰ相代谢酶表达,但PXR和CYP3A4的表达水平不高,通过瞬时转染实现外源性PXR的高表达可以达到考察PXR功能的目的[25]。因此,本文选用HepG2细胞作为PXR高表达的体外转基因模型的载体,考察IC-MAC是否通过PXR途径来调节CYP3A4的表达。实验结果显示,构建的PXR-HepG2细胞模型能高表达PXR,且PXR激动剂RIF能诱导CYP3A4的表达,表明体外细胞模型构建成功。
在此基础上,本研究分别用不同浓度的IC-MAC处理PXR-HepG2细胞和HepG2细胞,与阴性对照组相比,IC-MAC均能诱导2种细胞中CYP3A4的表达;经IC-MAC处理24 h后,相较于48和72 h,PXR-HepG2细胞中CYP3A4表达上调幅度最高;在瞬时转染的条件下,外源基因的表达会随着时间的推移逐渐降低;在24 h时,PXR-HepG2细胞中PXR的表达量可能达到最高值,然后逐渐下降,其对CYP3A4的转录调节能力也随之变化。所以IC-MAC处理后,PXR-HepG2细胞中CYP3A4表达水平在24 h的上调幅度最大。在24 h时,与HepG2细胞相比,PXR-HepG2细胞中CYP3A4表达水平上调幅度明显更大,说明IC-MAC可能通过PXR来上调CYP3A4的表达。
课题组前期曾利用Cocktail 探针药物法考察IC-MAC对大鼠细胞色素P450 主要亚型酶(CYP3A4、CYP1A2、CYP2C9、CYP2C19、CYP2D6和CYP2E1)比活性的影响,结果发现IC-MAC可抑制CYP3A4的比活性[16],但是本研究却发现IC-MAC可通激活PXR促进CYP3A4的表达。影响酶比活性的途径通常有2种,一是改变酶活性,二是通过改变酶的表达来增加或减少单位蛋白内的酶含量[26]。本文研究IC-MAC对PXR的基因转录调节活性的影响,主要是从酶表达调控的角度来研究IC-MAC下调CYP3A4活性的机制。因此,PXR被激活后,首先是上调CYP3A4 mRNA表达,进而增加其蛋白表达。有研究表明,蛋白翻译后修饰可能会导致P450酶mRNA,蛋白表达和活性不一致的现象[27]。因此,可推测IC-MAC 对CYP3A4表达及活性的影响是一个复杂的过程,可能涉及到蛋白翻译后修饰。在下一步的研究中,将从导致蛋白降解的泛素化修饰入手,以期解释IC-MAC处理后CYP3A4表达和活性不一致的问题。
猜你喜欢
江西农业学报(2021年4期)2021-04-20
江西医药(2020年4期)2020-04-28
食品安全导刊(2019年30期)2019-01-05
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
中国医药生物技术(2015年4期)2015-12-26
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
