
斑马鱼mcm3合子基因敲除实验模型的构建与鉴定*
2021-08-05 01:49夏佳敏陈炳宇黄四洲
成都医学院学报 2021年4期
夏佳敏,张 可,陈炳宇,黄四洲,刘 敏
1.成都医学院 基础医学院(成都 610500);2.成都医学院 人体解剖与组织胚胎学教研室(成都 610500); 3.成都医学院 发育与再生四川省重点实验室(成都 610500);4.成都医学院第一附属医院(成都 610500)
微小染色质复合物(minichromosome maintenance,MCM)蛋白因参与DNA复制起始而被人们熟知。MCM蛋白具有DNA解旋酶功能,是DNA复制的核心,并维持基因组的稳定性,参与DNA复制及细胞增殖。MCM2-7复合体是在细胞周期进展中存在的一种蛋白质[1],而mcm10则在DNA复制过程中介导RECQL4和MCM2-7复合物之间的相互作用[2]。MCM蛋白不仅与细胞周期有重要联系,还与癌症的发展密切相关。在多种癌组织和癌细胞系中检测到多种MCM蛋白过表达:mcm2与肺鳞状细胞癌恶性程度呈正相关[3];mcm2、mcm5、mcm6和mcm7在肾细胞癌中表达水平增高[4];mcm2和mcm5在胃肠道肿瘤的表达中具有临床意义[5];mcm2、mcm3和mcm7 mRNA的高表达可能是神经胶质瘤的分子预后标志物[6];mcm6被认为是S/G2细胞周期进程的驱动力,是肝癌潜在的诊断和预后标志物[7]。MCM家族在DNA复制及体外细胞水平上的研究颇多,但其在发育中的研究鲜有报道。关于秀丽隐杆线虫研究[8]发现,mcm4在皮下组织中的表达可以回救mcm4突变导致的生长迟缓,并且减少致死率。也有研究[9]表明,MCM2-7亚基复合体在拟南芥种子发育过程中呈现出非常重要的作用。由于细胞DNA在胚胎早期发育阶段和肿瘤细胞中均以快速复制的形式保证细胞增殖,故mcm3家族成员在肿瘤细胞中的作用,暗示其可能在胚胎发育中发挥重要作用。
MCM3是MCM复合体蛋白成员之一,其同样作为DNA复制中一种很重要的蛋白被广泛研究。Gao等[10]研究发现,PLK1介导的mcm3磷酸化是调节肾细胞癌增殖和凋亡的新机制。研究[11]表明,mcm3在大鼠中还参与了神经元的凋亡。临床研究[12]发现,肝癌组织中的mcm3表达明显高于正常组织。还有研究[13]发现,mcm3通过激活NF-κB通路促进肝癌放射治疗的耐受性。本课题组前期研究[14]发现,斑马鱼合子基因mcm3的敲降会导致肝脏变小。因此,mcm3除在细胞周期及疾病中发挥作用,在胚胎发育中也起到重要作用,但其在胚胎发育中的具体作用和调控机制还尚未阐明。由于斑马鱼的mcm3基因与人类mcm3基因具有很高的同源性,且斑马鱼产卵量大、培养成本低、周期短、胚胎早期透明等,故本研究以斑马鱼为动物模型,利用CRISPR/Cas9技术构建mcm3基因敲除的斑马鱼突变体,为进一步研究mcm3在胚胎发育中的作用及调控机制奠定了基础。
1 材料与方法
1.1 实验动物
本研究所用斑马鱼品系为野生型(AB品系)。斑马鱼严格按照标准饲养条件喂养,保持规律的昼夜交替。成年斑马鱼在28.5 ℃的系统水中养殖,系统水保持每日紫外线照射杀菌,每日监测pH值使其维持在7.6左右。在成年斑马鱼交配前1 d,将其放置在交配缸中,缸中放适量养殖系统水,并用塑料透明挡板分开雌、雄斑马鱼,于次日8∶00前将挡板拔除。在光照刺激下,斑马鱼开始产卵,30 min后即可用无菌的养殖系统水进行收集,收集后的胚胎放置于28 ℃恒温箱中培养,在胚胎受精12 h后,加入0.03%苯硫脲抑制黑色素形成。
1.2 序列比对分析
从国家生物技术信息中心(NCBI)数据库(https://www.ncbi.nlm.nih.gov/)中收集人类、小鼠和斑马鱼的MCM3氨基酸和核苷酸序列。氨基酸序列号如下:斑马鱼(AF506208)、人类 (NM_002388.6)、小鼠(NM_008563XM_001005862)、鸡(NM_001006421.1)、爪蟾(NM_001045766.1)等,采用Mega7.0、Jalview等软件对序列进行比对分析。
1.3 靶位点选择和sgRNA设计及合成
在NCBI网站上下载斑马鱼mcm3的全长序列,利用SnapGene软件对mcm3设计导向 RNA (guide RNA, gRNA) 靶位点,设置Cas9靶位点的前提:Cas的靶点应该包括20个碱基, 5′端为GG;靠近靶点3′端的3个碱基构成PAM区,序列为NGG(N为随意碱基)。以sgRNA1-F/R或sgRNA2-F/R为引物进行PCR,在一定的条件下(退火温度60 ℃,延伸时间30 s),用Primer-Star高保真酶(Takara,美国)进行扩增。随后以该PCR扩增的产物为模板,用T7转录试剂盒(Ambion,美国)进行转录,获得mcm3基因靶位点和sgRNA模板序列的sgRNA。转录产物用RNA纯化试剂盒进行纯化回收,测定浓度及电泳检测后-20 ℃保存备用。
1.4 探针制备及原位杂交
反义探针以cDNA为模板,由MCM3-F和MCM3-R(表1)通过PCR得到DNA产物,用PCR纯化试剂盒(Omega,美国)进行回收。用DIG RNA Labeling Kit(SP6/T7)试剂盒(Roche,美国)按如下反应体系制备探针:DNA模板3 μL (0.8 μg),Transcription Buffer(10×) 1 μL,Dig-labeled NTP mix(10×) 1 μL,T7 RNA聚合酶 0.5 μL,RNase inhibitor 0.5 μL,加RNase-free water至10 μL,在37 ℃反应2 h后加入0.5 μL T7 RNA聚合酶,在37 ℃反应1 h,最后加入1 μL不含RNA酶的DNase Ⅰ在37 ℃反应30 min,按80 μL异丙醇、10 μL乙酸钠、100 μL RNA free H2O的体系进行纯化,测浓度及电泳后,于 -20 ℃保存备用。斑马鱼胚胎收集后,用4%多聚甲醛固定过夜,第2天用磷酸盐吐温缓冲液漂洗(5 min/次,洗3次),用无水甲醇脱水漂洗(15 min/次,洗3次),最后放入-20 ℃冰箱备用。原位杂交按参考文献[15]方法进行。

表1 Real time RT-PCR引物
1.5 显微注射
使用显微注射仪Pico Pump及Olympus SZX2-ILLT荧光显微镜,在1细胞期向胚胎注射Cas9核酸酶(NEB,美国)和sgRNA混合液,将注射后的胚胎在28.5 ℃恒温培养箱中孵育并定期换水。
1.6 突变体的筛选
注射后第2天收集部分胚胎,提取基因组进行PCR扩增,用T7E1酶进行酶切检测突变情况,T7E1酶主要用于识别DNA上错配的碱基,并对识别出来的位点进行切割。T7E1酶检测到突变后,将剩余F0代胚胎饲养至性成熟后与野生型(AB品系)交配即得F1代。成年后对F1代的斑马鱼拔取鳞片,提取基因组目的DNA以进行PCR扩增,PCR扩增后的产物进行PCR纯化后测序,从而确定突变体类型。将获得突变体类型的F1代杂合体与野生型(AB品系)交配,得到F2代。待F2代斑马鱼培养至性成熟后,同样对其拔取鳞片以提取基因组目的DNA进行PCR扩增;随后将PCR产物通过基因测序检测目的片段序列,确定mcm3杂合体;最后mcm3杂合体自交可得mcm3突变体。
1.7 荧光显微镜拍照
当胚胎发育到一定时期,选取胚胎将其麻醉,使用低熔点琼脂凝胶固定合适体位后于共聚焦下拍照。原位杂交实验后的胚胎则使用甘油或低熔点琼脂凝胶固定后拍照,使用的显微镜为荧光显微镜或激光共聚焦显微镜(Olympus,日本)。
2 结果
2.1 MCM3生物信息学及基因结构分析
本研究对NCBI网站报道的9种MCM3蛋白进行了系统发育树分析,以了解斑马鱼MCM3与其他物种间的关系。系统发育树分析表明,这9种MCM3蛋白起源于同一祖先,其中人类mcm3基因与黑猩猩mcm3基因最接近,小鼠mcm3基因与牛mcm3基因最接近,而斑马鱼mcm3基因与非洲爪蟾基因最接近(图1A)。
氨基酸同源性分析表明,MCM3蛋白序列在人、小鼠、斑马鱼中高度保守,人与小鼠的氨基酸同源性为87.73%,人与斑马鱼的氨基酸同源性为85.57%,小鼠与斑马鱼的氨基酸同源性为85.57%(图1B)。这一结果提示斑马鱼MCM3与人、鼠MCM3相似,在维持生理功能和疾病发生中发挥重要作用。mcm3全长13 464 bp,共有17个外显子(图1C)。

图1 mcm3系统进化树分析及sgRNA靶位点设计
2.2 CRISPR/Cas9技术构建mcm3突变体及F0代胚胎的检测筛选
酶切检测结果显示,野生型的PCR产物不能被T7E1酶切开,而注射成功的胚胎能够被部分切开(图2)。该结果证明,F0代部分胚胎含有靶位点突变,因此将T7E1酶切成功的F0代胚胎培养为成年斑马鱼。

图2 斑马鱼F0代T7E1酶切检测
2.3 mcm3突变体F1和F2代稳定品系的筛选
基因测序检测结果发现,有1条斑马鱼在外显子3上插入1个碱基A,编码178号缬氨酸的密码子 (GTT)变为终止密码子 (TGA),导致翻译提前终止。因此将该突变的mcm3杂合体与野生型(AB品系)继续交配,得到F2代mcm3杂合体,随后mcm3杂合体自交即得mcm3纯合突变体(图3)。
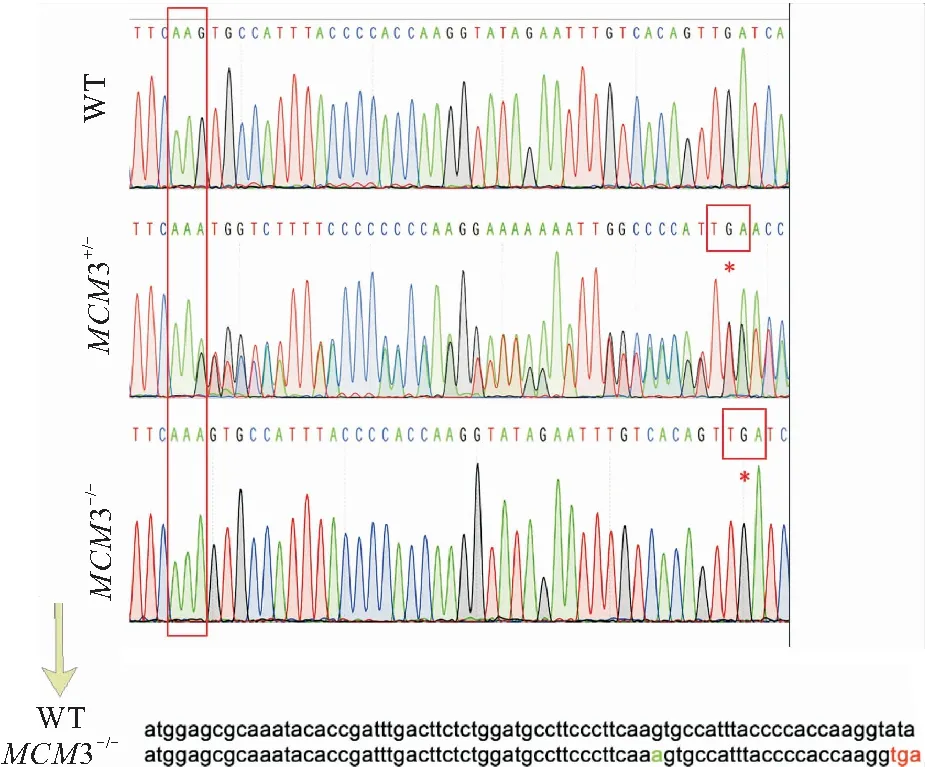
图3 获得可稳定遗传mcm3突变体斑马鱼
2.4 突变体中mcm3 mRNA的表达情况
整胚原位杂交实验结果显示,mcm3为合子基因,无论是野生型还是突变体,在16 cell时期没有表达;在野生型胚胎中的shield时期及bud时期则广泛表达;而在咽囊期24 h,mcm3主要在头部、眼睛、体节以及内胚层区域表达;孵化期48~72 h在头部、鱼鳃及胸腺等区域高表达,且在肝等内胚层组织器官也有较强表达。与野生型的mcm3 mRNA表达情况相比,mcm3突变体在以上描述时期及部位中的表达均明显减少,表明mcm3合子基因在mcm3突变体中被成功敲除(图4)。

图4 mcm3在野生型及突变体斑马鱼胚胎各时期的表达模式(3.2×)
2.5 mcm3突变体斑马鱼与野生型斑马鱼表型比较
受精早期,mcm3突变体与野生型斑马鱼相比外观上无明显变化。受精3 d后,与野生型斑马鱼相比,突变体眼睛和头变小,且突变体胚胎不长鱼鳔(图5A~B),但并未发现其他明显的发育缺陷(图5A)。受精4 d后,突变体的眼睛明显较野生型小(图5B~C),mcm3突变体在受精8 d后死亡(图5D)。实验结果表明,合子基因mcm3在斑马鱼早期胚胎发育过程中起到重要作用。

图5 mcm3突变体斑马鱼与野生型斑马鱼胚胎时期表型比较
3 讨论
突变体构建的方法有很多种,比如传统的转录激活因子样效应子(TALEN)[16]、锌指核酸酶(ZFN)[17]等。本研究选择CRISPR/Cas9技术,操作简便,且更能高效敲除目的基因。
mcm3是参与复制前复合物形成的MCM家族的成员之一,进化上具有高度保守性,主要参与启动DNA复制。mcm3在细胞周期中的作用已被证实,但mcm3在胚胎发育中的作用研究还很少。本课题组前期应用斑马鱼验证了mcm5在体节发生时期以非细胞周期依赖性的方式进行了调控[18],并且斑马鱼合子基因mcm3的敲降会导致肝脏变小[14],那么mcm3是否也以非细胞周期依赖性调控斑马鱼胚胎的发育尚不得而知。目前对于mcm3基因突变体的研究并不多。本研究通过CRISPR/Cas9技术,在mcm3基因的第3、4个外显子处设计sgRNA靶位点,成功构建了在3号外显子上插入突变可稳定遗传mcm3突变体斑马鱼。原位杂交结果表明,mcm3为合子基因,在胚胎卵裂期不表达,bud及shield时期广泛表达,咽囊期后24~72 h主要表达于头部、胸腺、内胚层等区域,而在突变体中这些部位的表达均明显减少;同时突变体斑马鱼胚胎早期呈现出小头、小眼睛的表型。这些结果表明,mcm3可能参与了斑马鱼眼睛、内胚层及神经的发育。
mcm3在肾癌、肝癌等组织中呈现高表达。研究[13]表明,mcm3可以通过激活NF-κB途径以促进肝细胞癌的放射耐药性, MCM3蛋白的表达与肝细胞癌组织中肿瘤侵袭增加相关[12],但mcm3是否可以直接或间接调节肝癌的生长尚未见报道。研究[19]发现,mcm3和Ki67在最常见的牙源性囊肿和肿瘤中表达,尤其是在侵袭性较强的病变中。临床研究[20]发现,在浸润性乳腺导管癌中,mcm3与肿瘤大小相关,mcm3高表达与不良预后相关。目前,关于mcm3在胚胎发育及神经发育中的研究鲜见相关报道,后续不仅可通过该突变体对mcm3基因在肿瘤方面的具体功能进行研究,还可对mcm3影响胚胎发育的具体调节机制进行研究;同时观察mcm3是否对内胚层器官或其他胚层及神经的发育有影响。
综上所述,选用CRISPR/Cas9成功构建mcm3突变体斑马鱼,该突变体呈现小头、小眼睛的表型,并且在受精8 d后突变体死亡,说明mcm3在胚胎发育中起到重要作用,所以构建该突变体为后续研究mcm3基因在胚胎发育中的功能提供了基础。
猜你喜欢
作物学报(2022年2期)2022-11-06
安徽医科大学学报(2022年6期)2022-07-13
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
皮肤性病诊疗学杂志(2021年5期)2021-11-27
小天使·二年级语数英综合(2021年8期)2021-08-16
三农资讯半月报(2021年1期)2021-01-27
中国医药导报(2016年33期)2017-03-06
作文评点报·作文素材小学版(2016年8期)2016-03-16
现代养生·下半月(2015年8期)2015-11-16
